

Abstract
Sequestration of Plasmodium falciparum-infected erythrocytes (Pf-iEs) in the microvasculature of vital organs plays a key role in the pathogenesis of life-threatening malaria complications, such as cerebral malaria and malaria in pregnancy. This phenomenon is marked by the cytoadhesion of Pf-iEs to host receptors on the surfaces of endothelial cells, on noninfected erythrocytes, and in the placental trophoblast; therefore, these sites are potential targets for antiadhesion therapies. In this context, glycosaminoglycans (GAGs), including heparin, have shown the ability to inhibit Pf-iE cytoadherence and growth. Nevertheless, the use of heparin was discontinued due to serious side effects, such as bleeding. Other GAG-based therapies were hampered due to the potential risk of contamination with prions and viruses, as some GAGs are isolated from mammals. In this context, we investigated the effects and mechanism of action of fucosylated chondroitin sulfate (FucCS), a unique and highly sulfated GAG isolated from the sea cucumber, with respect to P. falciparum cytoadhesion and development. FucCS was effective in inhibiting the cytoadherence of Pf-iEs to human lung endothelial cells and placenta cryosections under static and flow conditions. Removal of the sulfated fucose branches of the FucCS structure virtually abolished the inhibitory effects of FucCS. Importantly, FucCS rapidly disrupted rosettes at high levels, and it was also able to block parasite development by interfering with merozoite invasion. Collectively, these findings highlight the potential of FucCS as a candidate for adjunct therapy against severe malaria.
INTRODUCTION
Malaria still accounts for nearly one million deaths annually (1), and it is often associated with severe complications, such as cerebral malaria (CM) and malaria in pregnancy (MiP). MiP affects 125 million women each year and results in 200,000 infant deaths (2, 3). Despite recent advances in malaria treatment, CM still has mortality rates of 15 to 20%, even after appropriate antimalarial treatment (4, 5), and pregnant women are also at risk for additional complications (e.g., CM) (6).
High levels of proinflammatory cytokines are commonly observed in patients with CM (7, 8) and MiP (9–11), and pathological studies have revealed sequestration of Plasmodium falciparum-infected erythrocytes (Pf-iEs) in the brain microvasculature (12, 13) and in the placenta (14–16). Recently, sequestration of Pf-iEs in the brain microvasculature has been associated with severe CM and death (17). Additionally, poor outcomes for both mother and fetus in MiP are related to greater amounts of Pf-iEs sequestered in the placenta (18, 19).
Sequestration results in the removal of mature Pf-iE forms (>20 h after reinvasion) from the peripheral circulation, as a consequence of the adhesion of parasitized cells to the capillary and postcapillary endothelium and to placental trophoblasts (15, 16, 20, 21). Specific adhesion of Pf-iEs during the early ring stage has also been observed in vitro (22). Another major adhesion phenotype is rosetting, which results from the binding of noninfected erythrocytes (niEs) to Pf-iEs and has been associated with all severe malarial syndromes (23), including CM (23–28). In contrast, no significant correlation between rosetting and disease severity has been reported for isolates from Papua, New Guinea (29), indicating that geographic variations in parasite strains matter (29). Recently, a pivotal role for cytoadherence through the endothelial protein C receptor in the development of severe malaria was described (30). Cytoadhesion of Pf-iEs is mediated by members of the P. falciparum membrane protein 1 (PfEMP-1) family, which mediates parasite interactions with various host receptors, including CD36 (31), intercellular adhesion molecule 1 (ICAM-1) (32), and chondroitin sulfate A (CSA) (33, 34), a receptor frequently associated with MiP (15, 33, 35)
Glycosaminoglycans (GAGs), including heparin, have been employed as a strategy to prevent malaria complications due to their abilities to inhibit parasite cytoadhesion, to block invasion, and to disrupt rosettes (36–46). However, the side effects of heparin, mostly serious bleeding (47), and the potential risk of contamination (because some GAGs are obtained from mammals) have hampered GAG-based adjunct therapies. Recently, we have shown that fucosylated chondroitin sulfate (FucCS), a GAG isolated from the marine invertebrate sea cucumber, contains a central chondroitin sulfate backbone like mammalian GAGs; however, FucCS has sulfated fucose branches (48–50), displays anticoagulant activity (48), and prevents thrombosis (51). FucCS also reduces the recruitment of inflammatory cells, most likely by inhibiting cell adhesion to the host endothelium (52). Here, we investigate the effects of FucCS on the adhesion of Pf-iEs to endothelial cells and placenta cryosections, as well as its abilities to disrupt rosettes and to block parasite invasion.
