

Abstract
The aim of the present study was to investigate effect of exercise on learning and memory, long-term potentiation and levels of epinephrine in the rat hippocampus. Treadmill trained (one hour at 17 m·min-1 for 10 days) and corresponding control rats went through spatial learning process on a Morris water maze for 8 days. The time to reach the platform (latency), the length of swim path, and the swim speed were used for the evaluation of spatial learning. Our results showed that physical activity produced a significant enhancement in spatial learning, with a decreased path length (p<0.05) and latency (p<0.05) to the platform in Morris water maze, without affecting the swim speed. Furthermore, the levels of the epinephrine were significantly increased (p<0.05) in hippocampus of the exercised rats. In conclusion our findings suggest that the enhanced learning by exercise may be mediated through the activation of adrenoceptors in the hippocampus and epinephrine may play an important role in potentiation of learning.
Key Words: Exercise, memory, hippocampus, epinephrine
Introduction
Exercise increases neurogenesis in the dentate gyrus of the hippocampus, which is important for memory function. New neurons are added continuously to certain areas of the adult brain, such as the hippocampus and olfactory bulb (Gage et al., 1998). The functional significance of new cells in hippocampus is not clear. In birds, food storage and retrieval experience correlate with changes in hippocampal size and neurogenesis (Clayton and Krebs, 1994). In mice, neurogenesis in the dentate gyrus increases with exposure to an enriched environment, and it is associated with improved learning (Kempermann et al., 1997). Similarly, voluntary physical activity in a running wheel enhances the number of new hippocampal cells (Van Praag et al., 1999). Although it is not known whether running also affects learning, it has been shown that physical activity facilitates recovery from injury (Johansson and Ohlsson, 1997) and improves cognitive function (Fordyce and Wehner, 1993). Furthermore, trophic factors, associated with progenitor cell survival and differentiation (Ray et al., 1997), alterations in synaptic strength (Schuman, 1999), long-term potentiation (LTP) (Patterson et al., 1992), and memory function (Fischer et al., 1987), are elevated after exercise (Neeper et al., 1995). At the cellular level, wheel running enhances the firing rate of hippocampal cells in a manner that correlates with the running velocity (Czurko et al., 1999). Thus, exercise may increase synaptic plasticity and learning, as well as neurogenesis, but we have found no earlier reports in literature.
The aim of the present experiments was to test the hypothesis that exercise facilitates memory establishment and promotes LTP in rats. Furthermore, we aimed to measure the concentration of biogenic monoamines in the hippocampus.
Methods
Animals
Forty male outbreed, 9 weeks old Wistar rats were divided into two groups of 20, as control and exercised trained test group. Rats were fed ad libitum and housed as four animals to a cage at 22±2 °C room temperature, with 12:12 hrs light; dark cycles in the Finnish National Laboratory Center, University of Kuopio, Finland. The study was accepted by the Ethics Committee on Animal Experiments, University of Kuopio.
Exercise
All groups of animals were familiarized to treadmill apparatus in order to eliminate stress of exercise. Prior to spatial learning, test group was trained on a running treadmill for one hour at 17 m·min-1 for 10 days. All animals tolerated the speed and duration of the exercise and completed successfully the training session.
Spatial Learning
Control and test groups of rats were trained on a Morris water maze (Honma, 1986) with either two or four trials per day for eight days. The platform was hidden 1 cm below the surface of water. It was made opaque with white nontoxic paint. The starting points were changed every day. Each trial lasted either until the rat found the platform or for a maximum of 40 s. At the end of each trial, the rats were allowed to rest on the platform for 10 s. The time to reach the platform (latency), the length of swim path, and the swim speed were recorded semi-automatically by a video tracking system.
Sample preparation and analysis
Animals were sacrificed by decapitation at the end of behavioral testing. The brain was removed immediately on dry ice. Dopamine, 5-HT and norepinephrine and epinephrine were separated from rat hippocampus by modifying the method of Honma (Honma, 1986). Alumina cartridges supplied by International Sorbent Technology LTD (Mid Glamotgan, UK) were used. Norepinephrine, dopamine and serotonin amounts were too low to be measured in the samples and only the level of epinephrine was determined.
Epinephrine in rat hippocampus samples was measured by using the post-column system for polyamine analysis (Walters, 1984; Tracy et al., 1995). Samples were analyzed isocratically using the Pickering cation-exchange column, K+ form (Pickering Laboratories Inc., California, USA) and Gilson-system: pump 321, auto injector 234, interface 506C (Gilson S.A., Villiers-le-Bel, France). The fluorometric detector Jasco Model FP-1520 (Jasco Corporation, Tokyo, Japan) was used. Eluent flow-rate was 0.7 ml·min-1; column temperature 37°C and sample loop volume 20 µl. Epinephrine was quantified by comparing the peak heights of the samples with those given by known concentrations of standards.
Statistical Analysis
Comparisons between two groups were performed using the unpaired Student's t-test. Comparisons among several groups were performed using one-way analysis of variance. A value of p<0.05 was taken statistically significant.
