
Figure 10.
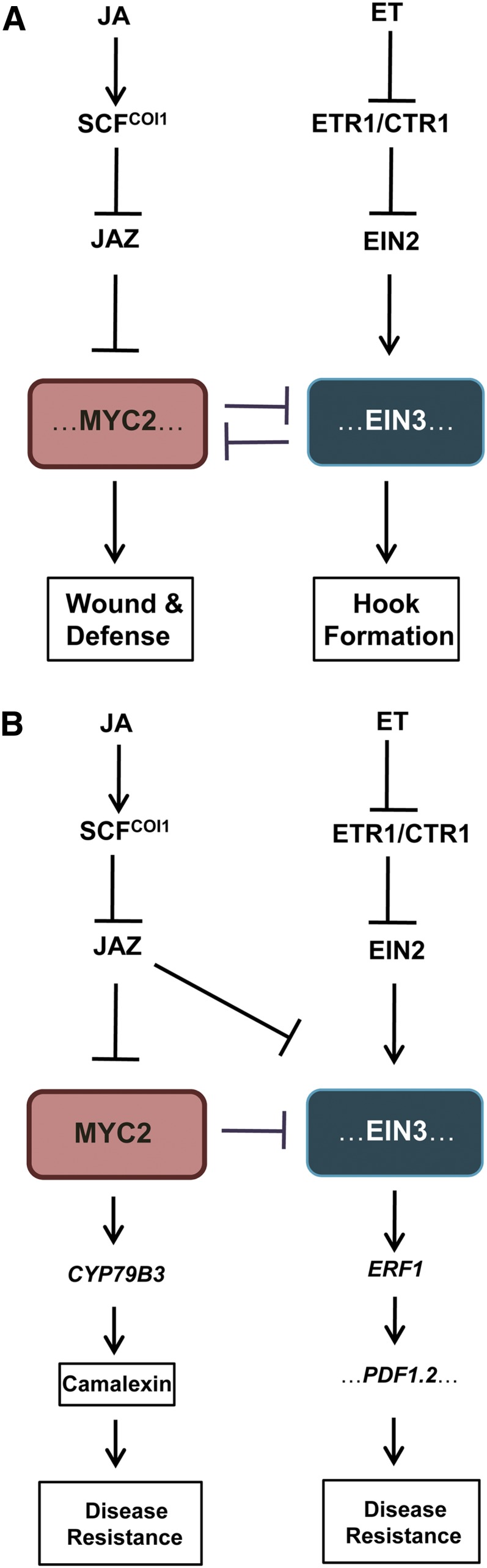
A Simplified Model for JA and ET Signaling Antagonism.
(A) Model for JA and ET antagonistic action in regulating hook curvature, wounding, and defense against insect attack. In response to JA signaling, SCFCOI1 recruits JAZs for ubiquitination and degradation. MYC2, MYC3, and MYC4 (indicated as MYC2) are then released to interact with and repress EIN3 and EIL1 (indicated as EIN3), which leads to attenuation of ET-enhanced hook curvature. ET signal inactivates the ET receptors (indicated as ETR1) and the negative regulator CTR1 to mediate EIN2 translocation into nucleus and to stabilize EIN3 and EIL1. EIN3 and EIL1 then interact with and repress MYC2, MYC3, and MYC4 to inhibit expression of wound responsive genes (e.g., VSP1, VSP2, and TAT3) and herbivory-inducible genes (e.g., CYP79B3, BCAT4, and BAT5) and suppress JA-regulated plant defense against generalist herbivores S. littoralis and S. exigua (indicated as wound and defense).
(B) Model for JA and ET crosstalk in regulating plant resistance against necrotrophic pathogen. JAZs and MYC2 interact with and repress ET-stabilized EIN3 and EIL1 (indicated as EIN3). In response to JA signaling, JAZ proteins are degraded to derepress EIN3/EIL1, leading to the increased disease resistance against necrotrophic pathogen B. cinerea (indicated as disease resistance) (Zhu et al., 2011). Meanwhile, JA-induced JAZ degradation releases MYC2, which counteracts EIN3 and EIL1 to prevent excessive disease resistance responses. In addition, other factors, including CYP79B3, which is required for biosynthesis of camalexin (Glawischnig et al., 2004; Kliebenstein et al., 2005), may be also regulated by MYC2 to modulate disease resistance. Regulation of plant resistance against B. cinerea might be complicated and modulated by the coordinated action of synergistic and antagonistic mechanisms.
[See online article for color version of this figure.]