

Abstract
The Burkholderia cepacia Complex assembles at least eighteen closely related species that are ubiquitous in nature. Some isolates show beneficial potential for biocontrol, bioremediation and plant growth promotion. On the contrary, other strains are pathogens for plants and immunocompromised individuals, like cystic fibrosis patients. In these subjects, they can cause respiratory tract infections sometimes characterised by fatal outcome. Most of the Burkholderia cepacia Complex species are mucoid when grown on a mannitol rich medium and they also form biofilms, two related characteristics, since polysaccharides are important component of biofilm matrices. Moreover, polysaccharides contribute to bacterial survival in a hostile environment by inhibiting both neutrophils chemotaxis and antimicrobial peptides activity, and by scavenging reactive oxygen species. The ability of these microorganisms to produce exopolysaccharides with different structures is testified by numerous articles in the literature. However, little is known about the type of polysaccharides produced in biofilms and their relationship with those obtained in non-biofilm conditions. The aim of this study was to define the type of exopolysaccharides produced by nine species of the Burkholderia cepacia Complex. Two isolates were then selected to compare the polysaccharides produced on agar plates with those formed in biofilms developed on cellulose membranes. The investigation was conducted using NMR spectroscopy, high performance size exclusion chromatography, and gas chromatography coupled to mass spectrometry. The results showed that the Complex is capable of producing a variety of exopolysaccharides, most often in mixture, and that the most common exopolysaccharide is always cepacian. In addition, two novel polysaccharide structures were determined: one composed of mannose and rhamnose and another containing galactose and glucuronic acid. Comparison of exopolysaccharides obtained from cultures on agar plates with those extracted from biofilms on cellulose membranes showed important differences, thus suggesting that extrapolating data from non-biofilm conditions might not always be applicable.
Introduction
Bacterial exopolysaccharides are commonly biosynthesized by microbial species and can be grouped in two main types: capsular polysaccharides and extracellular polysaccharides (EPOLs) [1]. The formers are strongly bound to bacterial cell forming a capsule, while the latter are excreted in the medium around bacteria, often forming a slime. Several biological functions are attributed to EPOLs, but not all are fully known. Certainly, they contribute to create a suitable hydrophilic niche where bacteria may more comfortably survive, by providing accumulation of micronutrients. Furthermore, they have protective functions by forming a physical barrier around cells and exhibiting more or less specific interactions with environmental molecules potentially dangerous for the bacteria. In this context, also environmental conditions in infection sites have to be considered, where bacteria are threatened by components of the immune system, such as antimicrobial peptides [2] and reactive oxygen species [3], [4]. It can thus be reasonable to assume that bacteria use EPOLs as versatile tools for their survival. As a matter of fact, a single species is often able to synthesize a variety of EPOLs with distinct primary structures, and therefore diverse conformations, thus implying potential different biological properties.
Besides the above considerations, an interesting bacterial way of life, the biofilm, is deeply connected to EPOL functions. EPOLs are considered one of the main component of bacterial biofilms [5]–[7], but it has to be taken into account that biofilm composition is rather variable and depends on several parameters including culture media and the nature of the support to which cell adhesion occurs. In fact, in biofilms EPOLs are often considered as part of the tools used by bacteria for cell adhesion. Moreover, anti-biofilm and anti-adhesive properties of some EPOLs have recently been described [8], thus increasing the versatility of these macromolecules in terms of biological properties. It has to be stressed that, as pointed by Høiby et al. [9], only biofilms produced by Pseudomonas aeruginosa and Staphylococcus aureus have been deeply investigated, but a more general definition of matrix properties based on a wide number of bacterial species is still lacking. For the above reasons, microbiological investigations aimed at the description of the interactions of bacteria with their surroundings, in the environment as well as in infections, should imply an accurate knowledge of the structure of extracellular polysaccharides, as one of the main molecular characteristic of bacteria.
In the frame of an investigation on bacteria involved in lung infections of cystic fibrosis (CF) patients, and with the aim of developing a study devoted to disclose the EPOLs role in biofilm formation and maintenance, we thoroughly investigated the structure and the macromolecular properties of EPOLs produced by some members of the Burkholderia cepacia Complex (BCC) [1]. BCC includes several species of the genus Burkholderia which are currently found in many environmental niches where they also have beneficial effects for plant protection and growth [10], [11]. Contrary to this, some species of the genus are pathogens for plant, animals and vulnerable humans, specifically CF and Chronic Granulomatous Disease patients. The ability of some BCC species to cause serious lung infections in CF patients was first reported in 1984 and since then the number of BCC species found in CF lungs increased substantially. Contrary to other CF pathogens, some BCC infections lead to a rapid fatal decline known as “cepacia syndrome”. Until now the BCC was described to include eighteen species out of more than eighty characteristic of the genus, but the taxonomical definition of the BCC is still an open task [12]–[14]. We then decided to investigate 9 species (Table 1) which include environmental bacteria as well as those more frequently involved in lung infection of CF patients. Most of them are part of the suggested reference panel [12], [13] defined by members of the “International Burkholderia cepacia Working Group” (http://users.ugent.be/~tcoenye/index_bestanden/index_files/Page383.htm), while some others are type strains. Although these species are often the subject of studies implying properties related with EPOLs functions, there is a rather poor information on the structure of EPOLs produced by these 9 species; very often it is taken for granted that they produce the already described Cepacian (CEP) [1] as the main EPOL.
