

Abstract
Background
Smokers are more prone to oral infections than are non-smokers. Cigarette smoke reaches the host cells but also microorganisms present in the oral cavity. The contact between cigarette smoke and oral bacteria promotes such oral diseases as periodontitis. Cigarette smoke can also modulate C. albicans activities that promote oral candidiasis. The goal of this study was to investigate the effect of cigarette smoke condensate on C. albicans adhesion, growth, and biofilm formation as well as the activation of EAP1, HWP1 and secreted aspartic protease 2.
Results
Cigarette smoke condensate (CSC) increased C. albicans adhesion and growth, as well as biofilm formation. These features may be supported by the activation of certain important genes. Using quantitative RT-PCR, we demonstrated that CSC-exposed C. albicans expressed high levels of EAP1, HWP1 and SAP2 mRNA and that this gene expression increased with increasing concentrations of CSC.
Conclusion
CSC induction of C. albicans adhesion, growth, and biofilm formation may explain the increased persistence of this pathogen in smokers. These findings may also be relevant to other biofilm-induced oral diseases.
Keywords: Cigarette smoke, Tobacco, C albicans, Adhesion, Growth, Biofilm, Genes, EAP-1, HWP-1, Sap2
Background
One of the most commonly encountered opportunistic microorganisms in humans is Candida albicans, a ubiquitous fungus that is a part of the normal microbial flora found on mucosal surfaces such as those of the oral cavity, gastrointestinal tract, and vagina in human beings and domestic animals [1]. This yeast is the most common cause of mucosal and invasive fungal infections observed in humans [2]. Host protection against C. albicans infection is complex and includes different subsets of the immune defense system [3-5].
During the development of oropharyngeal candidiasis (OPC), Candida adheres to and invades the tissue. The adhesion of this yeast to host tissue is the initial phase of a potential infection that enables microorganisms to survive inside the host and eventually colonize host tissues during the onset of candidiasis [6,7]. Following this irreversible attachment, fungal cells proliferate and optimally interact to ensure their sustainability in the imposed environment. After the proliferative phase comes the production and deposition of a thick extracellular matrix (mature biofilm) that procures chemical as well as physical protection for cells [8,9].
C. albicans adhesion and growth leading to biofilm formation are mediated by various genes such as HWP1 and EAP1 that encode well-characterized class of cell wall proteins that house a glycosylphosphatidylinositol (GPI) anchorage motif in their C–terminal domains and a signal peptide at their N-termini. These C- and N-terminal domains mediate the adherence of C. albicans to various surfaces [10]. HWP1, a hyphal-specific adhesion gene encodes the hyphal cell wall protein, and both are essential for biofilm formation [11]. The involvement of HWP1 in C. albicans adhesion can be supported by the EAP1 gene which encodes a glucan-cross-linked cell wall protein (adhesin Eap1p) and mediates the adhesion of C. albicans to different surfaces, including epithelial cells and polystyrene [12]. Similar to other genes, HWP1 and EAP1 are downstream effectors of EFG1[13], a transcription regulator [14]. The efg1 mutant strain has been shown to exhibit defects in growth, biofilm formation, and virulence [15].
C. albicans virulence is also mediated by proteolytic enzymes such as secreted aspartyl proteinases (Saps) [16,17]. The contribution of Saps to mucosal and systemic infections and their involvement in adherence, tissue damage, and evasion of host immune responses has been reported, showing the implication of the Sap2 gene in C. albicans growth in protein-containing media [18]. SAP1 and SAP3 are expressed during phenotypic switching [19,20], while SAP4, SAP5 and SAP6 are expressed upon hyphal formation [20]. SAP9 and SAP10 are involved in the mechanism of adhesion to host cells [21]. This proteolytic enzyme family is therefore involved in C. albicans virulence.
C. albicans infection can be promoted by several factors. Candidiasis has been associated with long-term antibiotics intake, AIDS, leukemia, malignancy, radiation therapy for head and neck cancer, or other risk factors that interfere with immunocompetence [22-24].
Environmental factors such as smoking may also promote Candida infections [25,26]. Tobacco smoke exposure has been shown to promote microbial biofilm formation [27]. Specifically, it has been shown that cigarette smoke interferes with S mutans and C. albicans adhesion, resulting in biofilm formation on dental restoration materials [28], which suggests that cigarette smokers are more susceptible to life-threatening oral infections including candidiasis. The aim of this study was to investigate the effect of cigarette smoke condensate on C. albicans adhesion, growth, and biofilm formation, and on the activation of several genes involved in the virulence of this yeast.
Results
Cigarette smoke condensate promoted C albicans adhesion and growth
C. albicans attachment to the surface of glass slides for 1, 3, and 6 h was measured by means of crystal violet staining. As shown in Figure 1C. albicans adhesion was significant (p < 0.05) at 3 and 6 h of incubation. All of the tested CSC concentrations promoted the adhesion of C. albicans. Adhesion was related to incubation period, with low adhesion reported at 1 h and high adhesion at 6 h. This result indicates that CSC can increase C. albicans adhesion and that this effect can lead to significant C. albicans growth. For this purpose, we investigated the effect of CSC on C. albicans growth. As shown in Figure 2, high C. albicans growth was obtained in the presence of CSC, compared to that obtained by the controls. Indeed, C. albicans growth significantly (P < 0.05) increased with as low as 10% CSC. Of interest is that the most effective concentrations of CSC were between 20 and 40%; at these concentrations, C. albicans growth was two to threefold higher than that recorded by the controls. Overall data thus demonstrate that cigarette smoke favors C. albicans adhesion and growth.
Figure 1.