MATERIALS AND METHODS
Native and chemically modified FucCS.
Native fucosylated chondroitin sulfate was extracted from the body wall of sea cucumbers (Ludwigothurea grisea), which were freshly collected from Guanabara Bay (Rio de Janeiro, Brazil). FucCS was extracted by papain digestion, and purification was performed as described previously (49, 50). The FucCS backbone is made up of repeating disaccharide units of alternating β-d-glucuronic acid and N-acetyl-β-d-galactosamine, which is the same structure as mammalian chondroitin sulfate (Fig. 1A). Partial removal of sulfated fucose branches from the fucosylated chondroitin sulfate, yielding defucosylated chondroitin sulfate (deFucCS) (Fig. 1B), was achieved by mild acid hydrolysis, as described elsewhere (48). Carboxyl-reduced chondroitin sulfate (CRFucCS) was obtained from reduction of the hexuronic acid carboxyl groups in the polysaccharide with 1-ethyl-3-(3-imethylaminopropyl)carbodiimide-NaBH4 (50, 53) (Fig. 1C).
FIG 1.

Chemical structures of fucosylated chondroitin sulfate and its modified forms. (A) Native FucCS. (B) Defucosylated chondroitin sulfate. (C) Carboxyl-reduced chondroitin sulfate. FucCS contains a central core with a chondroitin sulfate-like structure and branches of 2,4-disulfated fucose branches. These branches are easily removed by mild acid hydrolysis, yielding the defucosylated form. The glucuronic acid units found in the central core may be reduced and form glucose units in the carboxyl-reduced form of FucCS.
Cytotoxicity assays.
FucCS cytotoxicity was evaluated in hepatocellular carcinoma (HepG2) cells with the MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] (Sigma-Aldrich) assay. Briefly, cells were cultivated in Dulbecco's modified Eagle's medium (DMEM) (Nutricell, Brazil) supplemented with 10% fetal bovine serum, penicillin (1 unit/ml), and streptomycin (1 unit/ml), in a humidified atmosphere of 5% CO2 at 37°C. HepG2 cells were seeded into 96-well plates at a density of 2 × 104 cells per well and were incubated with different concentrations of FucCS. After 48 h, a solution of 5 mg/ml MTT was added, the cells were incubated for 4 h, and the resulting formazan crystals were resuspended in 50 μl of isopropanol. Absorbance at 590 nm (A590) of the cells, with medium alone (control) or different concentrations of FucCS, was determined by spectrophotometry (Asys Expert Plus, United Kingdom). Toxicity was determined as percent inhibition, which was calculated according to the following formula: percent inhibition = [1 − (At/Ac)] × 100. At and Ac refer to the absorbance of experimentally tested samples (FucCS) and control samples (medium), respectively. FucCS toxicity was also determined in noninfected erythrocytes (niEs) incubated in the presence or absence (control) of FucCS. After 48 h of incubation at 37°C under conditions similar to those used for P. falciparum growth, red blood cell density (RBCD) was determined by counting the intact cells in a Neubauer chamber. The percent RBCD was calculated with the following formula: percent RBCD = [1 − (no. of niEs treated with FucCS/no. of niEs not treated)] × 100. Nontoxic samples were those in which no significant inhibition of HepG2 cell growth or RBCD, relative to controls, was observed.