Table 1. BCC species investigated for the structure of their EPOLs.
| Species | LMG* | Other strain designation | |
| B. ambifaria | 19466 | AU0212 | Not ref. panel, not type strain |
| B. anthina | 20983 | C1765 | ref panel [13] |
| B. cenocepacia | 16659 | C1394 | ref. panel [12] |
| B. cenocepacia | 18829 | PC184 | ref. panel [12] |
| B. cepacia | 18821 | CEP 509 | ref. panel [12] |
| B. dolosa | 21820 | R12720 | type strain |
| B. lata | 22485 | b383 | type strain |
| B. multivorans | 16660 | C1576 | ref. panel[12] |
| B. stabilis | 18138 | Not ref. panel, not type strain | |
| B. vietnamiensis | 10929 | ref. panel [12] |
*BCCM/LMG = Belgian Co-ordinated Collections of Micro-organisms/Laboratory of Microbiology, Gent University, Belgium.
The investigation described in this paper was carried out resorting to NMR spectroscopy, gas-chromatography coupled to mass spectrometry (GC-MS) and high performance size exclusion chromatography (HP-SEC). It clarified the interesting versatility of this group of bacteria in terms of extracellular polysaccharides biosynthesis. In addition, preliminary data on matrix composition indicated that biofilm growth conditions can modulate EPOLs biosynthesis leading to production of a selected polymer or to quantitative changes of EPOLs molar ratio in case of mixtures, with respect to the non-biofilm mode of growth.
Materials and Methods
Bacterial strains and exopolysaccharides production and purification
Strains to be investigated were acquired from the Belgian Co-ordinated Collections of Micro-organisms at the Laboratory of Microbiology, Gent University, Belgium (Table 1). The strains B. cenocepacia BTS2, B. pyrrocinia BTS7 and B. cepacia BTS13, isolated from cystic fibrosis patients attending the Regional Centre for Cystic Fibrosis in Trieste, Italy [15], were used for the production of known EPOLs. Bacterial growth on Yeast Extract-Mannitol (YEM) agar plates, as a continuous layer, and EPOLs isolation and purification were performed as previously described [16]. Five independent EPOL extractions were performed with BTS2, twenty with BTS7 and six with BTS13 (these two strains are used as cepacian and galactan-Kdo producer, respectively), and two with all other strains.
Biofilm quantification in microtiter plates
B. cenocepacia BTS2 and B. multivorans C1576 were grown overnight in liquid Müller Hinton (MH) medium at 30°C with shaking and subsequently diluted to a concentration of 1×106 CFU/mL. A volume of 200 μL was used to inoculate the wells in a 96 wells microtiter plate. After incubating at 30°C for 48 h, bacterial growth was evaluated by reading the turbidity at 590 nm, while biofilm biomass was estimated at 570 nm after crystal violet staining [17]. The biofilm index, which is the ratio between the biomass and the cells growth, after subtraction of the proper blank, was used to quantify the biofilm.
Biofilm production on cellulose membranes
Biofilms were grown on cellulose membranes (Sigma, cut-off 12.400 Da) [17] which were prepared as follows: they were cut in circles to match the Petri dish, washed first in boiling 5% Na2CO3 and then in boiling water for 15 min, autoclaved and placed over Petri dishes, containing YEM or MH medium. Membranes covered the whole plate and excess of water was let to dry under the hood. An overnight liquid culture of bacteria (B. cenocepacia BTS2 or B. multivorans C1576) was diluted to 0.013 OD at 600 nm (about 1×106 CFU/mL) and 10 μL of the diluted suspension were placed on the membranes. The liquid medium used for the overnight culture was the same of the seeded Petri dish. After 7 days of incubation at 30°C, the material on the membranes was recovered in 5–10 mL of 0.9% NaCl, centrifuged at 4000 rpm at 4°C for 20 min, and finally filtered sterilised (Millipore membranes 0.22 μm). When YEM medium was used, separation of the cells was achieved by centrifuging at 20000 rpm at 4°C for 30 min, due to the high viscosity of the EPOL solution. Removal of proteins was achieved by treatment with protease (from Streptomyces griseus, Sigma) in TRIS-HCl 50 mM, pH 7.5. The EPOLs were purified by dialysis, first against 0.1 M NaCl and then water, the solutions were then taken to neutral pH and filtered. The extractions were repeated three times for both species.
General procedures for structural analysis
Hydrolysis of the polysaccharides was conducted with 2 M TFA for 1 h at 125°C and alditol acetates were prepared as already described [18]. Permethylation of the EPOL was achieved following the protocol by Harris [19], while carboxyl reduction was performed with NaBD4 as previously described [20]. Analytical gas-chromatography (GC) was performed on a Perkin–Elmer Autosystem XL gas chromatograph equipped with a flame ionisation detector and an SP2330 capillary column (Supelco, 30 m), using He as carrier gas. The following temperature programmes were used: for alditol acetates, 200–245°C at 4°C/min; for partially methylated alditol acetates, 150–250°C at 4°C/min. GC–MS analyses were carried out on an Agilent Technologies 7890A gas chromatograph coupled to an Agilent Technologies 5975C VL MSD. In the case of overlapping peaks of partially methylated alditol acetates, GC-MS was performed also on a HP-1 column (Agilent Technologies, 30 m) using the temperature program 120–245°C at 2°C/min.
NMR spectroscopy
The molecular mass of EPOLs was decreased by treating their solutions (1 g/L) with a Branson sonifier equipped with a microtip at 2.8 Å. Samples were cooled in an ice bath and sonicated using 5 bursts of 1 min each, separated by 1 min intervals. They were subsequently exchanged three times with 99.9% D2O by lyophilisation and finally dissolved in 0.7 mL 99.96% D2O. Spectra were recorded on a 500 MHz VARIAN spectrometer operating at 50°C. Chemical shifts are expressed in ppm using acetone as external reference (2.223 ppm for 1H). NMR spectra were analysed using Mestrenova software.