Results
Physical exercise produced a significant enhancement in spatial learning as decreased path length (Figure 1a) and latency (Figure 1b) to the platform in the Morris water maze. These results were not confounded by swim speed, because there was no significant difference between the groups (Figure 1c). Mean epinephrine levels in eight days in hippocampus were significantly higher in the trained rats compared with their controls (Figure 2).
Figure 1.
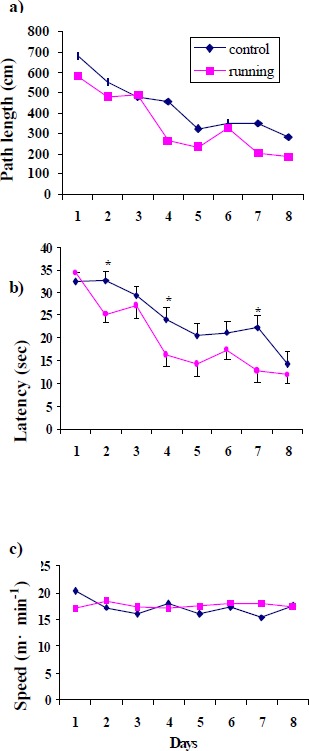
Water maze learning in control and runners trained with three trials per day. Rats were trained over eight days to find the hidden platform in the Morris water maze. Comparisons between two groups showed significant difference in runner compared with control rats in path length and latency, * p < 0. 05.
Figure 2.
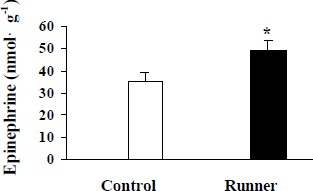
Comparison of mean level of epinephrine (nmol·g-1) in control and runners trained with three trials per day. Rats were trained over eight days to find the hidden platform in the Morris water maze. Comparisons between two groups showed significant difference in runner compared with control rats in mean level of epinephrine. * p< 0. 05.
Discussion
Our results showed that in adult rats increased physical activity significantly enhanced spatial learning performance, apparent with both decreased path length and latency finding the platform in Morris water maze. This is a novel finding.
Several factors that elevate production of new neurons are also associated with enhanced learning (Tracy et al., 1995). Both running and living in an enriched environment double the number of surviving newborn cells and improve water maze performance (Russo-Neustadt et al., 1999). Maintaining brain health and plasticity throughout life is an important public health goal. It is increasingly clear that behavioral stimulation and exercise can help us to achieve it. Over the past decade, a number of studies on humans have shown the benefits of exercise on brain health and function, particularly in aging populations (Ivy et al., 2001). It has been previously reported that running enhances neurogenesis, levels of brain-derived neurotrophic factor (BDNF) and other growth factors and neurotransmitters especially in the hippocampus (Honma, 1986). In addition, hippocampal-dependent learning may enhance survival of cells prior to spatial training (Nibuya et al., 1995). Moreover, treatment with hormones, such as estrogens, increases cell proliferation and improves memory function (Fujimaki et al., 2000).
Since mean levels of the hippocampal epinephrine in eight days training were increased significantly, our finding may suggests that the learning process may be mediated through activation of adrenoceptors in the dentate gyrus of the rat hippocampus. This enhancement of hippocampal epinephrine levels may play an important role in potentiation of learning in rat. In contrast, factors that reduce neurogenesis, such as corticosterone treatment, stress and aging are associated with diminished performance on spatial learning tasks (Liu and Mori, 1999).
Monoamine-mediated signaling also contributes to BDNF gene regulation. Several antidepressants that increase transmission at monoaminergic synapses also increase BDNF gene expression in the hippocampus (Russo-Neustadt et al., 1999; Nibuya et al., 1995). Interestingly, antidepressant treatment in combination with exercise enhances exercise-dependent BDNF upregulation in the hippocampus (Fujimaki et al., 2000). Therefore, noradrenaline-mediated signaling might be particularly important in the modulation of BDNF gene expression by exercise (Ivy et al., 2001).
Conclusion
In conclusion, our results emphasize the role of physical activity in the promotion of learning and memory functions. We suggest physical activity enhances leaning and memory functions probably by enhancing of epinephrine levels in rat hippocampus.
Biographies

Nasser AHMADIASL
Employment
Ass.Prof., Tabriz Univ., Iran
Degrees
Doctor of Veterinary Medicine PhD in Physiology
Research interests
Exercise physiology
E-mail: ahmadyn@tbzmed.ac.ir
Hojjatallah ALAEI
Employment
Department of Physiology, Faculty of Medicine, Esfahan University of Medical Sciences, Esfahan, Iran
Degrees
Doctor of Pharmacy, University of Esfahan, Iran,1982. PhD; Neurophysiology, Univ. of Sheffield, UK, 1990.
Research interest
Neurotransmitter function in the brain
E mail: alaei@med.mui.ac.ir

Osmo HÄNNINEN
Employment
Prof.of Physiology and Chairman of the Depart. of Physiology, Univ. of Kuopio, FIN
Degrees
MSc-Biochemistry, Univ. of Turku, FIN, 1962.
MD, Univ. of Turku, FIN, 1964.
DMS, in 1966.
