

Abstract
Homeotherms maintain an optimal body temperature that is most often above their environment or ambient temperature. As ambient temperature decreases, energy expenditure (and energy intake) must increase to maintain thermal homeostasis. With the wide spread adoption of climate control, humans in modern society are buffered from temperature extremes and spend an increasing amount of time in a thermally comfortable state where energetic demands are minimized. This is hypothesized to contribute to the contemporary increase in obesity rates. Studies reporting exposures of animals and humans to different ambient temperatures are discussed. Additional consideration is given to the potentially altered metabolic and physiologic responses in obese versus lean subjects at a given temperature. The data suggest that ambient temperature is a significant contributor to both energy intake and energy expenditure, and that this variable should be more thoroughly explored in future studies as a potential contributor to obesity susceptibility.
Keywords: Ambient Temperature, Obesity, Thermogenesis, Energy Expenditure, Brown Adipose Tissue, Diet, Thermoneutral
Introduction
The contemporary trend of increased prevalence of overweight and obesity in developed countries, including the United States, has brought with it an increasing concern of associated morbidity and mortality [1]. Whether the trend continues and the incidence increases or plateaus at the current level [2;3], an explanation for the underlying causes of this increase is somewhat disputed. Although it is well appreciated that a positive energy balance describes the situation where excess energy intake and/or decreased energy expenditure can clearly model weight gain, this descriptive statement does not address the underlying cause(s) and may not fully address the complexity of interactions that modulate both the food individuals eat nor the processes/activities that account for energy expenditure. In this regard, it has been suggested that multiple, less well appreciated factors may be additionally contributing to the increase in observed weight gain [4;5]. Humans, like other homeothermic mammals, have an optimal body temperature that is most frequently above the temperature of the environment in which they live. Thus, a cooler ambient temperature (Ta) results in a higher metabolic rate to meet the increased thermogenic demand. With the advent and wide spread adoption of climate control by heating and air conditioning in cars, homes and places of work, people enjoy a greater proportion of their daily lives in comfortable temperatures [4–6] that results in decreased energy expenditure for active thermal regulation – the thermoneutral zone (TNZ). It has been suggested that a greater amount of time spent within the TNZ and reduced variability (i.e. exposures to diurnal fluctuations to Ta) are associated with and contributes to increases in obesity rates, presumably through reductions of energy expenditure in the absence of cold or heat stress [4–7]. How this increase in the environmental comfort of life may be contributing to the rise in obesity and whether alterations in Ta exposure could offer a potential avenue to alter energy balance in a favorable way for weight loss or weight maintenance is discussed below.
Ambient Temperature: Energy Expenditure & Energy Intake
It is simple to recognize that if an individual eats more calories than they expend, the result should be weight gain. Additionally, a constant caloric intake concomitant with a significant decrease in energy expenditure – associated with reduced physical activity (or a sedentary lifestyle) – might also be expected to result in weight gain. Despite these obvious predictions and the contributions of each to energy balance, it has been difficult to ascribe either or both as the sole cause(s) of the increased overweight/obesity incidence in contemporary society. This has led some researchers to hypothesize that more subtle, but potentially significant factors, are individually or cooperatively contributing to a shift in the economy of energy balance and increase in obesity rates.
To be a significant contributor to the current obesity epidemic, any energy imbalance, no matter how big or small, must persist over an extended period of time. This has been clearly illustrated by modeling the impact of a fixed caloric excess (or reduced expenditure) on body weight gain [8;9]. The initial imbalance results in additional body mass accrual (both fat and lean mass), and a new energy balance is established within a relatively short period of time where the calories that were previously in excess are now necessary for body weight maintenance due to the increased body size [9]. Instead, if a relative caloric excess is maintained constantly over time a slow, but consistent, rise to a higher body weight will be attained.
One ubiquitous factor influencing both energy expenditure and energy intake is the ambient or environmental temperature in which we live. Homeothermic organisms, including humans, maintain an optimal body temperature within a fairly narrow range under normal conditions at ~37°C [10]. However, many homeotherms spend the majority of their time in an environment where the Ta is below their body temperature. This is acceptable as the basic processes of metabolism and physiology are subject to entropic loss resulting in heat production. Therefore, a thermal heat sink of lower Ta, where excess heat produced by metabolism can be dissipated, is beneficial for maintaining homeothermy. When the Ta drops below the lower limit of the TNZ, adaptive and thermogenic mechanisms are activated resulting in heat production and core body temperature preservation [11;12]. Conversely, when Ta rises above the upper TNZ limit, physiologic processes to amplify heat dissipation are activated resulting in a metabolic increase [13;14]. The temperature range where neither active heat generation nor dissipation is required for thermal homeostasis is defined as the thermoneutral zone (TNZ). Metabolism (energy expenditure) is increased either above or below the thermoneutral zone [13–15] (figure 1). Without adequate energy intake compensation (reduction) to offset the lowered metabolic demand at thermoneutrality, an energy imbalance should result contributing to weight gain. Similarly, under a fixed, basal energy intake condition, increases in energy expenditure associated with reductions in Ta could result in weight loss.
Figure 1.
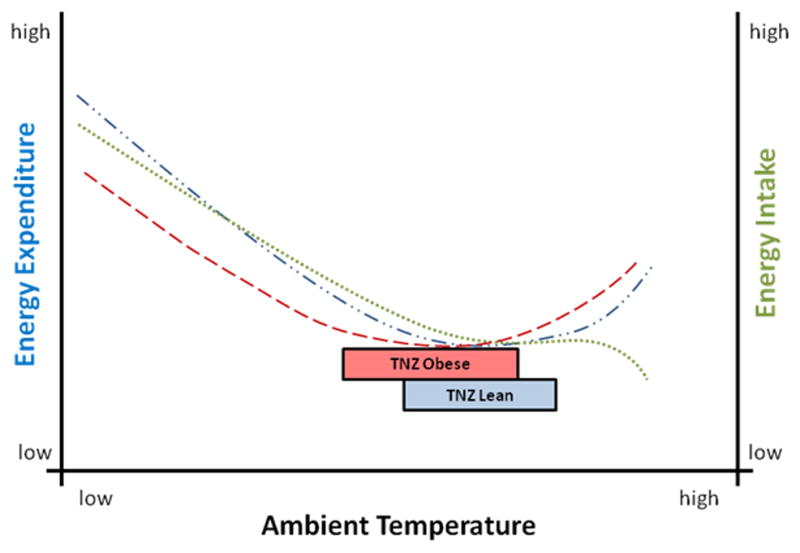
Energy Expenditure and Energy Intake at Different Ambient Temperatures. The thermoneutral zone (box area) is defined as the minimal basal energy expenditure when at rest in the absence of active thermogenesis or heat dissipation. Notice that as Ta decreases below the TNZ, energy expenditure (blue line
 ) increases. Conversely, as Ta increases above the TNZ, energy expenditure also increases. Food intake (green line
) increases. Conversely, as Ta increases above the TNZ, energy expenditure also increases. Food intake (green line
 ) is modeled across the Ta range based on small mammal and human data. Protection against Ta like the addition of clothing or changes in body composition (e.g. obesity/excess adiposity) shifts the energy expenditure curve to the left (red line
) is modeled across the Ta range based on small mammal and human data. Protection against Ta like the addition of clothing or changes in body composition (e.g. obesity/excess adiposity) shifts the energy expenditure curve to the left (red line
 ), representing an altered energetic response to the same Ta for individuals of different body weights and adiposities. Thus, the TNZ of obese individual may be expected to shift to a lower Ta range (box area).
