
Figure 2.
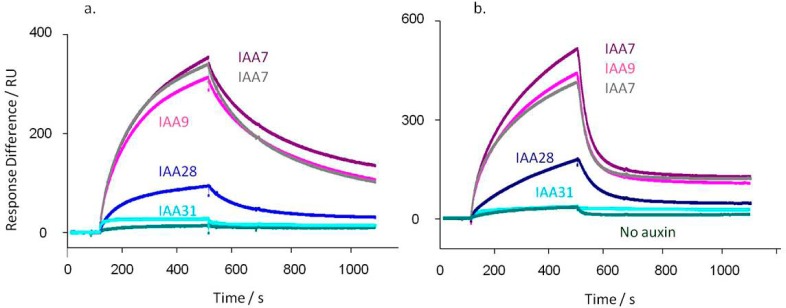
TIR1 (a) and AFB5 (b) co-receptor assembly is dependent on degron sequence. In each case TIR1 or AFB5 protein was mixed prior to injection with IAA at 50 μM, except for the control (no auxin). (a) Calculated kd’s for TIR1: IAA7 kd = 3.3 × 10–3 s–1; IAA7 repeat kd = 4.1 × 10–3 s–1; IAA9 kd = 3.5 × 10–3 s–1; IAA28 kd = 5.8 × 10–3 s–1; IAA31 kd = 3.6 × 10–3 s–1. X2 for the set = 1.24. (b) AFB5 dissociation rates are more rapid than for TIR1: IAA7 kd = 0.019 s–1; IAA7 repeat kd = 0.019 s–1; IAA9 kd = 0.020 s–1; IAA28 kd = 0.011 s–1; IAA31 kd = 0.002 s–1. X2 for the set = 0.70. Assays were set up with channel 1 blocked with biocytin, channel 2 coated with IAA7, and two other peptides on channels 3 and 4. Channel surfaces were saturated with biotinylated peptide in all cases. The IAA7 signal was available to normalize responses between chips, although within batches of protein this was not found necessary. The binding assays were done using a series of auxins. Only data collected at 50 μM IAA are shown, plus one of the series of control injections without auxin.