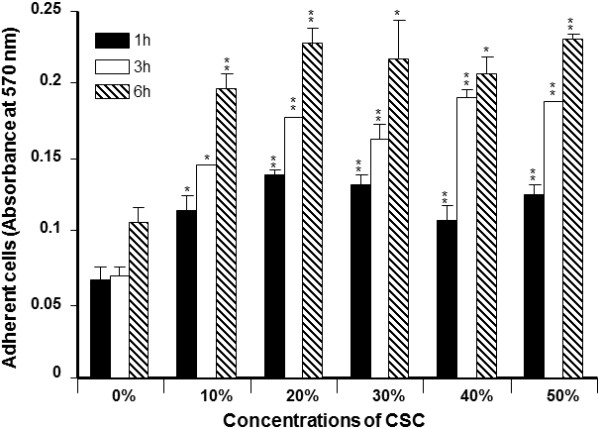
Effect of CSC on C. albicans adhesion. Yeast cells were grown on glass slides with or without various concentrations of CSC for different time points at 30°C. The attached cells were stained with crystal violet and read with an ELISA reader at 570 nm (n = 5). *p < 0.05; **p < 0.01, compared to the non-CSC-cultured C. albicans.
Figure 2.
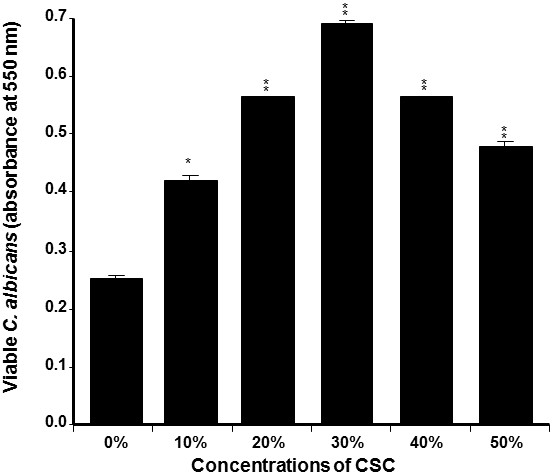
CSC promoted C. albicans growth. Yeast cells were seeded with or without CSC at various concentrations. The cultures were maintained for 24 h at 30°C. The live cells were ascertained by MTT assay (n = 4). *p < 0.05; **p < 0.01, compared to the non-CSC-cultured C. albicans.
Cigarette smoke condensate promoted C albicans biofilm formation
Because CSC contributed to increasing C. albicans adhesion and growth, we tested its potential to promote C. albicans biofilm formation. Using SEM analyses and a crystal violet assay, we were able to demonstrate the stimulatory effect of CSC on biofilm formation (Figure 3). SEM analyses revealed a high C. albicans density in the CSC-treated culture (Figure 3A). A high C. albicans density was observed in the scaffold in the presence of 30% CSC and this density increased with 50% CSC. To confirm these observations, quantitative analyses were conducted using the crystal violet staining method. Figure 3B showed that after 2 days of culture, CSC was able to significantly (p < 0.05) increase biofilm formation. This effect was observed beginning at a concentration of 20% CSC, and at 50% CSC, biofilm formation was greater than that observed in the controls and at 30% CSC.
Figure 3.
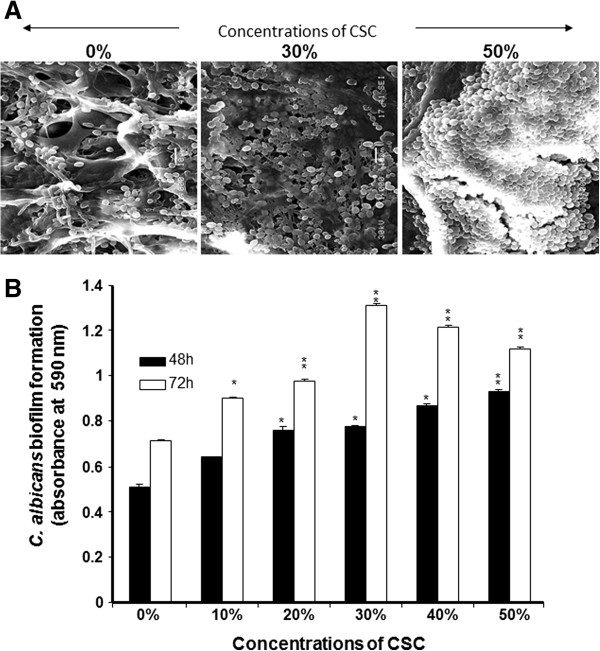
CSC promoted C. albicans biofilm formation. Yeast cells were grown with or without CSC at various concentrations for 48 and 72 h at 30°C. Panel A: Biofilms examined by SEM; photos are presented (n = 4). Panel B: C. albicans-seeded membranes stained with crystal violet. The optical density of each solution was read at 570 nm by means of a plate reader (n = 6). *p < 0.05; **p < 0.01, compared to the non-CSC-cultured C. albicans.
Cigarette smoke condensate modulated HWP1, EAP1, and SAP2 expression
Based on the data showing that CSC increased C. albicans adhesion, growth, and biofilm formation, we sought to determine whether this took place through the regulation of certain genes. Figure 4 reveals that HWP1 gene expression significantly increased following exposure of C. albicans to CSC. The activation of this gene significantly (p < 0.001) increased according to CSC concentration. As shown in Figure 4, a twofold increase in HWP1 gene expression was recorded with a concentration of 30% CSC, compared to that observed in the controls, and with 50% CSC, this increase was over threefold. Similarly, EAP1 gene, which encodes a glycosylphosphatidylinositol-anchored, glucan-cross-linked cell wall protein involved in adhesion and biofilm formation, was also affected by CSC treatment. Figure 5 shows that CSC significantly increased the expression of the EAP1 gene and that this increase was dependent on the concentration of CSC; the higher the concentration, the greater was the expression of this gene. The SAP2 gene was also modulated by CSC. Figure 6 shows that the CSC led to a significant (p < 0.001) increase of SAP2 gene expression. Of interest is the increased Sap2 gene expression with CSC concentration. Indeed, with 30% CSC, a twofold expression was recorded and with 50% CSC this expression increased threefold compared to that observed in the controls (C. albicans not exposed to CSC).
Figure 4.