), representing an altered energetic response to the same Ta for individuals of different body weights and adiposities. Thus, the TNZ of obese individual may be expected to shift to a lower Ta range (box area).
In homeotherms, Ta is not only associated with energy expenditure, but also energy intake [12;13;16–18]. Food intake functions both to meet physiological needs of macro and micronutrient requirements, but also serves as a significant source of energy for heat production and thermal homeostasis. When calories are insufficient, reductions in body temperature are possible [19]. With heat loss to the environment dependent on Ta (the lower the Ta below body temperature, the greater the heat loss), a significant, inverse relationship exists between Ta and food intake [16]. This increase in food intake with lowered Ta is observed across a wide range of temperatures in multiple species [12], including humans [13;20], although the amount of data collected on controlled human exposures to Ta, particularly above the TNZ, is limited compared with animal studies [16–18]. In general, exposure to Ta below the TNZ increases appetite and food intake while Ta above the TNZ results in a heat stress suppressing food intake, while inducing an elevated metabolic response (see references in [18] & [17]) (figure 1). This energetic divergence between intake and expenditure above the TNZ results in acute body weight and body fat loss in animals, with suggestive data available in humans [16–18;21–23]. Restaurants have capitalized on this environmental factor realizing that air conditioning in the summer helps increase sales, while more subtle alteration of ambience like room color can have an effect on temperature sensation with warm colors (reds, oranges and yellows) permitting a lower Ta before cold is perceived [17;24]. The variability of Ta and duration of exposure, whether chronic or acute, can have both individual and synergistic effects on energy intake patterns. Consistent cold exposure – housing and feeding – resulted in greater food intake than thermoneutral exposure in rats [25]. Similarly, rats that ate in the cold consumed more food than those fed at thermoneutrality, regardless of the general housing temperature, with greater weight gain in animals who lived at thermoneutrality but ate in the cold (versus eating at the same Ta) [25]. These results demonstrate the significant influence acute Ta exposure can have during food intake periods, but whether similar influences of acute cold exposure during dining would result in greater weight gain in humans requires further investigation.
Ambient Temperature and Diet Induced Thermogenesis
The thermic effect of feeding, or diet induced thermogenesis – DIT, following the ingestion of calories is associated with increases in metabolism and skin temperature [26;27]. Obligatory DIT represents the processes necessary for digestion, absorption, etc. that are intimately associated with the caloric content and form of the food [28–30]. Facultative DIT refers to an adaptive thermogenic response where calories are converted to heat energy through metabolic processes like mitochondrial uncoupling [28;29]. Ta influences both the caloric intake and the thermogenic potential of DIT by increasing (low Ta) or decreasing (high Ta) the thermal sink of the environment in relationship to body temperature. Thus, a greater DIT response is possible in reduced Ta conditions [26]. At Ta above the TNZ, food intake is further depressed, potentially resulting in negative energy balance due to the associated energy expenditure increase. Multiple reports have demonstrated potential differences in DIT between lean and obese subjects. One difficulty with these studies is the interpretation of any measured deficit of DIT as causative or correlative with obesity. Consider for instance, DIT was significantly lower in obese versus lean men when comparing individuals with similar lean body mass [31]. Furthermore, obese subjects were shown to have an initial DIT deficit compared with normal weight controls, which was partially recovered after weight reduction by gastric banding [32]. Equally intriguing is a study where an artificial form of obesity was applied. Comparing lean and obese subjects, a significant deficit of DIT was observed in obese individuals [33]. However, the application of a physical, insulative barrier to the greater abdominal surface area of the lean subjects by wrapping them in a thermal blanket resulted in a significant suppression of DIT, demonstrating what the authors suggest is a leakage of heat across the abdominal wall relevant to the extent of DIT [33]. Considering these results together it appears that an established obese phenotype, whether adiposity or artificial in nature, has significant implications for energy dissipation related to DIT. Thus, the altered morphologic characteristics of obese individuals with their large, insulative barrier of adipose tissue as assessed by lower abdominal skin surface temperature measurements compared with core and peripheral skin temperatures [34;35], may physically limit the amount of heat dissipation and blunt potential DIT responses to protect against a core hyperthermic response.
Yet, this does not rule out the possibility that individuals with reduced thermogenic capacity (through defective BAT or mitochondrial energetic economy [36]) could be biased toward a greater energetic efficiency and weight gain under specific temperature and diet conditions. In fact, more recent work in animal models has demonstrated the interdependence of metabolic potential and Ta in body weight gain and obesity related phenotypes. Feldmann et al 2009 addressed a puzzling result in the thermogenesis field by assessing why brown adipose tissue (BAT) defective rodents were normal body weight – not obese as expected [37]. By using uncoupling protein 1 (UCP1) ablated mice and controls, their work demonstrated that a low Ta induces a thermic response that must be achieved metabolically either through BAT uncoupling or other mechanisms in a BAT defective state [37]. However, protection from thermal stress by thermoneutral housing resulted in increased energetic efficiency with attenuated DIT, resulting in accelerated weight gain and obesity induction in BAT defective mice even when fed a normal chow (low fat) diet [37]. They concluded that “ambient temperature is qualitatively determinative for the outcome of metabolic studies, that no other protein and no other mechanism can substitute for UCP1 in mediating diet-induced adrenergic thermogenesis, and that UCP1 activity can be determinative for obesity development in mice and possibly in humans” [37]. Bearing in mind the limitations of translating animal results to human phenotypes, this study would suggest that individuals with defective BAT function may be more susceptible to excess weight gain under thermoneutral conditions, while semi-protected at a low Ta due to alternate mechanisms for cold-induced, elevated metabolism.