Molecular mass determination
High performance size exclusion chromatography (HP-SEC) was performed on an Agilent Technologies 1200 series HPLC equipped with three columns in series (Tosoh Bioscience, TSKgel G3000PW, G5000PW and G6000PW, i.d. 7.5 mm, length 30 cm) kept at 40°C with a thermostat (Waters Millipore). Calibration of the chromatographic system was performed using pullulan standards (Polymer Laboratories, Germany and Sigma for pullulan with MM = 1.6×106). Elution was performed with 0.15 M NaCl, with a flow rate of 0.4 mL/min and monitored using a refractive index detector (Knauer, Labservice Analytica), interfaced with a computer via Agilent software.
Results and Discussion
The knowledge already acquired on the structure of the EPOLs produced by some BCC species, and in particular on the structural fingerprints given by 1D 1H-NMR spectra, prompted the use of this technique to define the EPOLs produced by the selected species for the present investigation. Besides NMR, classical monosaccharide composition and glycosidic linkage analyses were also used to gain structural information on novel EPOLs. In addition, since the biological activity depends also on the dimension of the macromolecules, EPOLs molecular masses were conveniently investigated by means of HP-SEC. The data acquired were compared with those already available in our laboratory and in the literature.
In order to facilitate the following discussion on polysaccharides structure, the NMR data of the EPOLs produced by three BCC clinical isolates investigated in our laboratory, B. cenocepacia BTS2, B. pyrrocinia BTS7, and B. cepacia BTS13 [21] are hereafter summarised, since they were particularly useful for the present study. Moreover, NMR data for BTS2 and BTS7 polysaccharides were never published before.
EPOLs produced by B. pyrrocinia BTS7
B. pyrrocinia BTS7 produces the EPOL named cepacian (CEP), which is constituted by a complex repeating unit and contains glucose, mannose, galactose, the unusual D-rhamnose together with a glucuronic acid fully substituted on its hydroxyl groups (Figure 1). Prior to NMR, the sample was de-acetylated and sonicated, to enhance the resolution of the spectrum. In the 1H NMR spectrum (Figure 2A), anomeric proton assignments are reported in agreement with previous articles [22]–[24]. In the same ppm range, the small signals indicated with asterisks at 5.51, 5.41 and 4.99 ppm could not be attributed neither to CEP, nor to any other known EPOL of the BCC, thus suggesting the presence of a novel EPOL hereafter referred to as CO-CEP. Different polysaccharide productions always contained both EPOLs, with CO-CEP in very small amounts (less than 0.10 CO-CEP/CEP repeating units molar ratio). Separation of the two EPOLs by gel filtration chromatography was attempted (data not shown), but it was unsuccessful, probably due to the viscous nature of CEP solution and to its aggregation ability. The small amounts of CO-CEP produced by strain BTS7 always in mixture with CEP did not permit the determination of its structure.
Figure 1. Repeating units structure of exopolysaccharides produced by BCC species.

Primary structures of the repeating units of the exopolysaccharides CEP, GAL-KDO, BV-EP and CO-CEP.
Figure 2. 1H-NMR spectra of exopolysaccharides produced by BCC species.

A) 1H-NMR spectrum of de-acetylated CEP produced by B. pyrrocinia BTS7. The anomeric proton region is enlarged in the inset. Anomeric proton resonances of CEP are indicated, asterisks refer to signals of CO-CEP. B) 1H NMR spectrum of GAL-KDO produced by B. cepacia BTS13 grown on YEM agar plates: anomeric proton resonances together with H-2 of acetylated β-Gal and H-3 of Kdo are indicated. C) 1H NMR spectrum of the CEP/GAL-KDO mixture produced by B. cenocepacia BTS2 grown on YEM agar plates. The anomeric proton region is enlarged in the inset; signals belonging to CEP are indicated with the suffix C. Methyl groups of O-acetyl esters (-CH3 OAc) and rhamnose residues (-CH3 α-Rha) are shown.
EPOLs produced by B. cepacia BTS13
B. cepacia BTS13 produced two different EPOLs, depending on the medium used [21]: a linear polysaccharide constituted of three galactose residues, one acetyl substituent and one 3-deoxy-D-manno-oct-2-ulosonic acid (Kdo), named galactan-Kdo (GAL-KDO), (Figure 1, GAL-KDO) when grown on King A, and a mixture of the former with a linear 2,6-linked β-fructan, known as levan, in YEM medium. The 1H-NMR spectrum of GAL-KDO is shown in Figure 2B where anomeric proton resonances together with signals due to acetylation are indicated. Moreover, the two H-3 resonances of the Kdo residue are found at about 2.50 and 1.80 ppm, well separated from the other signals, thus being diagnostics for the presence of such residue.
EPOLs produced by B. cenocepacia BTS2
This clinical isolate produced a mixture of GAL-KDO and CEP when grown on YEM agar plates, as established by NMR spectroscopy analysis (Figure 2C). Identification of the NMR resonances was achieved by comparison with the NMR spectra of the individual polysaccharides described in the previous paragraphs. The anomeric region of the 1H-NMR spectrum (Figure 2c) was rather crowded, but the resonances of CEP were clearly visible. On the contrary, the anomeric protons of GAL-KDO were not well resolved, since they partially overlapped with H-1 of 3-linked Glc of CEP and with other resonances due to acetylation. However, the presence of GAL-KDO was undoubtedly established by the H-3′s signals at 2.51 and 1.83 ppm. Integration of the peak areas belonging to H-3′s of Kdo and that one of C-6 of Rha (1.24 ppm) of CEP repeating unit established a ratio of about 0.18 for the two EPOLs.
Composition analysis of the EPOLs produced by the investigated strains
The overall composition homogeneity of the EPOLs produced by the 10 investigated isolates, belonging to the 9 selected species, was determined by GC analysis of their alditol acetates derivatives and the results were compared with data available in the literature for CEP (Table 2) [23], the most common exopolysaccharide of the BCC. Out of the ten products examined, only two EPOLs had a substantially different composition: the one from B. vietnamiensis, containing fucose residues, and the one from B. multivorans C1576, characterised by a relative molar ratios of saccharides quite different from CEP. The composition of all other polysaccharides did not show striking differences and indicated mainly the production of CEP.