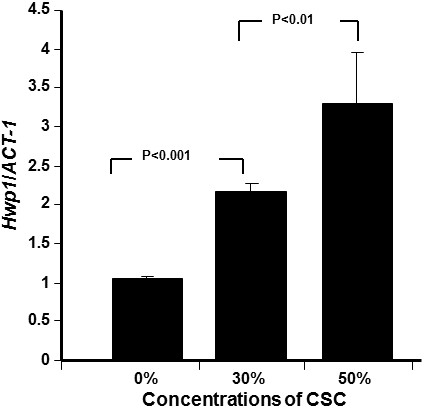
HWP1 mRNA expression by C. albicans increased with CSC stimulation. Yeast cells were grown with or without CSC (30 and 50%) for 24 h. HWP1 gene expression was analyzed by qRT-PCR (n = 5).
Figure 5.
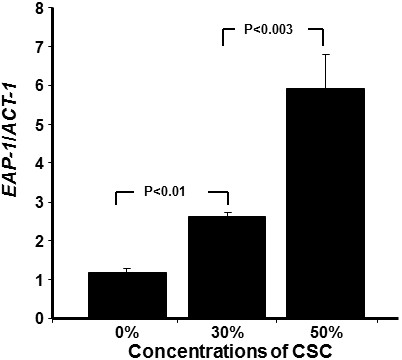
Cigarette smoke condensate induced EAP1 mRNA expression by C. albicans. Yeast cells were grown with or without CSC (30 and 50%) for 24 h. EAP1 gene expression was analyzed by qRT-PCR (n = 6).
Figure 6.
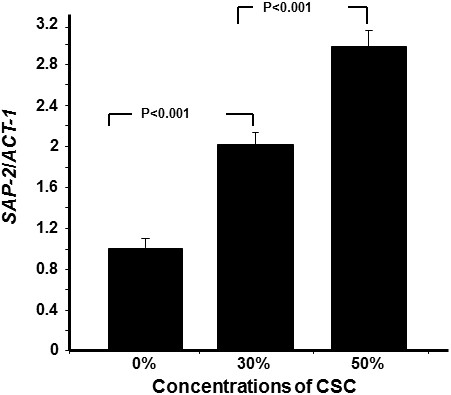
C. albicans expressed high levels of Sap2 mRNA following exposure to CSC. Yeast cells were grown with or without CSC (30 or 50%) for 24 h prior to RNA extraction and Sap2 mRNA expression analysis by qRT-PCR (n = 5).
Discussion
Smoking is known to induce a variety of changes in the oral cavity. Cigarette smoke affects both saliva [29] and oral microorganisms, including C. albicans, a leading cause of oral candidiasis [28]. However, the specific effect of cigarette smoke on C.albicans remains to be elucidated. The first question we addressed in this study was: What is the effect of cigarette smoke on C. albicans adhesion? We demonstrated that CSC promoted C. albicans adhesion, which is in agreement with previously reported studies showing a high rate of oral candida carriage in tobacco smokers compared to non-smokers [30,31]. Furthermore, bacteria exposed to CSC were shown to adhere more to epithelial cells compared to non-exposed specimens [32], which supports our data with C. albicans. Increased adhesion of C. albicans in the presence of CSC may occur due to changes in the interaction of C. albicans with its environment through the expression of high levels of adhesins, as previously suggested [33]. Furthermore, it is important to realize that cigarette smoke comprises a high number of individual compounds [34], including acetaldehyde, benzene, 1,3-butadiene, and isoprene with an elevated mutagenic potential [35]. Thus, it is possible that these compounds may have exerted specific effect on C. albicans adhesion, growth and probably biofilm formation. The effect of cigarette smoke promoting cell adhesion was previously reported by Baboni et al., (2009) showing linear dose response adhesion [25]. The mechanism involved in such effect could involve kinase pathways. These pathways can be promoted by CSC compounds at certain concentration, but inhibited when these compounds are high explaining the decrease of C. albicans adhesion/biofilm formation at 40% and 50% of CSC. Further research is mandatory to shed light on the mechanisms leading to the up-regulation of C. albicans adhesion when exposed to cigarette smoke.
C. albicans adhesion is one of the key events leading to candidiasis [36,37]. This adhesion is usually followed by overgrowth and invasion [38]. Consequently, in promoting C. albicans adhesion, CSC may lead to an over-growth of this yeast. Our study confirms this hypothesis showing a growth increase of C. albicans in the presence of CSC. This concurs with previously published reports showing that smoking can be an important predisposing factor for oral candidiasis [28], which may be enhanced by cigarette smoke through an increased secretion of histolytic enzymes by C. albicans, thus contributing to its virulence [28]. However, the exact pathogenic influence of smoking has yet to be investigated.
C. albicans adhesion and growth are particularly necessary for biofilm formation [39,40]. Because CSC significantly increased C. albicans adhesion and growth in the present study, it is suggested that CSC may also promote C. albicans biofilm formation. Using appropriate conditions to form biofilms, our findings indicate that CSC was indeed capable of promoting biofilm formation. Of interest is that a significant increase of biofilm formation was obtained at both tested concentrations, and that this phenomenon was dependent on CSC concentration. These useful data are comparable to those of other studies showing increased microbial biofilm formation with cigarette smoke [41-43]. By showing the significant stimulatory effect on increasing C. albicans biofilm formation, cigarette smoke can thus be labeled as an infection-promoting agent.
Promoting C. albicans adhesion, growth, and biofilm formation may operate through the modulated expression of certain C. albicans genes [44,45], as supported by our study demonstrating that CSC led to a high expression of the EAP1 gene. As a member of the GPI-CWP family in C. albicans[46], Eap1p was originally identified because of its ability to mediate adhesion to polystyrene when the EAP1 gene was expressed in a flocculin-deficient Saccharomyces cerevisiae strain. EAP1 expression in a C. albicans efg1/efg1 mutant was able to restore C. albicans adhesion to epithelial cells [12]. Deleting EAP1 in C. albicans was shown to reduce cell adhesion to polystyrene and to epithelial cells in a gene dosage-dependent manner [12,46]. Indeed, this suggests that exposure to CSC increases EAP1 expression, which may in turn contribute to increasing C. albicans adhesion, and ultimately, biofilm formation and pathogenesis.