Ambient Temperature and Brown Adipose Tissue
In rodents, BAT is known to function in both cold-induced, non-shivering thermogenesis (NST) and facultative DIT [11;29;37]. By manipulations of Ta, BAT depots can be either expanded (cold Ta) or repressed (warm Ta) with SNS induced activity directly related to the activation potential of the tissue [11]. Adaptive responses in BAT function can be observed over a 1–2 week period upon shifting to a different Ta [11]. Additionally, a strong association between NST and DIT potential is present due to their shared mechanisms of action [11]. Even in the absence of functional BAT, cold-induced increases in energy expenditure are expected and observed due to other mechanisms of increasing energy expenditure (e.g. shivering, etc.) [38]. However, the facultative component of DIT may be exclusive to BAT and thus is irreplaceable in its function of dissipating excess caloric intake as heat [29;37]. As such, animals, and presumably people, with defective or suppressed BAT function may be expected to be more susceptible to weight gain during excess caloric intake [30;37;39;40]. Additionally, the suppression of BAT function in thermoneutral conditions would predict a decreased DIT. This energetic impairment would be expected to equally affect both lean and obese individuals, independent of BAT status, and could theoretically contribute to weight or fat gain in thermoneutral conditions. In human subjects, large differences in DIT between individuals and under different conditions are often observed [28;41–43]. By measuring the thermogenic response to mild cold and overfeeding in humans, a highly correlated individual response to both stimuli was observed and was related to norepinepherine levels [41;44]. Whether a shared mechanism of BAT was responsible for the SNS-related response in human subjects remains to be demonstrated.
Since 2009 a number of groups have utilized positron emission tomography and computed tomography (PET/CT) with 2-fluorodeoxyglucose label to demonstrate a metabolically active adipose tissue located in the cervical and shoulder region of healthy adults. Biopsies of tissue from these sites possess brown adipose tissue (BAT) histological and biochemical markers [45–47]. Retrospective PET/CT studies of patients undergoing diagnostic scans for cancer had indicated the likelihood of BAT presence, showing BAT incidence associated with younger age, lower BMI and acute Ta influence (cold temperature activated) [48–51]. Previous anatomic and histologic observations in human cadavers had demonstrated cells with brown adipose like characteristics including multi-locular lipid droplets, numerous dense mitochondria and rich vascular supply [52–56]. In conditions with expected high sympathetic activity, depletion of lipid content of the brown adipose tissue was considered indirect evidence of activity and regulation [54;57;58]. Despite these earlier reports, a lack of capable technologies for identifying and studying BAT in vivo had hindered progress in determining the significance of BAT in humans, although it was proposed as a potential mediator of energetic balance [59;60]. Greater than 90% of healthy, young adults that were studied exhibited inducible BAT activity under cold stimulation that was inversely related to BMI and percent body fat [46], with other studies estimating ~60% of the total population as BAT positive [45;61–63]. While BAT is clearly protective against obesity and diet-related metabolic impairment (and compromised in models of obesity and diabetes) in animal models [64], the clinical relevance in humans remains to be demonstrated [60;65]. As additional technologies are developed to measure BAT quantity and activity in vivo [66–68], future studies will hopefully clarify the role, if any, BAT may play in body weight regulation and disease prevention.
Ambient Temperature and Thermoneutral Zone Estimation
With the advent of climate control through heat and air conditioning, secular trends for a reduced exposure to variability in Ta and a greater proportion of time spent in the TNZ has been proposed as a contributor to energetic efficiency and weight gain/obesity due to reduced cold- or heat-induced metabolic responses [4–7]. Regarding the TNZ, it is worth noting that many of the studies that have measured the metabolic response to different Tas may not be representative of diverse human populations and living environments. This is in part because multiple factors influence temperature perception including air speed, humidity, age, body size, body fat, clothing, acclimation, diet, activity, etc. [69]. We have all utilized some of these techniques to buffer against a temperature response (e.g. adding a coat during the winter, turning on fans during the summer). Therefore, a TNZ range can be most narrowly defined in a homogenous population of individuals where each of these factors is well controlled. Furthermore, physiologic, anthropometric and morphologic differences between normal weight and obese subjects may be of importance as the additional insulation provided by layers of adiposity should be protective against cold temperatures [70], but potentially disadvantageous during heat stress conditions [71;72]. Therefore, as with body temperature, measurement variability between subjects and at the site of measure within individuals [73], it would appear necessary to acknowledge that no single Ta can be considered definitive as the point of thermoneutrality for all subjects, thus, physiologic and metabolic responses will likely differ between subjects exposed to different Tas.
While recent findings have reinvigorated an interest in thermal regulation and body weight gain, Ta within the range experienced in daily activities in developed countries has been proposed as a potential contributor to energy balance for some time [74]. After demonstrating a greater 24 hour heat production at 22°C compared with 28°C it was concluded that “(a) environmental temperature may play a more important role than was previously recognized in the energy balance…[indicating] that mild cold should at least be considered as a factor which can affect the energy metabolism of people” [74]. While a thermogenic response may be expected under cold stimulated conditions, other physiologic responses (e.g. peripheral vasoconstriction) and behavioral modifications (e.g. clothing choice) could protect against small cold challenges, reducing any needed thermogenic response. When subjects were allowed clothing choice and energy expenditure was measured at 28°C and 20°C, cooler temperature was still associated with increased expenditure despite increased usage of clothing and bedding [75]. To what extent the selected clothing buffered the cold challenge within subjects was not measurable. Interestingly, 2 of 3 obese subjects demonstrated a lower than expected expenditure under the cooler condition, suggesting their basal expenditure level associated with thermoneutrality was potentially shifted toward a lower Ta under the tested conditions, although obese subjects as a group were not significantly different than lean subjects [75].