Cultivation of laboratory strains and clinical isolates of P. falciparum-infected erythrocytes.
In this work, we used P. falciparum strains FCR3 (54, 55) and FCR3S1.2 (56). FCR3S1.2 is a highly rosetting and autoagglutinating parasite, kindly provided by Mats Wahlgren (Karolinska Institute, Stockholm, Sweden). The rosetting rate was kept at >75% by centrifugation on Ficoll-Paque (GE, Germany), as described previously (56). For some of the cytoadhesion assays, we used two fresh P. falciparum isolates (Pf38A and PfKA) that were recently harvested from infected patients in Acre State (western Amazon Basin of Brazil). All Pf-iE strains and isolates were cultured as described previously (57). Briefly, Pf-iEs were cultivated in fresh type O+ human erythrocytes (Blood Center, Universidade Estadual de Campinas) and suspended at a final hematocrit level of 4% in complete parasite medium (RPMI 1640 medium supplemented with l-glutamine, 25 mM HEPES, 2 g/liter glucose, and 10% homologous human plasma [pH 7.4]).
Selection of monophenotypic P. falciparum parasites.
The following cell types were used in this study: human lung endothelial cells (HLECs) (58, 59) adapted for culture from primary explants and Chinese hamster ovary (CHO) cells transfected with human ICAM-1 (CHO-ICAM) or CD36 (CHO-CD36) (60). HLECs, CHO cells, and FCR3 parasites were kindly donated by Jürg Gysin and Artur Scherf (Institute Pasteur, Paris, France). FCR3 parasites that adhered to CSA (Pf-iEsCSA) were selected by panning (5 rounds) on HLECs (61, 62) with the addition of soluble CSA (100 mg/ml; Sigma-Aldrich). Briefly, enriched P. falciparum FCR3 parasites were incubated over confluent HLEC monolayers in cytoadhesion medium (RPMI 1640 medium [pH 6.8]) for 1 h at 37°C. Cells were then washed extensively with cytoadhesion medium to remove nonadherent Pf-iEs. For specific elution of CSA-binders, soluble CSA (100 μg/ml; Sigma-Aldrich) was added and detached parasites were recultured. Monophenotypic Pf-iEs capable of binding to ICAM-1 (Pf-iEsICAM) or CD36 (Pf-iEsCD36) were obtained after panning (5 rounds) of mature-stage Pf-iEs on CHO-ICAM or CHO-CD36 cells, respectively (58, 63). To ensure selection of monophenotypic Pf-iEsCSA, static cytoadhesion assays were performed with HLECs that had been previously treated with chondroitinase-ABC (0.5 U/ml; Sigma-Aldrich) or by incubation with soluble CSA. High-level expression of var2csa-related transcripts in selected Pf-iEsCSA was confirmed by quantitative reverse transcription-PCR (qRT-PCR) using cDNA from ring-stage parasites as a template and oligonucleotides specific for the genes PFL0020c, PFD0995c/PFD1000c, and PFL0030w (var2csa) and the internal control targets PF07_0073 and PF14_0425, as described previously (64). The selection of Pf-iEsICAM and Pf-iEsCD36 was confirmed using anti-ICAM-1 (10 μg/ml, 84H10; Chemicon) and anti-CD36 (10 μg/ml, FA6-152; Santa Cruz Biotechnology) monoclonal antibodies, respectively.
Static cytoadhesion assays.