We also demonstrated that CSC increased HWP1 mRNA expression. HWP1 is a downstream component of the cAMP-dependent PKA pathway and is positively regulated by EFG1[47]. The transcript level of HWP1 increased with increasing CSC stimulation, which suggests that CSC did affect cAMP–EFG1 pathway activity, resulting in an increase of C. albicans adhesion and growth with biofilm formation. Further investigations are therefore warranted to gain greater insight into the interaction between cigarette smoke and C. albicans leading to infection.
Candida pathogenesis is associated with the production and secretion of histolytic enzymes [48]. Secreted aspartyl proteases (Saps) and phospholipases were specifically reported as being directly related to C. albicans virulence [49]. During infection, Saps are incriminated degrading host proteins involved in tissue barriers and immune defense [18,50,51]. Here, we report that CSC upregulated Sap2 mRNA expression. It is known that Sap gene upregulation contributes to increasing C. albicans transition, and later, its pathogenicity through an augmented secretion of proteinases [52]. Our study thus establishes, for the first time, a clear link between cigarette smoke and C. albicans pathogenesis through the behavior of key genes such as EAP1, HWP1 and Sap2. These genes are known to be involved in controlling Candida adhesion, growth, and biofilm formation [53]; however, the precise interactions between these different genes and cigarette smoke during C. albicans pathogenesis have not yet been fully investigated. Data suggest that gene activation can be involved in C. albicans adhesion, growth and biofilm formation promoted by CSC. This may involve kinase pathways contributing to C. albicans adaptation to the CSC environment as previously suggested [25].
Conclusions
This study demonstrated that CSC upregulates C. albicans adhesion and growth that promote biofilm formation. Of interest is that these effects were supported by the modulation of C. albicans genes EAP1, HWP1, and Sap2. Overall results therefore suggest a possible link between cigarette smoke, C. albicans activation, and oral candidiasis.
Methods
Preparation of cigarette smoke condensate
1R3F cigarettes were purchased from the Kentucky Tobacco Research & Development Center (Orlando, FL) and were used to prepare the cigarette smoke condensate solution, as shown in Figure 7. Each cigarette was placed into one end of a silicone tube linked to an Erlenmeyer flask containing 200 ml of 0.09% sodium chloride. On the other end, a second silicone tube linked to the Erlenmeyer was connected to a standard vacuum. The cigarette was attached to the cigarette holder and lit and the smoke was extracted by applying vacuum, pulling the smoke directly into the 0.09% sodium chloride solution. The process was repeated for a total of ten whole cigarettes. The resulting cigarette smoke condensate (CSC) solution was then sterilized by filtration through a 0.22 μm filter and stored at 4°C until use.
Figure 7.
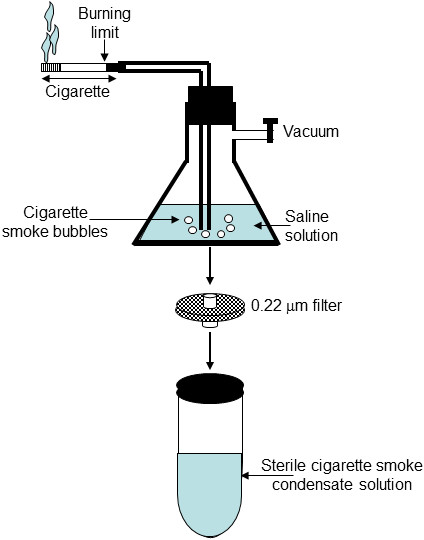
Generation of sterile CSC. Smoke from a burning cigarette was passed through a 0.9% saline solution. The CSC-rich solution was then sterilized by filtration through 0.22-um sterile filters and later stored at -20°C until use.
Candida strain
C. albicans SC5314 was cultured for 24 h on Sabouraud dextrose agar plates (Becton Dickinson, Oakville, ON, Canada) at 30°C. For the C. albicans suspensions, one colony was used to inoculate 10 ml of Sabouraud liquid medium supplemented with 0.1% glucose, pH 5.6. The cultures were then grown to the stationary phase in a shaking water bath for 18 h at 30°C, after which time the yeast cells were collected, washed with phosphate-buffered saline (PBS), counted by means of a haemocytometer, and adjusted to 106/ml prior to use.
Effect of CSC on C albicans adhesion
A glass slide (0.5 cm in diameter) was placed into each well of a 12-well plate then covered with 1 ml of sterile artificial saliva solution at room temperature for 30 min under agitation to ensure that the slides were covered by liquid at all times. Before use, the artificial saliva (2.5 g of NaCl, 332.97 mg of CaCl2, 250 mg of MgCl2.6H2O, 189.48 mg of KCl, 3 g of anhydrous potassium acetate (C2H3O2K), 772.00 mg of K3PO4.3 H2O, and 0.1 ml of H3PO4 (85 wt. % in H2O, Sigma Aldrich)) was supplemented with 140 mg of Type II mucin in 1000 ml of the prepared saliva solution, pH 7. Following the saliva coating, the glass slides were gently rinsed with sterile saline, transferred to new wells of sterile 12-well plates, and subsequently dried for 3 h under a sterile culture hood. Each saliva-coated glass slide was then covered with 104C. albicans cells in 50 μl of Sabouraud medium supplemented or not with CSC at various concentrations (10, 20, 30, 40 or 50%) and incubated for 60 nm at 30°C under stable conditions to prevent the medium from leaking from the slide onto the plastic. Following this incubation period, 1 ml of Sabouraud with or without CSC was added to each well and the cultures were further incubated for 1, 3, or 6 h. At the end of each incubation period, each slide was removed from its well and placed into new wells of a 12-well plate, subsequently washed twice with warm PBS, and subjected to crystal violet staining. One milliliter of 1% w/v crystal violet solution in demineralized water was added, and the slides were further incubated at room temperature for 30 min. After incubation, the non-bound dye was removed from the wells by thorough washing with demineralized water, followed by drying at 37°C. Bound crystal violet was dissolved by adding 1 ml of absolute ethanol and incubating on a rocking platform for 15 min at room temperature. The absorbance levels of the dissolved dye were measured at a wavelength of 590 nm by means of an optical density reader (X-Mark Microplate Spectrophotometer, Bio-Rad Laboratories, Mississauga, ON, Canada).