When measuring energy expenditure and energy intake in young, adult, healthy, normal body weight subjects, a continual response of increasing metabolic rate with decreasing Ta (even within the range of temperature normally encountered over a 24-hour period), accompanied by an increased energy intake in cooler temperatures when fed ad libitum, is observed [26;76–78]. In response to this mild cold, sleeping metabolic rate (SMR) and diet induced thermogenesis were both increased, with diet induced thermogenesis comprising a proportionally larger percentage of total daily energy expenditure [26]. This is particularly interesting as SMR would represent an individual’s metabolic capacity in the absence of either food intake or physical activity, and might demonstrate greater energy expenditure during sleep in some individuals. An increased metabolic response to cold has been reported in winter [79;80], and sometimes in summer [79], with peripheral vasoconstriction as measured by core minus distal skin temperature related to total energy expenditure [79]. This highlights the seasonal effects on metabolism and various ways that individuals respond to cold stress (insulative:vasoconstriction vs. metabolic:thermogenesis), which the authors point out could have potential implications for subject specific energy balance [79]. A divergent response between lean and obese women to Ta is observed with heat loss being decreased at cool Ta in obese subjects (−2.1%) and increased in lean subjects (+7.8%; but not statistically significant) [81]. When comparing lean and obese men subjected to cooling, both groups had an increase in metabolism, but the relative increase was significantly larger in lean subjects [34]. Whether this is because of an innate defect in cold-induced thermogenesis or a blunted response due to the insulative protection of adiposity is not clearly delineated. Nevertheless, it is clear that while energy expenditure normally increases during cold exposure, differences between individuals of different body compositions and thermogenic capacities may dispose them to a greater or lesser sensitivity and metabolic response.
Ambient Temperature Reduction in Lean Versus Obese Subjects
Reduced Ambient Temperature
Consider a scenario of two individuals of similar age but vastly different body composition (one lean with a low percentage body fat and the other obese with a high percentage body fat) exposed to the same environmental temperatures. As discussed previously, the known effects of cold are an increased metabolic rate and food intake. Considering a thermoneutral temperature of ~22°C for the lean, clothed individual, a shift to 16°C would result in a cold stress met by an increased sympathetic nervous system response and increased thermogenesis. This would include an increase in peripheral vasoconstriction, heart rate, blood pressure, circulating fatty acids, insulin, thyroxine, cortisol [82], with stimulation of brown adipose tissue and increased food intake. In contrast, an obese individual exposed to 16°C would still be expected to mount a SNS response and small thermogenic response; however, the insulative properties of the excess adipose tissue would result in a suppressed cold response, as the obese individual would be closer to the lower limit of thermoneutrality or possibly within the range. The obese individual would demonstrate little or no activated BAT, but thermogenesis, if needed, may originate from other tissues (e.g. muscle). Additionally, the obese individual could increase food intake, but with a blunted DIT response relative to lean individuals [27;31–33;83]. Considering the presence of BAT in lean individuals and its relative limitation or absence in obese individuals, the adaptive/facultative DIT possible in BAT would be significantly greater in the lean individual than the obese. Thus, a lower Ta would be expected to be “protective” against the development of a positive energy balance and weight gain for the lean individual, while low Ta could be metabolically insignificant or a potential contributor to a greater energetic efficiency of the obese individual, predisposing them to additional weight gain. Whether this could result in a positive feedback scenario where increased metabolic efficiency with increased body mass/fat mass at lower Ta biases to excess energy storage and additional weight gain needs to be further explored.
Elevated Ambient Temperature
In the case of a lean individual exposed to a high Ta, metabolism would be increased while food intake would be decreased. In short term studies this has resulted in a negative energy balance and body weight and fat loss. Similarly, in an obese individual, exposure to Ta above the TNZ (which we argue could be shifted to a lower Ta upper limit due to increased adiposity) would also increase metabolism as processes are activated to increase heat dissipation against a smaller thermal gradient. Food intake would be reduced, resulting in a negative energy balance and body weight/fat loss. In both of these “hyperthermic” conditions, the relative contribution of DIT to energy expenditure (and energy balance) would be reduced as food intake is decreased and thermogenic mechanisms like BAT would be suppressed during any chronic heat exposure acclimation.
Finally, consider the separation of Ta during meals versus daily activities and sleep/rest. Since acute exposure to Ta may increase energy intake, if meals were consumed in cool conditions followed by a transition to a warmer Ta, an unfortunate occurrence of mismatched intake and expenditure could occur. Whether this would result in excess weight gain in humans, and if obese individuals with suppressed DIT would be more susceptible could be explored in the future.
Ambient Temperature and Animal Models – Cautions & Considerations
The effects and interactions of Ta, nutrition and weight gain may be most highly researched in animal models including rodents, chickens and swine. While each demonstrate Ta-dependent alterations in energy expenditure and intake [12;84;85], the applicability for predicting human responses suffer from a few limitations. Of relevance to cold- and diet-induced thermogenesis, both pigs and chickens possess BAT-like cells, but lack a functional uncoupling protein 1 gene [86;87]. With the reinvigorated interest in the significance of BAT and its contribution to energy expenditure under cold stimulation and diet conditions, caution should be applied in over-generalizing results from both of these organisms to humans, though their utility as a model of impaired BAT function may yet be exploited. While most rodents possess BAT, their surface area to volume (or mass) ratio is significantly larger than with humans, resulting in a significantly greater thermal challenge under a given Ta exposure [11;12;29]. This is evidenced by the proportionally greater BAT mass relative to total body weight (~10 fold large in mice than humans) which is associated with a significantly greater metabolic increase in response to SNS BAT stimulation [11;12]. However, thermoneutral housing may partially alleviate this cold-induced effect and improve the utility of the model for translational research [29;88].
Conclusion
Ambient temperature can clearly influence both energy intake and energy expenditure, even within the range of Ta encountered in modern society. Additionally, time spent in the thermoneutral zone could be a contributor to greater energetic efficiency (decreased metabolic rate) and weight gain. Although conclusive data is still needed, it is not clear that lowering Ta would be effective in treating established obesity. Rather, current data support the hypothesis that an increased energy intake in response to mild cold, with a decreased DIT in obese subjects, could contribute to a positive energy balance and greater weight gain under mild cold conditions.. Although it is well recongnized that large inter-individual differences in metabolic rate exist, a systematic assessment of metabolic responses to a range of Tas may be informative for indentifying individuals who are more “energetically efficient” and thus susceptible to excess weight gain. Specifically, measures of diet-induced thermogenesis, activity-related thermogenesis and sleeping metabolic rate at temperatures below, above and within the TNZ may provide a more relevant potrait of the daily cycles of life as compared with the imposed, artificial environments of clinical laboratory facilities. Future questions that will contribute to our understanding of Ta and obesity include:
What is the metabolic response of lean versus obese subjects across a broad range of ambient temperatures? Is the TNZ simply shifted lower or expanded in the obese state?
Can lowering Ta shift energy balance sufficiently to induce weight loss in the absence of dietary control in humans? Could raising Ta above the TNZ be used as a dietary suppressant, weight-loss regimen?
Are the physiologic and metabolic responses to Ta above and below the TNZ a risk factor for cardiovascular complications?