Table 2. Determination of neutral monosaccharides in the EPOLs of different BCC bacterial isolates and comparison with literature data for CEP [23].
| monosaccharidea | |||||
| Species | Rha | Fuc | Man | Gal | Glc |
| B. ambifaria | 0.99 | 0.16 | 2.69 | 1.00 | |
| B. anthina | 0.92 | 0.64 | 2.43 | 1.00 | |
| B. cenocepacia 16659 | 0.74 | 0.52 | 2.48 | 1.00 | |
| B. cenocepacia 18829 | 0.77 | 0.79 | 3.06 | 1.00 | |
| B. cepacia | 0.91 | 0.48 | 3.12 | 1.00 | |
| B. dolosa | 0.80 | 0.34 | 2.05 | 1.00 | |
| B. lata | 0.92 | 0.42 | 2.44 | 1.00 | |
| B. multivorans | 2.22 | 1.64 | 2.60 | 1.00 | |
| B. stabilis | 0.62 | 0.34 | 2.07 | 1.00 | |
| B. vietnamiensis | 0.31 | 0.23 | 0.08 | 1.00 | 1.00 |
| Cepacian [23] | 0.71 | 0.50 | 2.79 | 1.00 | |
Monosaccharide content is expressed as molar ratio relative to glucose.
1H-NMR spectroscopy and linkage determination of the EPOLs produced by the investigated strains
The EPOLs produced by all strains were investigated by 1D 1H-NMR spectroscopy in their native as well as de-acetylated forms. The spectra showed that they all produced CEP, confirming that its biosynthetic gene cluster is widely distributed within the BCC. NMR spectroscopy revealed also that many isolates were able to biosynthesise polysaccharides different from CEP. In fact, the production of CEP was almost invariably partnered by CO-CEP, except in the case of B. multivorans C1576. B. anthina, B. ambifaria, and the two strains of B. cenocepacia C1394 and 18829 exhibited the highest CO-CEP co-synthesizing ability. The ratio between CO-CEP and CEP in these species was in the range 0.1-0.3, as measured by integrating the anomeric proton signals at 4.76 ppm, belonging to 3-linked Glc in CEP, and the one at 4.98 ppm, attributed to CO-CEP. Due to the impossibility of isolating this polysaccharide, its structural investigation was carried out on the EPOLs mixture produced by B. ambifaria, characterised by a rather high amount of CO-CEP. 2D-NMR spectroscopy investigation (data not shown) was not decisive, because of signals overlapping. Useful information was obtained with methylation analysis, performed on the native as well as the carboxyl reduced EPOLs mixture. The results (Table 3) revealed the presence of two novel residues, besides the expected ones composing CEP: 1,6-linked-Gal and 1,2,4-linked-Hex. Due to reduction with NaBD4, the latter derivative was di-deuterated on C-6, thus identifying it with an hexuronic acid in the native polysaccharide. The lack of 1,2,4-linked standard hexoses to determine the identity of the branched hexuronic acid was filled by comparing the retention time of several branched hexoses available in the lab with published data [25]. The results suggested to identify it with glucose establishing that native CO-CEP contains 1,2,4-linked GlcA. Moreover, the lack of a terminal non-reducing residue different from t-Gal, suggested that t-Gal itself could be the substitution on the branched glucuronic acid. These three residues represent the composition of CO-CEP and are in agreement with the presence of three NMR signals at 5.51, 5.41 and 4.99 ppm (signals with asterisks in Figure 2). A tentative structure is then constituted of a disaccharidic backbone 6)-α-D-Galp-(1→4/2)-α-D-GlcAp-(1→ with a terminal α-D-Galp substituting carbon 2 or 4 of the glucuronic acid residue. The position of the side chain substitution could not be assigned.
Table 3. Determination of glycosidic linkages in the EPOLs of B. ambifaria before (Ba) and after carboxyl reduction (Ba-R), and B. multivorans C1576 separated on SP2330 (Bm-I) and on HP1 (Bm-II) columns.
| Linked residuea | RRTb | Ba | Ba-R | Bm-I | Bm-II | RRTc |
| 2-Rha | 0.91 | 0.72 | 0.64 | 1.00 | 1.00 | 0.84 |
| 3-Rha | 0.93 | 0.43 | 0.43 | 0.86 | ||
| t-Gal | 1.00 | 2.27 | 2.11 | 1.71 | 1.82 | 1.00 |
| 3-Glc | 1.13 | 1.00 | 1.00 | 0.68 | ||
| 2-Man+3-Man | 1.14 | 0.92 | ||||
| 3-Glc+2-Man | 1.31 | 1.17 | ||||
| 3-Man | 0.47 | 1.20 | ||||
| 3-Gal | 1.17 | 0.20 | 0.21 | 0.13 | 0.17 | 1.22 |
| 6-Gal | 1.30 | 0.20 | 0.19 | |||
| 2,4-GlcD2 d | 1.40 | 0.09 | ||||
| 3,6-Man | 1.43 | 0.39 | 0.50 | 0.30 | 0.29 | 1.51 |
| 2,3,4-GlcD2 d | 1.47 | - | 0.21 | - | - |
Position of glycosidic linkages.
Relative retention time on SP2330 column.
Relative retention time on HP1 column.
C6 di-deuterated according to GC–MS.
B. cepacia, B. ambifaria, B. dolosa, and B. stabilis were able to co-synthesize also the GAL-KDO exopolysaccharide, as revealed by the presence of the diagnostic resonances of Kdo H-3′s at about 2.50 and 1.80 ppm (data not shown). B. cepacia showed the highest GAL-KDO/CEP repeating units molar ratio (0.3), followed by B. stabilis (0.03), and by the other isolates which had a content below 0.03.