Effect of CSC on C albicans growth
The yeast was seeded into separate tubes (105C. albicans per tube) in 4 ml of Sabouraud culture medium supplemented or not with CSC at 10, 20, 30, 40, or 50%. The cultures were maintained at 30°C for 24 h, after which time C. albicans growth was evaluated by means of the (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide) (MTT) assay (Sigma-Aldrich, St. Louis, MO) which measures viable cells as a function of mitochondrial activity. Briefly, an MTT stock solution (5 mg/ml) was prepared in PBS and added to each culture at a final concentration of 10% (v/v). The C. albicans cultures were then incubated with the MTT solution at 30°C for 4 h, after which time the plate was centrifuged for 10 min at 1200 rpm and the supernatant was removed. The remaining pellet in each condition was then washed with warm PBS, and 2 ml of 0.04 N HCl in isopropanol were added to each pellet, with a further incubation for 15 min. Absorbance (optical density, OD) was subsequently measured at 550 nm by means of an xMark microplate spectrophotometer (Bio-Rad, Mississauga, ON, Canada).
Effect of CSC on C albicans biofilm formation
C. albicans biofilms were obtained by culturing the yeast on a porous collagen scaffold which facilitated C. albicans penetration through the pores and its adhesion to the scaffold through collagen affinity. This also promoted biofilm formation and handling with no cell loss, thus contributing to the maintenance of the biofilm structure. For this purpose, 5 mm × 5 mm samples of porous scaffold (Collatape, Zimmer Dental Inc., Carlsbad, CA, USA) were placed in a 24-well plate, rinsed twice with culture medium, seeded with C. albicans (106 cells), and finally incubated for 30 min at 30°C without shaking to allow for adherence. Fresh Sabouraud medium was added to each well in the presence or absence of various concentrations of CSC (10, 30, or 50%). The C. albicans-seeded scaffolds were then incubated for 2 or 3 days at 30°C. Following each culture period, C. albicans growth and biofilm formation was assessed by scanning electron microscopy and the crystal violet assay.
Scanning electron microscopy (SEM) analysis
C. albicans-rich scaffolds (biofilms) were fixed in ethylene glycol for 60 min followed by a rinse with sterile PBS. Dehydration was performed in a series of 5-min treatments with ethanol solutions of increasing concentration (50, 70, 90, and twice at 100%). The dehydrated biofilms were kept overnight in a vacuum oven at 25°C, after which time they were sputter-coated with gold, examined, and imaged under a JEOL 6360 LV SEM (Soquelec, Montréal, QC, Canada) operating at a 30 kV accelerating voltage.
Biofilm staining by crystal violet
Following incubation for 24 h at 30°C, biofilms were stained with 1% crystal violet (100 μl) for 15 min. They were then washed three times with PBS to remove unbound crystal violet dye and were dried overnight at room temperature. The biofilms were then covered with 1 ml of absolute ethanol and were incubated on a rocking platform for 20 min at room temperature to release the stain from the biofilms. Absorbance was recorded at 590 nm. Each biofilm assay was run in triplicate and the means ± standard deviations of four separate experiments were calculated and plotted.
Effect of CSC on C albicans gene activation/repression
C. albicans (105 cells) was cultured in the presence or absence of CSC at various concentrations (30 and 50%) at 30°C for 24 h under agitation. At the end of this incubation period, the cultures were centrifuged 10 min at 13,000 rpm, the supernatants were discarded, and each pellet was suspended in 0.6 ml of lysis buffer (1 M glycerol, 0.1 M EDTA). Glass beads (0.425-0.6 mm in diameter; 0.2 ml) were then added to each suspended pellet prior to sonication (4 × 1 min, followed by 2 min of incubation in ice) by means of a MiniBead-beater (Biospec Products, Bartlesville, OK, USA). Following cell lysis, the total RNA was extracted from each sample by means of the Illustra RNAspin Mini kit (GE Health Care UK Limited, Buckingham, UK). The concentration, purity, and quality of the extracted RNA were determined using the Experion system and the RNA StdSens analysis kit according to instructions provided by the manufacturer (Bio-Rad, Hercules, CA, USA). Appropriate RNAs were used to perform quantitative RT-PCR.
Quantitative real-time RT-PCR
RNA (500 ng of each sample) was reverse transcribed into cDNA by means of the iScript cDNA synthesis kit (Bio-Rad, Mississauga, ON, Canada). The conditions for the preparation of the cDNA templates for PCR analysis were 5 min at 25°C, 1 h at 42°C, and 5 min at 85°C. Quantitative PCR (qPCR) was carried out. The quantity of mRNA transcripts was measured with the Bio-Rad CFX96 real-time PCR detection system. Reactions were performed using a PCR supermix (iQ SYBR Green Supermix, Bio-Rad). Primers (Table 1) were added to the reaction mix to a final concentration of 250 nM. Five microliters of each cDNA sample was added to a 20 μl PCR mixture containing 12.5 μl of the iQ SYBR Green supermix, 0.5 μl of specific primers ACT1, SAP2, HWP1, and EAP1 (Invitrogen Life Technologies Inc., Burlington, ON, Canada), and 7 μl of RNase/DNase-free water (MP Biomedicals, Solon, OH, USA). Each reaction was performed in a Bio-Rad MyCycler thermal cycler. For the qPCR, the CT was automatically determined by the accompanying Bio-Rad CFX manager. Thermocycling conditions for the ACT1, Sap2, and EAP1 were established at 5 min at 95°C, followed by 30 cycles of 15 s at 95°C, 30 s at 60°C, and 30 s at 72°C, with each reaction performed in triplicate. For the HWP1, the thermocycling conditions were 3 min at 95°C, followed by 30 cycles of 10 s at 95°C, 30 s at 54°C, and 40 s at 72°C, with each reaction also performed in triplicate. The specificity of each primer pair was determined by the presence of a single melting temperature peak. The ACT1 produced uniform expression levels varying by less than 0.5 CTs between sample conditions and thus became the reference gene for this study. The results were analyzed using the 2-ΔΔCt (Livak) relative expression method.