What is the free living, 24-hour Ta exposure profile for humans in modern society? Can reduced Ta at night be utilized to increase energy expenditure and stimulate thermogenic potential for energy balance and weight maintenance? How do clothing, bedding, activity and other variables influence these exposures? How do Ta exposures change with the seasons?
Could intermittent, acute exposures to cold Ta (outside of meal times) provide sufficient stimulus to promote prolonged energy expenditure and brown adipose tissue expansion or is chronic exposure necessary?
Do Ta exposures during growth and development influence long-term metabolism and weight gain in humans?
While more research is needed, it is becoming increasingly apparent that ambient temperature can have significant impacts on our energy expenditure, energy intake and thus ultimately our energy balance. As a return to varied, natural ingredient diets is proposed for health, so too exposure to varied, natural range of ambient temperatures may be just what we need.
Reference List
- 1.Olshansky SJ, Passaro DJ, Hershow RC, Layden J, Carnes BA, Brody J, Hayflick L, Butler RN, Allison DB, Ludwig DS. A potential decline in life expectancy in the United States in the 21st century. N Engl J Med. 2005;352:1138–1145. doi: 10.1056/NEJMsr043743. [DOI] [PubMed] [Google Scholar]
- 2.Yanovski SZ, Yanovski JA. Obesity prevalence in the United States--up, down, or sideways? N Engl J Med. 2011;364:987–989. doi: 10.1056/NEJMp1009229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Flegal KM, Carroll MD, Ogden CL, Curtin LR. Prevalence and trends in obesity among US adults, 1999–2008. JAMA. 2010;303:235–241. doi: 10.1001/jama.2009.2014. [DOI] [PubMed] [Google Scholar]
- 4.Keith SW, Redden DT, Katzmarzyk PT, Boggiano MM, Hanlon EC, Benca RM, Ruden D, Pietrobelli A, Barger JL, Fontaine KR, Wang C, Aronne LJ, Wright SM, Baskin M, Dhurandhar NV, Lijoi MC, Grilo CM, DeLuca M, Westfall AO, Allison DB. Putative contributors to the secular increase in obesity: exploring the roads less traveled. Int J Obes(Lond) 2006;30:1585–1594. doi: 10.1038/sj.ijo.0803326. [DOI] [PubMed] [Google Scholar]
- 5.McAllister EJ, Dhurandhar NV, Keith SW, Aronne LJ, Barger J, Baskin M, Benca RM, Biggio J, Boggiano MM, Eisenmann JC, Elobeid M, Fontaine KR, Gluckman P, Hanlon EC, Katzmarzyk P, Pietrobelli A, Redden DT, Ruden DM, Wang C, Waterland RA, Wright SM, Allison DB. Ten putative contributors to the obesity epidemic. Crit Rev Food Sci Nutr. 2009;49:868–913. doi: 10.1080/10408390903372599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Johnson F, Mavrogianni A, Ucci M, Vidal-Puig A, Wardle J. Could increased time spent in a thermal comfort zone contribute to population increases in obesity? Obes Rev. 2011;12:543–551. doi: 10.1111/j.1467-789X.2010.00851.x. [DOI] [PubMed] [Google Scholar]
- 7.Hansen JC, Gilman AP, Odland JO. Is thermogenesis a significant causal factor in preventing the “globesity” epidemic? Med Hypotheses. 2010;75:250–256. doi: 10.1016/j.mehy.2010.02.033. [DOI] [PubMed] [Google Scholar]
- 8.Weinsier RL, Bracco D, Schutz Y. Predicted effects of small decreases in energy expenditure on weight gain in adult women. Int J Obes Relat Metab Disord. 1993;17:693–700. [PubMed] [Google Scholar]
- 9.Katan MB, Ludwig DS. Extra calories cause weight gain--but how much? JAMA. 2010;303:65–66. doi: 10.1001/jama.2009.1912. [DOI] [PubMed] [Google Scholar]
- 10.Benzinger TH. Heat regulation: homeostasis of central temperature in man. Physiol Rev. 1969;49:671–759. doi: 10.1152/physrev.1969.49.4.671. [DOI] [PubMed] [Google Scholar]
- 11.Cannon B, Nedergaard J. Brown adipose tissue: function and physiological significance. Physiol Rev. 2004;84:277–359. doi: 10.1152/physrev.00015.2003. [DOI] [PubMed] [Google Scholar]
- 12.Chaffee RR, Roberts JC. Temperature acclimation in birds and mammals. Annu Rev Physiol. 1971;33:155–202. doi: 10.1146/annurev.ph.33.030171.001103. [DOI] [PubMed] [Google Scholar]
- 13.DAVIS TR. THE INFLUENCE OF CLIMATE ON NUTRITIONAL REQUIREMENTS. Am J Public Health Nations Health. 1964;54:2051–2067. doi: 10.2105/ajph.54.12.2051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.HERRINGTON LP. The heat regulation of small laboratory animals at various environmental temperatures. Am J Physiol. 1940;129:123–139. [Google Scholar]
- 15.ERIKSON H, KROG J, ANDERSEN KL, Scholander PF. The critical temperature in naked man. Acta Physiol Scand. 1956;37:35–39. doi: 10.1111/j.1748-1716.1956.tb01339.x. [DOI] [PubMed] [Google Scholar]
- 16.Brobeck JR. Food intake as a mechanism of temperature regulation. Yale J Biol Med. 1948;20:545–552. [PMC free article] [PubMed] [Google Scholar]
- 17.Herman CP. Effects of Heat on Appetite. In: Marriott BM, editor. Nutrional Needs in Hot Environments. 1993. pp. 187–214. [Google Scholar]
- 18.Yates AA. Food Intake, Appetite, and Work in Hot Environments. In: Marriott BM, editor. Nutritional Needs in Hot Environments. 1993. pp. 297–304. [Google Scholar]
- 19.Keys A, Brozek J, Henschel A, Mickelsen O, Taylor HL. The Biology of Human Starvation. University of Minnesota Press; 1950. [Google Scholar]
- 20.Westerterp-Plantenga MS. Effects of extreme environments on food intake in human subjects. Proc Nutr Soc. 1999;58:791–798. doi: 10.1017/s002966519900107x. [DOI] [PubMed] [Google Scholar]