A remarkable case of co-synthesis is B. vietnamiensis which produced two distinct EPOLs: CEP and the one named BV-EP (Figure 1) whose structural definition was reported in a separate article [26]. The 1H-NMR spectrum of the mixture CEP and BV-EP is reported in Figure 3A. BV-EP is totally different from CEP and it is the only EPOL of the BCC to contain fucose residues, a feature generally not very common in bacterial polysaccharides. Out of three independent growths and extractions, CEP and BV-EP were co-produced twice in the same molar ratio, as revealed by 1H-NMR spectra (data not shown), while a third batch contained only the polysaccharide BV-EP. This is the only case observed in our studies of such a drastic change in EPOL biosynthesis; usually the types of polysaccharides produced are constant and, in the case of mixtures, the ratio of the components may vary only slightly.
Figure 3. 1H-NMR spectra of the exopolysaccharides produced by B. vietnamiensis and B. multivorans.

A) 1H-NMR spectrum of the EPOLs mixture produced by B. vietnamiensis on YEM agar plates. Anomeric proton signals and methyl groups of 6-deoxy sugars belonging to CEP and BV-EP are shown. B) 1H-NMR spectrum of the EPOLs mixture produced by B. multivorans C1576 on YEM agar plates: anomeric proton signals and methyl groups of 6-deoxy sugar of CEP are indicated, while triangles indicate new signals.
Considering B. multivorans C1576, the EPOLs proton anomeric region (Figure 3B) showed resonances belonging to CEP together with new signals at 5.27, 5.24 and 5.04 ppm, indicated with triangles. In addition, two distinct –CH3 resonances attributed to H-6′s of rhamnose were found at 1.30 and 1.24 ppm, indicating the presence of at least another Rha, besides that one in CEP. The ratio of the area of the two peaks was 1.2, indicating roughly equimolar amounts of rhamnose in the two polymers. These findings were in agreement with the composition analysis (Table 2), where rhamnose and mannose residues were more abundant than in CEP alone [23], thus suggesting the biosynthesis of a novel EPOL. In order to gain structural information, the mixture of B. multivorans C1576 EPOLs was subjected to methylation analysis and the results are reported in Table 3. Besides the typical residues attributable to CEP, 3-linked rhamnose, 2-linked mannose and 3-linked mannose were also present. In addition, the 2-linked rhamnose was present in higher amount than found in CEP, indicating that it could also be part of the novel EPOL. Quantitative data were impeded by co-elution of 2-linked Man and 3-linked Man when GC was performed on the SP-2330 column, as revealed by the e.i. mass spectrum. However, using the HP-1 column, 3-linked Man was well separated, while co-elution resulted for 2-linked Man and 3-linked Glc (Table 3). Since the integration for all well separated derivatives on both columns was very similar, the amount of 2-linked Man was obtained with a simple subtraction to give: 2-Man = 0.49 and 3-Man = 0.43, relative to 2-Rha. These data are in good agreement with the NMR results and indicate that B. multivorans C1576 produces, besides CEP, a novel rhamnomannan (RHA-MAN) constituted of 2-linked Rha, 3-linked Rha, 2-linked Man and 3-linked Man in equimolar amounts. More structural information on this EPOL was obtained during the preliminary investigation of the carbohydrate fractions in biofilms, as described in the last paragraph of the "Results and Discussion" section.
Macromolecular characterization
Besides primary structure, the overall conformation of a polymer depends on its dimensions. The EPOL molecular mass has a strong impact on the biological properties, particularly those related to the structure of the matrix in biofilms, a cellular scaffold filled with water and other useful macro- and micro-molecules. Therefore, the weight average molecular masses of the EPOLs produced by BCC strains were determined by HP-SEC analysis and the values are reported in Table 4 together with the polydispersity index (D) values. This index is the ratio between the weight- and the number-average molecular mass and indicates the degree of molecular masses dispersion for a given polymeric species: the higher the figure the higher the dispersion. It is interesting to note that only the EPOLs mixture produced by B. multivorans C1576 and B. vietnamiensis exhibited a bimodal elution profile, as illustrated in Figure 4. For the EPOLs of the former species, peak assignment was based on the relative amounts of the two polysaccharide, with RHA-MAN being less abundant than CEP (about 0.5 repeating units molar ratio), as indicated by integration of the C-6′s of rhamnose residues in the 1H-NMR spectrum. Therefore, the most intense peak, characterised by a higher MM was attributed to CEP, while the less intense one, having a lower MM, was assigned to RHA-MAN (Figure 4A). In the case of B. vietnamiensis (Figure 4B), CEP was characterised by a lower molecular mass than BV-EP, as established by composition analysis of the two well separated fractions [26]. Contrary to this, the presence of the CO-CEP polymer was not accompanied by extra peaks in the HP-SEC experiments, the only exception was the HP-SEC trace obtained for the EPOLs of B. cenocepacia 18829, where a shoulder on the right side of the main polymer elution (Figure 4c) might be attributed to CO-CEP, based on the relative abundance of the two EPOLs. The co-production of GAL-KDO did not result in a bimodal distribution or in the appearance of a shoulder in the HP-SEC chromatograms.