Table 1.
Primer sequences used for the qRT-PCR
| Gene | Primer sequence 5′ to 3′ | Amp size (bp) |
|---|---|---|
|
ACT1
|
Forward: GCTGGTAGAGACTTGACCAACCA |
87 |
| Reverse: GACAATTTCTCTTTCAGCACTAGTAGTGA | ||
|
EAP1
|
Forward: CTGCTCACTCAACTTCAATTGTCG |
51 |
| Reverse: GAACACATCCACCTTCGGGA | ||
|
HWP1
|
Forward: GCTCAACTTATTGCTATCGCTTATTACA |
67 |
| Reverse: GACCGTCTACCTGTGGGACAGT | ||
|
SAP2
|
Forward: TCCTGATGTTAATGTTGATTGTCAAG |
82 |
| Reverse: TGGATCATATGTCCCCTTTTGTT |
Statistical analysis
Each experiment was performed at least four times, with experimental values expressed as means ± SD. The statistical significance of the differences between the control (absence of CSC) and test (presence of CSC) values was determined by means of a one-way ANOVA. Posteriori comparisons were performed using Tukey’s method. Normality and variance assumptions were verified using the Shapiro-Wilk test and the Brown and Forsythe test, respectively. All of the assumptions were fulfilled. P values were declared significant at ≤ 0.05. The data were analyzed using the SAS version 8.2 statistical package (SAS Institute Inc., Cary, NC, USA).
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
MR and AS conceived the study. AS, KK and HA conducted the experiments. KK, AS, HA, WC, and MR analyzed and interpreted the data. AS, KK and HA drafted the Materials and Methods section. MR completed the manuscript with the help of WC. All of the authors read and approved the final manuscript.
Contributor Information
Abdelhabib Semlali, Email: asemlali@KSU.EDU.SA.
Kerstin Killer, Email: kkerstin.killer@gmail.com.
Humidah Alanazi, Email: hameeda-7@hotmail.com.
Witold Chmielewski, Email: Witold.Chmielewski@fmd.ulaval.ca.
Mahmoud Rouabhia, Email: mahmoud.rouabhia@fmd.ulaval.ca.
Acknowledgements
This research was supported by NSERC discovery grant number-155234-07, as well as a grant from the Fonds Émile-Beaulieu (Laval University). The authors also thank Ms. Claire Kingston (Traduction CFK) for proofreading and editing this manuscript.
References
- Richardson M, Lass-Florl C. Changing epidemiology of systemic fungal infections. Clin Microbiol Infect. 2008;14:5–24. doi: 10.1111/j.1469-0691.2008.01978.x. [DOI] [PubMed] [Google Scholar]
- Morschhauser J. Regulation of multidrug resistance in pathogenic fungi. Fungal Genet Biol. 2010;47:94–106. doi: 10.1016/j.fgb.2009.08.002. [DOI] [PubMed] [Google Scholar]
- Rouabhia M, Ross G, Page N, Chakir J. Interleukin-18 and gamma interferon production by oral epithelial cells in response to exposure to Candida albicans or lipopolysaccharide stimulation. Infect Immun. 2002;70:7073–7080. doi: 10.1128/IAI.70.12.7073-7080.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rouabhia M, Schaller M, Corbucci C, Vecchiarelli A, Prill SK, Giasson L, Ernst JF. Virulence of the fungal pathogen Candida albicans requires the five isoforms of protein mannosyltransferases. Infect Immun. 2005;73:4571–4580. doi: 10.1128/IAI.73.8.4571-4580.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saunus JM, Kazoullis A, Farah CS. Cellular and molecular mechanisms of resistance to oral Candida albicans infections. Front Biosci. 2008;13:5345–5358. doi: 10.2741/3085. [DOI] [PubMed] [Google Scholar]
- Park H, Liu Y, Solis N. Transcriptional responses of Candida albicans to epithelial and endothelial cells. Eukaryot Cell. 2009;8:1498–1510. doi: 10.1128/EC.00165-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zakikhany K, Naglik JR, Schmidt-Westhausen A, Holland G, Schaller M, Hube B. In vivo transcript profiling of Candida albicans identifies a gene essential for inter-epithelial dissemination. Cell Microbiol. 2007;9:2938–2954. doi: 10.1111/j.1462-5822.2007.01009.x. [DOI] [PubMed] [Google Scholar]
- Costerton JWS, Stewart PS, Greenberg EP. Bacterial biofilms: a common cause of persistent infections. Science. 1999;284:1318–1322. doi: 10.1126/science.284.5418.1318. [DOI] [PubMed] [Google Scholar]
- Grant SS, Hung DT. Persistent bacterial infections, antibiotic tolerance, and the oxidative stress response. Virulence. 2013;4:273–283. doi: 10.4161/viru.23987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sundstrom P. Adhesion in Candida spp. Cell Microbiol. 2002;4:461–469. doi: 10.1046/j.1462-5822.2002.00206.x. [DOI] [PubMed] [Google Scholar]
- Nobile CJ, Nett JE, Andes DR, Mitchell AP. Function of Candida albicans adhesin Hwp1 in biofilm formation. Eukaryot Cell. 2006;5:1604–1610. doi: 10.1128/EC.00194-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li F, Palecek SP. EAP1, a Candida albicans gene involved in binding human epithelial cells. Eukaryot Cell. 2003;2:1266–1273. doi: 10.1128/EC.2.6.1266-1273.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sohn K, Urban C, Brunner H, Rupp S. EFG1 is a major regulator of cell wall dynamics in Candida albicans as revealed by DNA microarrays. Mol Microbiol. 2003;47:89–102. doi: 10.1046/j.1365-2958.2003.03300.x. [DOI] [PubMed] [Google Scholar]
- Stoldt VR, Sonneborn A, Leuker CE, Ernst JF. Efg1p, an essential regulator of morphogenesis of the human pathogen Candida albicans, is a member of a conserved class of bHLH proteins regulating morphogenetic processes in fungi. EMBO J. 1997;16:1982–1991. doi: 10.1093/emboj/16.8.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lo HJ, Köhler JR, DiDomenico B, Loebenberg D, Cacciapuoti A, Fink GR. Nonfilamentous C albicans mutants are avirulent. Cell. 1997;90:939–949. doi: 10.1016/S0092-8674(00)80358-X. [DOI] [PubMed] [Google Scholar]