- 21.Chaffee RR, Kaufman WC, Kratochvil CH, Sorenson MW, Conaway CH, Middleton CC. Comparative chemical thermoregulation in cold- and heat-acclimated rodents, insectivores, protoprimates, and primates. Fed Proc. 1969;28:1029–1034. [PubMed] [Google Scholar]
- 22.Chaffee RR, Allen JR. Effects of ambient temperature on the resting metabolic rate of cold- and heat-acclimated Macaca mulatta. Comp Biochem Physiol A. 1973;44:1215–1225. doi: 10.1016/0300-9629(73)90260-0. [DOI] [PubMed] [Google Scholar]
- 23.Jakubczak LF. Food and water intakes of rats as a function of strain, age, temperature, and body weight. Physiol Behav. 1976;17:251–258. doi: 10.1016/0031-9384(76)90073-1. [DOI] [PubMed] [Google Scholar]
- 24.Stroebele N, De Castro JM. Effect of ambience on food intake and food choice. Nutrition. 2004;20:821–838. doi: 10.1016/j.nut.2004.05.012. [DOI] [PubMed] [Google Scholar]
- 25.Refinetti R. Effects of food temperature and ambient temperature during a meal on food intake in the rat. Physiol Behav. 1988;43:245–247. doi: 10.1016/0031-9384(88)90247-8. [DOI] [PubMed] [Google Scholar]
- 26.Westerterp-Plantenga MS, van Marken Lichtenbelt WD, Strobbe H, Schrauwen P. Energy metabolism in humans at a lowered ambient temperature. Eur J Clin Nutr. 2002;56:288–296. doi: 10.1038/sj.ejcn.1601308. [DOI] [PubMed] [Google Scholar]
- 27.Booth G, Strang JM. Changes in temperature of the skin following ingestion of food. Arch Intern Med. 1936;57:533–543. [Google Scholar]
- 28.Himms-Hagen J. Role of thermogenesis in the regulation of energy balance in relation to obesity. Can J Physiol Pharmacol. 1989;67:394–401. doi: 10.1139/y89-063. [DOI] [PubMed] [Google Scholar]
- 29.Cannon B, Nedergaard J. Nonshivering thermogenesis and its adequate measurement in metabolic studies. J Exp Biol. 2011;214:242–253. doi: 10.1242/jeb.050989. [DOI] [PubMed] [Google Scholar]
- 30.Jequier E. Thermogenic responses induced by nutrients in man: their importance in energy balance regulation. Experientia Suppl. 1983;44:26–44. doi: 10.1007/978-3-0348-6540-1_3. [DOI] [PubMed] [Google Scholar]
- 31.Segal KR, Gutin B, Albu J, Pi-Sunyer FX. Thermic effects of food and exercise in lean and obese men of similar lean body mass. Am J Physiol. 1987;252:E110–E117. doi: 10.1152/ajpendo.1987.252.1.E110. [DOI] [PubMed] [Google Scholar]
- 32.Thorne A, Hallberg D, Wahren J. Meal-induced thermogenesis in obese patients before and after weight reduction. Clin Physiol. 1989;9:481–498. doi: 10.1111/j.1475-097x.1989.tb01002.x. [DOI] [PubMed] [Google Scholar]
- 33.Brundin T, Thorne A, Wahren J. Heat leakage across the abdominal wall and meal-induced thermogenesis in normal-weight and obese subjects. Metabolism. 1992;41:49–55. doi: 10.1016/0026-0495(92)90190-l. [DOI] [PubMed] [Google Scholar]
- 34.Claessens-van Ooijen AM, Westerterp KR, Wouters L, Schoffelen PF, van Steenhoven AA, van Marken Lichtenbelt WD. Heat production and body temperature during cooling and rewarming in overweight and lean men. Obesity (Silver Spring) 2006;14:1914–1920. doi: 10.1038/oby.2006.223. [DOI] [PubMed] [Google Scholar]
- 35•.Savastano DM, Gorbach AM, Eden HS, Brady SM, Reynolds JC, Yanovski JA. Adiposity and human regional body temperature. Am J Clin Nutr. 2009;90:1124–1131. doi: 10.3945/ajcn.2009.27567. Demonstration by infrared thermography of the altered thermal profile of lean and obese individuals exposed to the same ambient temperature conditions. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Krzywanski DM, Moellering DR, Fetterman JL, Dunham-Snary KJ, Sammy MJ, Ballinger SW. The mitochondrial paradigm for cardiovascular disease susceptibility and cellular function: a complementary concept to Mendelian genetics. Lab Invest. 2011;91:1122–1135. doi: 10.1038/labinvest.2011.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37•.Feldmann HM, Golozoubova V, Cannon B, Nedergaard J. UCP1 ablation induces obesity and abolishes diet-induced thermogenesis in mice exempt from thermal stress by living at thermoneutrality. Cell Metab. 2009;9:203–209. doi: 10.1016/j.cmet.2008.12.014. By housing mice within their thermoneutral zone, a defect in thermogenesis reveals an increased energetic efficiency and propensity to increased weight gain and fat accrual. Thus, a gene by environment interaction disposes toward excess weight gain and obesity phenotypes only in the absence of cold stress. [DOI] [PubMed] [Google Scholar]
- 38.Liu X, Rossmeisl M, McClaine J, Riachi M, Harper ME, Kozak LP. Paradoxical resistance to diet-induced obesity in UCP1-deficient mice. J Clin Invest. 2003;111:399–407. doi: 10.1172/JCI15737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Himms-Hagen J. Impaired thermogenesis and brown fat in obesity. Can J Surg. 1984;27:125. [PubMed] [Google Scholar]
- 40.Rothwell NJ, Stock MJ. A role for brown adipose tissue in diet-induced thermogenesis. Obes Res. 1997;5:650–656. doi: 10.1002/j.1550-8528.1997.tb00591.x. [DOI] [PubMed] [Google Scholar]
- 41.Wijers SL, Saris WH, van Marken Lichtenbelt WD. Individual thermogenic responses to mild cold and overfeeding are closely related. J Clin Endocrinol Metab. 2007;92:4299–4305. doi: 10.1210/jc.2007-1065. [DOI] [PubMed] [Google Scholar]
- 42.Stock MJ. Gluttony and thermogenesis revisited. Int J Obes Relat Metab Disord. 1999;23:1105–1117. doi: 10.1038/sj.ijo.0801108. [DOI] [PubMed] [Google Scholar]
- 43.van Marken Lichtenbelt WD, Schrauwen P, van De KS, Westerterp-Plantenga MS. Individual variation in body temperature and energy expenditure in response to mild cold. Am J Physiol Endocrinol Metab. 2002;282:E1077–E1083. doi: 10.1152/ajpendo.00020.2001. [DOI] [PubMed] [Google Scholar]