Table 4. Weight average molecular masses (MM, Da) and polydispersity index (D) of the EPOL fractions produced by the investigated BCC species.
| Peak I | Peak II | |||
| Species | MM (Da) | D | MM (Da) | D |
| B. ambifaria LMG19466 | 2.56×105 | 1.84 | ||
| B. anthina LMG20983 | 2.34×106 | 3.42 | ||
| B. cenocepacia LMG16659 | 8.49×106 | 4.38 | ||
| B. cenocepacia LMG18829 | 8.99×106 | 6.68 | ||
| B. cepacia LMG18821 | 1.80×107 | 4.94 | ||
| B. dolosa LMG21820 | 9.63×106 | 5.79 | ||
| B. lata LMG22485 | 1.19×105 | 2.40 | ||
| B. multivorans LMG16660 | 2.09×107 | 2.48 | 4.98×105 | 1.05 |
| B. stabilis LMG18138 | 7.57×106 | 4.18 | ||
| B. vietnamiensis LMG10929 | 1.43×107 | 3.61 | 2.06×104 | 1.93 |
Figure 4. HP-SEC chromatograms of exopolysaccharides mixtures.
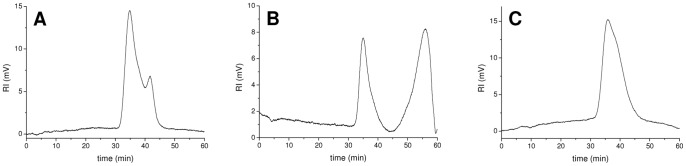
EPOL mixtures produced by B. multivorans C1576 (A), B. vietnamiensis (B) and B. cenocepacia 18829 (C).
Concerning numerical values of the molecular masses, all the polymeric fractions investigated had rather high figures. These data might reflect the aggregation capacity exhibited by CEP which forms complexes with low stoichiometry (two- or four-stranded complexes) as already reported in previous articles [27], [28]. Therefore, the real values of molecular masses for single CEP polymeric chains might be lower than those detected.
EPOLs produced in biofilms
In a preliminary study of biofilm matrix composition, the structure of EPOLs produced by two species of the BCC was investigated. The focus was centred on B. multivorans C1576 and B. cenocepacia BTS2, both producing mixtures of EPOLs on YEM agar plates. The latter strain was selected because it is an abundant biofilm producer, as indicated by crystal violet staining when cultured in microtiter plates (data not shown).
Biofilms were grown for seven days on cellulose porous membranes placed on top of agar plates, containing the appropriate culture medium, either YEM or MH. As previously described, in non-biofilm conditions (i.e. classical agar plates), on YEM medium C1576 produced a mixture of CEP and a novel EPOL named RHA-MAN (Figure 3, Table 3). Surprisingly, 1H-NMR spectra showed that in biofilm-forming conditions, YEM medium stimulated only the biosynthesis of CEP, whilst MH medium induced only production of the RHA-MAN as shown in the proton NMR spectrum (Figure 5A). In fact, the anomeric region showed three resonances at 5.27, 5.24 and 5.04 ppm, having integration values of 1.0: 1.0: 2.0. At 1.30 ppm a signal attributable to a methyl group of 6-deoxyhexose gave an integration value of 6∶0, thus suggesting the presence of two rhamnose residues. Composition analysis indicated that it contained rhamnose and mannose in equimolar amounts, thus undoubtedly identifying it with the RHA-MAN EPOL, co-produced with CEP when bacteria were grown on YEM medium agar plates in non-biofilm conditions.
Figure 5. 1H-NMR spectra of exopolysaccharides from biofilms.

A) anomeric region of the 1H-NMR spectra of the EPOLs obtained from biofilms of B. multivorans C1576 produced on cellulose membranes deposited on MH (top) and YEM (bottom) agar plates; B) anomeric region of the 1H NMR spectra of EPOLs produced by B. cenocepacia BTS2 in biofilm conditions in MH (top) and YEM (bottom) media. CEP, GAL-KDO and RHA-MAN indicate signals belonging to the respective polysaccharides. Circles indicate new resonances of unknown identity.
Therefore, the biofilm culture conditions drove C1576 to produce specific polysaccharides as a function of the medium employed. This observation is very interesting since it brings to the conclusion that EPOLs produced in biofilms developed on cellulose membranes might be very different from those biosynthesized during growth on solid agar medium. This conclusion was also achieved by investigating the clinical isolate B. cenocepacia BTS2. This isolate produced a mixture of CEP and GAL-KDO (Figure 2C) when grown directly on agar. However, in biofilm-forming conditions, the mixture was produced only in YEM medium where an increase in the amount of GAL-KDO was observed (0.60 GAL-KDO/CEP repeating units), while MH stimulated the production of GAL-KDO with only scanty amount of CEP (Figure 5B). At the same time, two new resonances with considerable intensities were detected in the 1H NMR spectrum at 5.14 and 4.62 ppm (indicated with circles), thus showing the production of a further novel EPOL.
Conclusions
More than two decades ago, Sage and co-workers [29] described the slime produced by CF isolates of Burkholderia cepacia, at that time still named Pseudomonas cepacia, as being composed of galactose, glucose, mannose, glucuronic acid, and rhamnose, when grown on excess of glucose or mannitol as carbon source, thus suggesting the presence of CEP. Since then, the research in this field made great progress: the BCC was established as a group composed of at least eighteen closely related species [12]–[14] and detailed structural studies on the biosynthesised EPOLs appeared in the literature. CEP resulted to be the main exopolysaccharide of the BCC, especially when bacteria were grown on YEM medium and always in non-biofilm conditions [16], [30]. Besides CEP, other EPOLs were described: PS-I was first found in France [31] and afterwards in our laboratory [30]; dextran was produced together with CEP and PS-I by a B. cenocepacia clinical isolate investigated in Canada [31], and levan, a fructose polymer, was detected in our laboratory, co-synthetized with GAL-KDO by B. cepacia BTS13 [21]. Very recently, B. vietnamiensis EPOL was determined to have a hexasaccharidic repeating unit containing fucose, glucose and glucuronic acid [26]. The results reported in this paper further contributed to unravel the complex picture of the polysaccharide biosynthetic potential of the BCC, but not all the details were clarified so that additional studies should be carried out. In the present study, CEP was confirmed to be the EPOL produced on YEM medium by the majority of the species of the BCC in non-biofilm conditions. Moreover, eight species investigated co-synthesized another extracellular polysaccharide (CO-CEP) containing t-Gal, 6-Gal and 1,2,4-GlcA. B. multivorans C1576 co-synthesized a novel linear EPOL containing 2-Rha, 3-Rha, 2-Man and 3-Man in equimolar amounts (Table 5).