- Schaller M, Borelli C, Korting HC, Hube B. Hydrolytic enzymes as virulence factors of Candida albicans. Mycoses. 2005;48:365–377. doi: 10.1111/j.1439-0507.2005.01165.x. [DOI] [PubMed] [Google Scholar]
- Décanis N, Tazi N, Correia A, Vilanova M, Rouabhia M. Farnesol, a fungal quorum-sensing molecule triggers Candida albicans morphological changes by downregulating the expression of different secreted aspartyl proteinase genes. Open Microbiol J. 2011;5:119–126. doi: 10.2174/1874285801105010119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naglik JR, Challacombe SJ, Hube B. Candida albicans secreted aspartyl proteinases in virulence and pathogenesis. Microbiol Mol Biol Rev. 2003;67:400–428. doi: 10.1128/MMBR.67.3.400-428.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hube B, Naglik J. Candida albicans proteinases: resolving the mystery of a gene family. Microbiology. 2001;147:1997–2005. doi: 10.1099/00221287-147-8-1997. [DOI] [PubMed] [Google Scholar]
- White TC, Agabian N. Candida albicans secreted aspartyl proteinases: isoenzyme pattern is determined by cell type, and levels are determined by environmental factors. J Bacteriol. 1995;177:5215–5221. doi: 10.1128/jb.177.18.5215-5221.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albrecht A, Felk A, Pichova I, Naglik JR, Schaller M, de Groot P, Maccallum D, Odds FC, Schäfer W, Klis F, Monod M, Hube B. Glycosylphosphatidylinositol-anchored proteases of Candida albicans target proteins necessary for both cellular processes and host-pathogen interactions. J Biol Chem. 2006;281:688–694. doi: 10.1074/jbc.M509297200. [DOI] [PubMed] [Google Scholar]
- Nave R, Mueller H. From inhaler to lung: clinical implications of the formulations of ciclesonide and other inhaled corticosteroids. Int J Gen Med. 2013;6:99–107. doi: 10.2147/IJGM.S39134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinel B, Cassou-Mounat T, Bensadoun RJ. Oropharyngeal candidiasis and radiotherapy. Cancer Radiother. 2012;3:222–229. doi: 10.1016/j.canrad.2011.11.004. [DOI] [PubMed] [Google Scholar]
- Fidel PL Jr. Candida-host interactions in HIV disease: implications for oropharyngeal candidiasis. Adv Dent Res. 2011;1:45–49. doi: 10.1177/0022034511399284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baboni FB, Barp D, Izidoro AC, Samaranayake LP, Rosa EA. Enhancement of Candida albicans virulence after exposition to cigarette mainstream smoke. Mycopathologia. 2009;168(5):227–235. doi: 10.1007/s11046-009-9217-5. [DOI] [PubMed] [Google Scholar]
- Soysa NS, Ellepola AN. The impact of cigarette/tobacco smoking on oral candidosis: an overview. Oral Dis. 2005;5:268–273. doi: 10.1111/j.1601-0825.2005.01115.x. [DOI] [PubMed] [Google Scholar]
- Goldstein-Daruech N, Cope EK, Zhao KQ, Vukovic K, Kofonow JM, Doghramji L, González B, Chiu AG, Kennedy DW, Palmer JN, Leid JG, Kreindler JL, Cohen NL. Tobacco smoke mediated induction of sinonasal microbial biofilms. PLoS One. 2011;6:e15700. doi: 10.1371/journal.pone.0015700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baboni FB, Guariza Filho O, Moreno AN, Rosa EA. Influence of cigarette smoke condensate on cariogenic and candidal biofilm formation on orthodontic materials. Am J Orthod Dentofacial Orthop. 2010;4:427–434. doi: 10.1016/j.ajodo.2009.05.023. [DOI] [PubMed] [Google Scholar]
- Weiner D, Khankin EV, Levy Y, Aizenbud D, Reznick AZ. Effects of cigarette smoke on salivary protein tyrosine nitration. Eur J Med Res. 2010;2:211–216. doi: 10.1186/2047-783X-15-S2-211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abu-Elteen KH, Abu-Alteen RM. The prevalence of Candida albicans populations in the mouths of complete denture wearers. New Microbiol. 1998;21(1):41–48. [PubMed] [Google Scholar]
- Willis AM, Coulter WA, Fulton CR, Hayes JR, Bell PM, Lamey PJ. Oral candidal carriage and infection in insulin-treated diabetic patients. Diabet Med. 1999;16(8):675–679. doi: 10.1046/j.1464-5491.1999.00134.x. [DOI] [PubMed] [Google Scholar]
- Teughels W, Van Eldere J, van Steenberghe D, Cassiman JJ, Fives-Taylor P, Quirynen M. Influence of nicotine and cotinine on epithelial colonization by periodontopathogens. J Periodontol. 2005;8:1315–1322. doi: 10.1902/jop.2005.76.8.1315. [DOI] [PubMed] [Google Scholar]
- Feldman C, Anderson R. Cigarette smoking and mechanisms of susceptibility to infections of the respiratory tract and other organ systems. J Infect. 2013;3:169–184. doi: 10.1016/j.jinf.2013.05.004. [DOI] [PubMed] [Google Scholar]
- Gaworski CL, Dozier MM, Eldridge SR, Morrissey R, Rajendran N, Gerhart JM. Cigarette smoke vapor-phase effects on the rat upper respiratory tract. Inhalation Toxicol. 1998;10:857–873. doi: 10.1080/089583798197420. [DOI] [Google Scholar]
- Adam T, Mitschke S, Streibel T, Baker RR, Zimmermann R. Quantitative puff-by-puff-resolved characterization of selected toxic compounds in cigarette mainstream smoke. Chem Res Toxicol. 2006;19:511–520. doi: 10.1021/tx050220w. [DOI] [PubMed] [Google Scholar]