- 44.Wijers SL, Saris WH, van Marken Lichtenbelt WD. Recent advances in adaptive thermogenesis: potential implications for the treatment of obesity. Obes Rev. 2008 doi: 10.1111/j.1467-789X.2008.00538.x. [DOI] [PubMed] [Google Scholar]
- 45•.Virtanen KA, Lidell ME, Orava J, Heglind M, Westergren R, Niemi T, Taittonen M, Laine J, Savisto NJ, Enerback S, Nuutila P. Functional brown adipose tissue in healthy adults. N Engl J Med. 2009;360:1518–1525. doi: 10.1056/NEJMoa0808949. Documents the presence of metabolically active brown adipose tissue in healthy adult humans by PET/CT scans. [DOI] [PubMed] [Google Scholar]
- 46•.van Marken Lichtenbelt WD, Vanhommerig JW, Smulders NM, Drossaerts JM, Kemerink GJ, Bouvy ND, Schrauwen P, Teule GJ. Cold-activated brown adipose tissue in healthy men. N Engl J Med. 2009;360:1500–1508. doi: 10.1056/NEJMoa0808718. Documents the presence of metabolically active brown adipose tissue in healthy adult humans by PET/CT scans using a mild cold ambient temperature exposure. Inverse associations of BAT activity with body weight and body fat are reported. [DOI] [PubMed] [Google Scholar]
- 47.Saito M, Okamatsu-Ogura Y, Matsushita M, Watanabe K, Yoneshiro T, Nio-Kobayashi J, Iwanaga T, Miyagawa M, Kameya T, Nakada K, Kawai Y, Tsujisaki M. High Incidence of Metabolically Active Brown Adipose Tissue in Healthy Adult Humans: Effects of Cold Exposure and Adiposity. Diabetes. 2009 doi: 10.2337/db09-0530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hany TF, Gharehpapagh E, Kamel EM, Buck A, Himms-Hagen J, von Schulthess GK. Brown adipose tissue: a factor to consider in symmetrical tracer uptake in the neck and upper chest region. Eur J Nucl Med Mol Imaging. 2002;29:1393–1398. doi: 10.1007/s00259-002-0902-6. [DOI] [PubMed] [Google Scholar]
- 49.Nedergaard J, Bengtsson T, Cannon B. Unexpected evidence for active brown adipose tissue in adult humans. Am J Physiol Endocrinol Metab. 2007;293:E444–E452. doi: 10.1152/ajpendo.00691.2006. [DOI] [PubMed] [Google Scholar]
- 50.Cypess AM, Lehman S, Williams G, Tal I, Rodman D, Goldfine AB, Kuo FC, Palmer EL, Tseng YH, Doria A, Kolodny GM, Kahn CR. Identification and importance of brown adipose tissue in adult humans. N Engl J Med. 2009;360:1509–1517. doi: 10.1056/NEJMoa0810780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Nedergaard J, Bengtsson T, Cannon B. Three years with adult human brown adipose tissue. Ann NY Acad Sci. 2010;1212:E20–E36. doi: 10.1111/j.1749-6632.2010.05905.x. [DOI] [PubMed] [Google Scholar]
- 52.Heaton JM. The distribution of brown adipose tissue in the human. J Anat. 1972;112:35–39. [PMC free article] [PubMed] [Google Scholar]
- 53.Tanuma Y, Tamamoto M, Ito T, Yokochi C. The occurrence of brown adipose tissue in perirenal fat in Japanese. Arch Histol Jpn. 1975;38:43–70. doi: 10.1679/aohc1950.38.43. [DOI] [PubMed] [Google Scholar]
- 54.Tanuma Y, Ohata M, Ito T, Yokochi C. Possible function of human brown adipose tissue as suggested by observation on perirenal brown fats from necropsy cases of variable age groups. Arch Histol Jpn. 1976;39:117–145. doi: 10.1679/aohc1950.39.117. [DOI] [PubMed] [Google Scholar]
- 55.Nnodim JO. The occurrence of brown adipose in man inhabiting the tropics. Z Mikrosk Anat Forsch. 1990;104:721–728. [PubMed] [Google Scholar]
- 56.Lever JD, Jung RT, Nnodim JO, Leslie PJ, Symons D. Demonstration of a catecholaminergic innervation in human perirenal brown adipose tissue at various ages in the adult. Anat Rec. 1986;215:251–259. doi: 10.1002/ar.1092150307. [DOI] [PubMed] [Google Scholar]
- 57.Huttunen P, Hirvonen J, Kinnula V. The occurrence of brown adipose tissue in outdoor workers. Eur J Appl Physiol Occup Physiol. 1981;46:339–345. doi: 10.1007/BF00422121. [DOI] [PubMed] [Google Scholar]
- 58.Huttunen P, Kortelainen ML. Long-term alcohol consumption and brown adipose tissue in man. Eur J Appl Physiol Occup Physiol. 1990;60:418–424. doi: 10.1007/BF00705030. [DOI] [PubMed] [Google Scholar]
- 59.Himms-Hagen J. Obesity may be due to a malfunctioning of brown fat. Can Med Assoc J. 1979;121:1361–1364. [PMC free article] [PubMed] [Google Scholar]
- 60.Himms-Hagen J. Does brown adipose tissue (BAT) have a role in the physiology or treatment of human obesity? Rev Endocr Metab Disord. 2001;2:395–401. doi: 10.1023/a:1011856617047. [DOI] [PubMed] [Google Scholar]
- 61.Nedergaard J, Bengtsson T, Cannon B. Three years with adult human brown adipose tissue. Ann NY Acad Sci. 2010;1212:E20–E36. doi: 10.1111/j.1749-6632.2010.05905.x. [DOI] [PubMed] [Google Scholar]
- 62.Orava J, Nuutila P, Lidell ME, Oikonen V, Noponen T, Viljanen T, Scheinin M, Taittonen M, Niemi T, Enerback S, Virtanen KA. Different metabolic responses of human brown adipose tissue to activation by cold and insulin. Cell Metab. 2011;14:272–279. doi: 10.1016/j.cmet.2011.06.012. [DOI] [PubMed] [Google Scholar]
- 63.Lee P, Greenfield JR, Ho KK, Fulham MJ. A critical appraisal of the prevalence and metabolic significance of brown adipose tissue in adult humans. Am J Physiol Endocrinol Metab. 2010;299:E601–E606. doi: 10.1152/ajpendo.00298.2010. [DOI] [PubMed] [Google Scholar]
- 64.Cannon B, Nedergaard J. Metabolic consequences of the presence or absence of the thermogenic capacity of brown adipose tissue in mice (and probably in humans) Int J Obes(Lond) 2010;34 (Suppl 1):S7–16. doi: 10.1038/ijo.2010.177. [DOI] [PubMed] [Google Scholar]