Table 5. Summary of the different EPOLs produced by each bacterial species when grown on YEM solid medium.
| Species | LMG | CEP | GAL-KDO | CO-CEP | BV-EP | RHA-MAN |
| B. ambifaria | 19466 | + | + | + | ||
| B. anthina | 20983 | + | + | |||
| B. cenocepacia | 16659 | + | + | |||
| B. cenocepacia | 18829 | + | + | |||
| B. cepacia | 18821 | + | + | + | ||
| B. dolosa | 21820 | + | + | + | ||
| B. lata | 22485 | + | + | |||
| B. multivorans | 16660 | + | + | |||
| B. stabilis | 18138 | + | + | + | ||
| B. vietnamiensis | 10929 | + | + |
Some chemical features are common to the majority of the EPOLs investigated. Specifically, the molecular mass is usually larger than 106 Daltons and the polymer chains often bear negative charges due to the presence of carboxylate groups, mostly uronic acids, thus foreseeing a certain degree of chain rigidity due to charge repulsion.
Considering the species investigated in biofilm conditions, it must be underlined that the biosynthesis of CEP was stimulated by YEM medium both in non-biofilm and in biofilm mode of growth. However, when biofilm was formed on cellulose membranes deposited on MH agar plates, polysaccharides other than CEP were prevalent. This is an important information underlying that the type of EPOLs synthesised not only varies with the medium used, but it is also influenced by the presence of the solid support.
In conclusion, the species of the BCC are characterised by an extremely high variability in the EPOLs production. Taking into account that the presence of the extracellular polysaccharidic matrix might be strongly influenced by the bacteria environmental conditions, the EPOL identity should be carefully considered for microbiological investigations including biofilm formation ability, where EPOL might have a specific role in controlling the hydrophilic matrix surrounding bacterial cells and modulating its properties in response to external conditions.
Funding Statement
This work was supported by the Italian Ministry of University and Research (http://www.istruzione.it/): PRIN 2009; Friuli Venezia Giulia region (http://www.regione.fvg.it/rafvg/cms/RAFVG/): project R3A2 within L.R.26/2005; and the University of Trieste (http://www.units.it/): FRA 2011. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Cescutti P (2009) Bacterial capsular polysaccharides and exopolysaccharides. In Moran AP, Brennan PJ, Holst O, von Itzstein M, editors. Microbial Glycobiology: Structures, Relevance and Application. New York: Academic Press. pp. 93–108.
- 2. Foschiatti M, Cescutti P, Tossi A, Rizzo R (2009) Inhibition of cathelicidin activity by bacterial exopolysaccharides. Mol Microbiol 72(5): 1137–1146. [DOI] [PubMed] [Google Scholar]
- 3. Cuzzi B, Cescutti P, Furlanis L, Lagatolla C, Sturiale L, et al. (2012) Investigation on bacteria resistance to the immune system response. Cepacian depolymerisation by reactive oxygen species. Inn Immun 18(4): 661–671. [DOI] [PubMed] [Google Scholar]
- 4. Bylund J, Burgess LA, Cescutti P, Ernst RK, Speert DP (2006) Exopolysaccharides from Burkholderia cenocepacia inhibit neutrophil chemotaxis and scavenge reactive oxygen species. J Biol Chem 281: 2526–2532. [DOI] [PubMed] [Google Scholar]
- 5. Flemming HC, Wingender J (2010) The biofilm matrix. Nat Rev Microbiol 8: 623–633. [DOI] [PubMed] [Google Scholar]
- 6. Mann EE, Wozniak DJ (2012) Pseudomonas biofilm matrix composition and niche biology. FEMS Microbiol Rev 36: 893–916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Bales PM, Miljkovic Renke E, May SL, Shen Y, Nelson DC (2013) Purification and characterization of biofilm-associated EPS exopolysaccharides from ESKAPE organisms and other pathogens. Plos One 8(6): 1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Rendueles O, Kaplan JB, Ghigo JM (2012) Antibiofilm polysaccharides. Environ Microbiol 15(2): 334–346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Bjarnsholt T, Høiby N, Donelli G, Imbert C, Försberg Å (2012) Understanding biofilms-are we there yet? FEMS Immunol Med Microbiol 65: 125–126 DOI:10.1111/j.1574-695X.2012.00984.x [DOI] [PubMed] [Google Scholar]
- 10. Mahenthiralingam E, Urban TA, Goldberg JB (2005) The multifarious, multireplicon Burkholderia cepacia complex. Nat Rev Microbiol 3: 144–156. [DOI] [PubMed] [Google Scholar]
- 11. Payne GW, Ramette A, Rose HL, Weightman AJ, Jones TH, et al. (2006) Application of a recA gene-based identification approach to the maize rhizosphere reveals novel diversity in Burkholderia species. FEMS Microbiol Lett 259: 126–132. [DOI] [PubMed] [Google Scholar]