PhD-Biochemistry, Univ. of Turku, FIN, 1968.
Assoc.Prof. of Physiology, Univ. of Turku, FIN, 1967.
Assoc. Prof. of Biochemistry, Univ. of Turku, FIN, 1969.
Prof. of Physiology, Univ. of Kuopio, FIN, 1972-.
Research interest
Muscle metabolism and function. Ergonomics. Biotransformation, biomonitoring and comparative biochemical toxicology.
E-mail: osmo.hanninen@uku.fi
References
- Clayton N.S., Krebs J.R. (1994) Hippocampal Growth and Attrition in Birds Affected by Experience. Proceedings of the National Academy of Sciences of the United States of America, 91, 7410-7414 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Czurko A., Hirase H., Csicsvari J., Buzsaki G. (1999) Sustained activation of hippocampal pyramidal cells by 'space clamping' in a running wheel. The European Journal of Neuroscience 11, 344-352 [DOI] [PubMed] [Google Scholar]
- Fischer W., Wictorin K., Bjorklund A., Williams L.R., Varon S., Gage F.H. (1987) Amelioration of cholinergic neuron atrophy and spatial memory impairment in aged rats by nerve growth factor. Nature (London) 329, 65-68 [DOI] [PubMed] [Google Scholar]
- Fordyce D.E., Wehner J.M. (1993) Physical activity enhances spatial learning performance with anassociated alteration in hippocampal protein kinase C activity in C57BL/6 and DBA/2 mice. Brain Research 619, 111-119 [DOI] [PubMed] [Google Scholar]
- Fujimaki K., Morinobu S., Duman R.S. (2000) Administration of a cAMP phosphodiesterase 4 inhibitor enhances antidepressant-induction of BDNF mRNA in rat hippocampus. Neuropsychopharmacology 22, 42-51 [DOI] [PubMed] [Google Scholar]
- Gage F.H., Kempermann G., Palmer T.D., Peterson D.A., Ray J. (1998) Multipotent progenitor cells in the adult dentate gyrus. Journal of Neurobiology 36, 249-266 [DOI] [PubMed] [Google Scholar]
- Honma T. (1986) Comparative study of fluorometric and electrochemical detection of catecholamine, serotonin, and metabolites in rat brain by high-performance liquid chromatography. Analytical Letters 19, 417-431 [Google Scholar]
- Ivy A.S., Rodriguez F.G., Russo-Neustadt A. (2001) The effects of NE and 5-HT receptor antagonists on the regulation of BDNF expression during physical activity. Society for Neuroscience Abstracts 253, 213 [Google Scholar]
- Johansson B.B., Ohlsson A. (1997) Environment, Social Interaction, and Physical Activity as Determinants of Functional Outcome after Cerebral Infarction in the Rat. Experimental Neurology 139, 322-327 [DOI] [PubMed] [Google Scholar]
- Kempermann G., Kuhn H.G., Gage F.H. (1997) More hippocampal neurons in adult mice living in an enriched environment. Nature (London) 386, 493-495 [DOI] [PubMed] [Google Scholar]
- Liu J., Mori A. (1999) Stress, aging, and brain oxidative damage. Neurochemical Research 11, 1479-1497 [DOI] [PubMed] [Google Scholar]
- Neeper S.A., Gomez-Pinilla F., Choi J., Cotman C. (1995) Exercise and brain neurotrophins. Nature (London) 373, 109. [DOI] [PubMed] [Google Scholar]
- Nibuya M., Morinobu S., Duman R.S. (1995) Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. Journal of Neuroscience 15, 7539-7547 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patterson S.L., Grover L.M., Schwartzkroin P.A., Bothwell M. (1992) Neurotrophin expression in rat hippocampal slices: a stimulus paradigm inducing LTP in CA1 evokes increases in BDNF and NT-3 mRNAs. Neuron 9, 1081-1088 [DOI] [PubMed] [Google Scholar]
- Ray J., Palmer T.D., Suhonen J., Takahashi J., Gage F.H. (1997) Isolation, Characterization, and Utilization of CNS Stem Cells. Christen Y., Gage F. H.Springer, Berlin: 129-149 [Google Scholar]
- Russo-Neustadt A., Beard R.C., Cotman C.W. (1999) Exercise, antidepressant medications, and enhanced brain derived neurotrophic factor expression. Neuropsychopharmacology 21, 679-682 [DOI] [PubMed] [Google Scholar]
- Schuman E.M. (1999) Neurotrophin regulation of synaptic transmission. Current Opinion in Neurobiology 9, 105-109 [DOI] [PubMed] [Google Scholar]
- Tracy M.L., Pickering M.V., VerHulst T. (1995) Food Testing and Analysis 1, 48-50 [Google Scholar]
- Van Praag H., Kempermann G., Gage F.H. (1999) Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nature Neuroscience 2, 266-270 [DOI] [PubMed] [Google Scholar]
- Walters M.J. (1984) Determination of histamine in fish by liquid chromatography with post-column reaction and fluorometric detection. Journal of the Association of Official Analytical Chemists 67, 1040-1043 [PubMed] [Google Scholar]