- 65.Nedergaard J, Bengtsson T, Cannon B. Three years with adult human brown adipose tissue. Ann NY Acad Sci. 2010;1212:E20–E36. doi: 10.1111/j.1749-6632.2010.05905.x. [DOI] [PubMed] [Google Scholar]
- 66.Hu HH, Smith DL, Jr, Nayak KS, Goran MI, Nagy TR. Identification of brown adipose tissue in mice with fat-water IDEAL-MRI. J Magn Reson Imaging. 2010;31:1195–1202. doi: 10.1002/jmri.22162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Hamilton G, Smith DL, Jr, Bydder M, Nayak KS, Hu HH. MR properties of brown and white adipose tissues. J Magn Reson Imaging. 2011;34:468–473. doi: 10.1002/jmri.22623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Branca RT, Warren WS. In vivo brown adipose tissue detection and characterization using water-lipid intermolecular zero-quantum coherences. Magn Reson Med. 2011;65:313–319. doi: 10.1002/mrm.22622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Wagner JA, Robinson S, Marino RP. Age and temperature regulation of humans in neutral and cold environments. J Appl Physiol. 1974;37:562–565. doi: 10.1152/jappl.1974.37.4.562. [DOI] [PubMed] [Google Scholar]
- 70.Anderson GS. Human morphology and temperature regulation. Int J Biometeorol. 1999;43:99–109. doi: 10.1007/s004840050123. [DOI] [PubMed] [Google Scholar]
- 71.BUSKIRK ER, Lundegren H, Magnusson L. Heat acclimatization patterns in obese and lean individuals. Ann NY Acad Sci. 1965;131:637–653. doi: 10.1111/j.1749-6632.1965.tb34827.x. [DOI] [PubMed] [Google Scholar]
- 72.Chung NK, Pin CH. Obesity and the occurrence of heat disorders. Mil Med. 1996;161:739–742. [PubMed] [Google Scholar]
- 73.Livingstone SD, Grayson J, Frim J, Allen CL, Limmer RE. Effect of cold exposure on various sites of core temperature measurements. J Appl Physiol. 1983;54:1025–1031. doi: 10.1152/jappl.1983.54.4.1025. [DOI] [PubMed] [Google Scholar]
- 74.Dauncey MJ. Influence of mild cold on 24 h energy expenditure, resting metabolism and diet-induced thermogenesis. Br J Nutr. 1981;45:257–267. doi: 10.1079/bjn19810102. [DOI] [PubMed] [Google Scholar]
- 75.Warwick PM, Busby R. Influence of mild cold on 24 h energy expenditure in ‘normally’ clothed adults. Br J Nutr. 1990;63:481–488. doi: 10.1079/bjn19900135. [DOI] [PubMed] [Google Scholar]
- 76.Westerterp-Plantenga MS, van Marken Lichtenbelt WD, Cilissen C, Top S. Energy metabolism in women during short exposure to the thermoneutral zone. Physiol Behav. 2002;75:227–235. doi: 10.1016/s0031-9384(01)00649-7. [DOI] [PubMed] [Google Scholar]
- 77.Wilkerson JE, Raven PB, Horvath SM. Critical temperature of unacclimatized male Caucasians. J Appl Physiol. 1972;33:451–455. doi: 10.1152/jappl.1972.33.4.451. [DOI] [PubMed] [Google Scholar]
- 78.Yoshimura M, Yoshimura H. Cold tolerance and critical temperature of the Japanese. Int J Biometeorol. 1969;13:163–172. doi: 10.1007/BF01552737. [DOI] [PubMed] [Google Scholar]
- 79.van Ooijen AM, van Marken Lichtenbelt WD, van Steenhoven AA, Westerterp KR. Seasonal changes in metabolic and temperature responses to cold air in humans. Physiol Behav. 2004;82:545–553. doi: 10.1016/j.physbeh.2004.05.001. [DOI] [PubMed] [Google Scholar]
- 80.Kashiwazaki H, Dejima Y, Suzuki T. Influence of upper and lower thermoneutral room temperatures (20 degrees C and 25 degrees C) on fasting and post-prandial resting metabolism under different outdoor temperatures. Eur J Clin Nutr. 1990;44:405–413. [PubMed] [Google Scholar]
- 81.Blaza S, Garrow JS. Thermogenic response to temperature, exercise and food stimuli in lean and obese women, studied by 24 h direct calorimetry. Br J Nutr. 1983;49:171–180. doi: 10.1079/bjn19830022. [DOI] [PubMed] [Google Scholar]
- 82.Celi FS, Brychta RJ, Linderman JD, Butler PW, Alberobello AT, Smith S, Courville AB, Lai EW, Costello R, Skarulis MC, Csako G, Remaley A, Pacak K, Chen KY. Minimal changes in environmental temperature result in a significant increase in energy expenditure and changes in the hormonal homeostasis in healthy adults. Eur J Endocrinol. 2010;163:863–872. doi: 10.1530/EJE-10-0627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Steiniger J, Karst H, Noack R, Steglich HD. Diet-induced thermogenesis in man: thermic effects of single protein and carbohydrate test meals in lean and obese subjects. Ann Nutr Metab. 1987;31:117–125. doi: 10.1159/000177258. [DOI] [PubMed] [Google Scholar]
- 84.Jensen AH, Kuhlman DE, Becker DE, Harmon BG. Response of growing-finishing swine to different housing environments during winter seasons. J Anim Sci. 1969;29:451–456. doi: 10.2527/jas1969.293451x. [DOI] [PubMed] [Google Scholar]
- 85.Young BA. Cold stress as it affects animal production. J Anim Sci. 1981;52:154–163. doi: 10.2527/jas1981.521154x. [DOI] [PubMed] [Google Scholar]
- 86.Berg F, Gustafson U, Andersson L. The uncoupling protein 1 gene (UCP1) is disrupted in the pig lineage: a genetic explanation for poor thermoregulation in piglets. PLoS Genet. 2006;2:e129. doi: 10.1371/journal.pgen.0020129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Mezentseva NV, Kumaratilake JS, Newman SA. The brown adipocyte differentiation pathway in birds: an evolutionary road not taken. BMC Biol. 2008;6:17. doi: 10.1186/1741-7007-6-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Overton JM. Phenotyping small animals as models for the human metabolic syndrome: thermoneutrality matters. Int J Obes (Lond) 2010;34 (Suppl 2):S53–S58. doi: 10.1038/ijo.2010.240. [DOI] [PubMed] [Google Scholar]