- 12. Mahenthiralingam E, Coenye T, Chung JW, Speert DP, Govan JRW, et al. (2000) Diagnostically and experimentally useful panel of strains from the Burkholderia cepacia Complex. J Clin Microbiol 38(2): 910–913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Coenye T, Vandamme P, LiPuma JJ, Govan JRW, Mahenthiralingam E (2003) Updated version of the Burkholderia cepacia Complex experimental strain panel. J Clin Microbiol 41(6): 2797–2798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Peeters C, Zlosnik JE, Spilker T, Hird TJ, LiPuma JJ, et al. (2013) Burkholderia pseudomultivorans sp. nov., a novel Burkholderia cepacia complex species from human respiratory samples and the rhizosphere. Syst Appl Microbiol 36(7): 483–489 doi:10.1016/j.syapm.2013.06.003 [DOI] [PubMed] [Google Scholar]
- 15. Lagatolla C, Skerlavaj S, Dolzani L, Tonin EA, Monti Bragadin C, et al. (2002) Microbiological characterisation of Burkholderia cepacia isolates from cystic fibrosis patients. Investigation of the exopolysaccharides produced. FEMS Microbiol Lett 209: 89–94. [DOI] [PubMed] [Google Scholar]
- 16. Chiarini L, Cescutti P, Drigo L, Impallomeni G, Herasimenka Y, et al. (2004) Exopolysaccharides produced by Burkholderia cenocepacia recA lineages IIIA and IIIB. J Cyst Fibros 3: 165–172. [DOI] [PubMed] [Google Scholar]
- 17.Merritt JH, Kadouri DE, O'Toole GA (2011) Growing and Analyzing Static Biofilms. Curr Protoc Microbiol 1B.1.1–1B.1.18. [DOI] [PMC free article] [PubMed]
- 18. Albersheim P, Nevins DJ, English PD, Karr A (1967) A method for the analysis of sugars in plant cell-wall polysaccharides by gas-liquid chromatography. Carbohydr Res 5: 340–345. [Google Scholar]
- 19. Harris PJ, Henry RJ, Blakeney AB, Stone BA (1984) An improved procedure for the methylation analysis of oligosaccharides and polysaccharides. Carbohydr Res 127: 59–73. [DOI] [PubMed] [Google Scholar]
- 20. Osman SF, Fett WF, Fishman ML (1986) Exopolysaccharides of the phytopathogen Pseudomonas syringae pv. glycinea . J Bacteriol 166(1): 66–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Cescutti P, Impallomeni G, Garozzo D, Sturiale L, Herasimenka Y, et al. (2003) Exopolysaccharides produced by a clinical strain of Burkholderia cepacia isolated from a cystic fibrosis patient. Carbohydr Res 338: 2687–2695. [DOI] [PubMed] [Google Scholar]
- 22. Cérantola S, Lemassu-Jacquier A, Montrozier H (1999) Structural elucidation of a novel exopolysaccharide produced by a mucoid clinical isolate of Burkholderia cepacia. Characterization of a trisubstituted glucuronic acid residue in a heptasaccharide repeating unit. Eur J Biochem 260: 373–83. [DOI] [PubMed] [Google Scholar]
- 23. Cescutti P, Bosco M, Picotti F, Impallomeni G, Leitão JH, et al. (2000) Structural study of the exopolysaccharide produced by a clinical isolate of Burkholderia cepacia . Biochem Biophys Res Comm 273: 1088–1094. [DOI] [PubMed] [Google Scholar]
- 24. Linker A, Evans LR, Impallomeni G (2001) The structure of a polysaccharide from infectious strains of Burkholderia cepacia . Carbohydr Res 335: 45–54. [DOI] [PubMed] [Google Scholar]
- 25.Carpita NC, Shea EM (1989) Linkage structure of Carbohydrates by gas chromatography-mass spectrometry (GC-MS) of partially methylated alditol acetates. In: Biermann CJ, McGinnis GD, editors. Analysis of carbohydrates by GLC and MS. Boca Raton, Florida: CRC Press, Inc. pp. 157–216. [Google Scholar]
- 26. Cescutti P, Cuzzi B, Herasimenka Y, Rizzo R (2013) Structure of a novel exopolysaccharide produced by Burkholderia vietnamiensis, a cystic fibrosis opportunistic pathogen. Carbohydr Pol 94: 253–260. [DOI] [PubMed] [Google Scholar]
- 27. Sist P, Cescutti P, Skerlavaj S, Urbani R, Leitão JH, et al. (2003) Macromolecular and solution properties of Cepacian: the exopolysaccharide produced by a strain of Burkholderia cepacia isolated from a cystic fibrosis patient. Carbohydr Res 338: 1861–1867. [DOI] [PubMed] [Google Scholar]
- 28. Herasimenka Y, Cescutti P, Sampaio Noguera CE, Ruggiero JR, Urbani R, et al. (2008) Macromolecular properties of cepacian in water and dimethylsuphoxide. Carbohydr Res 343: 81–89. [DOI] [PubMed] [Google Scholar]
- 29. Sage A, Linker A, Evans LR, Lessie TG (1990) Hexose phosphate metabolism and exopolysaccharide formation in Pseudomonas cepacia . Curr Microbiol 20(3): 191–198. [Google Scholar]
- 30. Herasimenka Y, Cescutti P, Impallomeni G, Campana S, Taccetti G, et al. (2007) Exopolysaccharides produced by clinical strains belonging to the Burkholderia cepacia complex. J Cyst Fibros 6: 145–152. [DOI] [PubMed] [Google Scholar]
- 31. Cérantola S, Marty N, Montrozier H (1996) Structural studies of the acidic exopolysaccharide produced by a mucoid strain of Burkholderia cepacia isolated from cystic fibrosis. Carbohydr Res 285: 59–67. [DOI] [PubMed] [Google Scholar]
- 32. Conway BA, Chu KK, Bylund J, Altman E, Speert DP (2004) Production of exopolysaccharide by Burkholderia cenocepacia results in altered cell-surface interactions and altered bacterial clearance in mice. J Infect Dis 190(5): 957–966. [DOI] [PubMed] [Google Scholar]