- Akerey B, Le-Lay C, Fliss I, Subirade M, Rouabhia M. In vitro efficacy of nisin Z against Candida albicans adhesion and transition following contact with normal human gingival cells. J Appl Microbiol. 2009;107:1298–1307. doi: 10.1111/j.1365-2672.2009.04312.x. [DOI] [PubMed] [Google Scholar]
- Kempf M, Cottin J, Licznar P, Lefrançois C, Robert R, Apaire-Marchais V. Disruption of the GPI protein-encoding gene IFF4 of Candida albicans results in decreased adherence and virulence. Mycopathologia. 2009;168:73–77. doi: 10.1007/s11046-009-9201-0. [DOI] [PubMed] [Google Scholar]
- Finkel JS, Xu W, Huang D, Hill EM, Desai JV, Woolford CA, Nett JE, Taff H, Norice CT, Andes DR, Lanni F, Mitchell AP. Portrait of Candida albicans adherence regulators. PLoS Pathog. 2012;2:e1002525. doi: 10.1371/journal.ppat.1002525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inglis DO, Skrzypek MS, Arnaud MB, Binkley J, Shah P, Wymore F, Sherlock G. Improved gene ontology annotation for biofilm formation, filamentous growth, and phenotypic switching in Candida albicans. Eukaryot Cell. 2013;12(1):101–108. doi: 10.1128/EC.00238-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silva S, Negri M, Henriques M, Oliveira R, Williams DW, Azeredo J. Adherence and biofilm formation of non-Candida albicans Candida species. Trends Microbiol. 2011;19(5):241–247. doi: 10.1016/j.tim.2011.02.003. [DOI] [PubMed] [Google Scholar]
- Kulkarni R, Antala S, Wang A, Amaral FE, Rampersaud R, Larussa SJ, Planet PJ, Ratner AJ. Cigarette smoke increases staphylococcus aureus biofilm formation via oxidative stress. Infect Immun. 2012;80(11):3804–3811. doi: 10.1128/IAI.00689-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mutepe ND, Cockeran R, Steel HC, Theron AJ, Mitchell TJ, Feldman C, Anderson R. Effects of cigarette smoke condensate on pneumococcal biofilm formation and pneumolysin. Eur Respir J. 2013;41(2):392–395. doi: 10.1183/09031936.00213211. [DOI] [PubMed] [Google Scholar]
- Bagaitkar J, Daep CA, Patel CK, Renaud DE, Demuth DR, Scott DA. Tobacco smoke augments porphyromonas gingivalis-streptococcus gordonii biofilm formation. PLoS One. 2011;6(11):e27386. doi: 10.1371/journal.pone.0027386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutiérrez-Escribano P, Zeidler U, Suárez MB, Bachellier-Bassi S, Clemente-Blanco A, Bonhomme J, de Aldana CR V, d’Enfert C, Correa-Bordes J. The NDR/LATS kinase Cbk1 controls the activity of the transcriptional regulator Bcr1 duringbiofilm formation in Candida albicans. PLoS Pathog. 2012;8(5):e1002683. doi: 10.1371/journal.ppat.1002683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rouabhia M, Semlali A, Chandra J, Mukherjee P, Chmielewski W, Ghannoum MA. Disruption of the ECM33 gene in Candida albicans prevents biofilm formation, engineered human oral mucosa tissue damage and gingival cell necrosis/apoptosis. Mediators Inflamm. 2012;2012:398207. doi: 10.1155/2012/398207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li F, Svarovsky MJ, Karlsson AJ, Wagner JP, Marchillo K, Oshel P, Andes D, Palecek SP. Eap1p, an adhesin that mediates Candida albicans biofilm formation in vitro and in vivo. Eukaryot Cell. 2007;6:931–939. doi: 10.1128/EC.00049-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharkey LL, McNemar MD, Saporito-Irwin SM, Sypherd PS, Fonzi WA. HWP1 functions in the morphological development of Candida albicans downstream of EFG1, TUP1, and RBF1. J Bacteriol. 1999;181:5273–5279. doi: 10.1128/jb.181.17.5273-5279.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dostál J, Hamal P, Pavlícková L, Soucek M, Ruml T, Pichová I, Hrusková-Heidingsfeldová O. Simple method for screening Candida species isolates for the presence of secreted proteinases: a tool for the prediction of successful inhibitory treatment. J Clin Microbiol. 2003;41(2):712–716. doi: 10.1128/JCM.41.2.712-716.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kantarcioglu AS, Yücel A. Phospholipase and protease activities in clinical Candida isolates with reference to the sources of strains. Mycoses. 2002;45(5–6):160–165. doi: 10.1046/j.1439-0507.2002.00727.x. [DOI] [PubMed] [Google Scholar]
- Kaminishi H, Miyaguchi H, Tamaki T, Suenaga N, Hisamatsu M, Mihashi I, Matsumoto H, Maeda H, Hagihara Y. Degradation of humoral host defense by Candida albicans proteinase. Infect Immun. 1995;63:984–988. doi: 10.1128/iai.63.3.984-988.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rouabhia M, Semlali A, Audoy J, Chmielewski W. Antagonistic effect of Candida albicans and IFNγ on E-cadherin expression and production by human primary gingival epithelial cells. Cell Immunol. 2012;280(1):61–67. doi: 10.1016/j.cellimm.2012.11.008. [DOI] [PubMed] [Google Scholar]
- Naglik J, Albrecht A, Bader O, Hube B. Candida albicans proteinases and host/pathogen interactions. Cell Microbiol. 2004;6(10):915–926. doi: 10.1111/j.1462-5822.2004.00439.x. [DOI] [PubMed] [Google Scholar]
- Samaranayake YH, Cheung BP, Yau JY, Yeung SK, Samaranayake LP. Human serum promotes Candida albicans biofilm growth and virulence gene expression on silicone biomaterial. PLoS One. 2013;8(5):e62902. doi: 10.1371/journal.pone.0062902. [DOI] [PMC free article] [PubMed] [Google Scholar]