We assessed the ability of native or modified FucCS and other GAGs to inhibit the adhesion of Pf-iEsCSA, Pf-iEsICAM, Pf-iEsCD36, and P. falciparum isolates to human lung endothelial cells (HLECs) by performing static cytoadhesion assays, as described elsewhere (34, 61, 65, 66), with minor modifications. Briefly, HLECs (5 × 104 cells) were grown to confluence on 8-well culture slides (each well, 0.69 cm2; Becton, Dickinson). We then added 5 × 104 Voluven-enriched Pf-iEs per well in a total volume of 300 μl of cytoadhesion medium (RPMI 1640 medium [pH 6.8]), alone or in the presence of increasing concentrations of heparin (Calbiochem, China), CSA, native FucCS, or modified FucCS structures (deFucCS and CRFucCS). Culture slides were incubated for 1 h at 37°C and then extensively washed in cytoadhesion medium. Alternatively, we performed static cytoadhesion assays on placenta cryosections as described previously (54, 58, 65, 66), with minor modifications. Briefly, placental biopsy specimens from healthy pregnant women were collected immediately after delivery, snap-frozen in liquid nitrogen/n-hexane (Merck, Germany), and then stored frozen in Tissue-Tek (Thermo) before use. Serial placenta cryosections (5 to 7 mm) were cut with a cryotome and mounted on individual glass slides. Cryosections were washed and air-dried, and an area of approximately 1 cm2 was delimited with a Dako-Pen device. Cytoadhesion assays were performed as for endothelial cells; however, 1 × 105 Pf-iEsCSA were added to placenta cryosections. The percent inhibition, relative to control (medium) values, was determined by counting, with the aid of a microscope after Giemsa staining, the number of Pf-iEs per mm2 that adhered to an endothelial cell monolayer or a placenta cryosection. Adhesion assays were performed at least 2 or 3 times independently, and inhibitory values did not differ significantly.
Flow-based cytoadhesion assays.
The ability of FucCS to desequester Pf-iEsCSA was assessed through flow-based cytoadhesion assays, as described previously (54, 58). Briefly, HLECs (5 × 105) were cultured to confluence in single-well culture slides (8.6 cm2; Becton, Dickinson); then, 5 × 105 Pf-iEsCSA were added to the culture slides and were left for static cytoadherence. After 1 h of incubation at 37°C, the culture slides were mounted in a flow chamber system (Immunetics), and cytoadhesion medium (control) or FucCS (100 μg/ml) was circulated at a shear stress of 0.09, 0.36, or 1.44 Pa, for 10, 5, or 2.5 min, respectively. For the flow adhesion assay, enriched Pf-iEs (2 × 106) were subjected to the flow system for 10 min at a shear stress of 0.09 Pa. The culture slides were coupled in the flow chamber system, through which passed cytoadhesion medium (control) or FucCS (100 μg/ml) together with parasites. In both experiments, the remaining adherent Pf-iEsCSA per mm2 were counted in 30 randomly chosen fields, which were captured using a 5.0-megapixel digital camera (Moticam 2500; Motic) adapted to the inverted microscope.
Rosette disruption assay.
This assay was performed essentially as described previously (36). Briefly, different concentrations of FucCS, deFucCS, heparin, or CSA were added to 18-μl aliquots of a rosetting FCR3S1.2 culture in a 96-well microtiter plate, which was then incubated for 1 h at 37°C. FCR3S1.2 was also monitored over time (2, 10, 30, and 60 min) after treatment with 50 μg/ml of FucCS or heparin, to examine the kinetics of rosette disruption. A rosette was scored when a parasitized cell bound to at least two noninfected red blood cells. Rosetting was assessed after acridine orange (Sigma-Aldrich) staining. The percent rosetting, relative to control (medium) values, was expressed as follows: percent rosetting = (no. of rosette-forming late-stage Pf-iEs/total no. of late-stage Pf-iEs) × 100.
Growth inhibition assays.
The effects of FucCS, deFucCS, CRFucCS, and other GAGs on Pf-iE development were assessed by growth inhibition assays (GIAs) (67). Briefly, synchronized young-stage forms (<24 h) of P. falciparum cultures with 4% parasitemia and hematocrit levels were grown for 24 or 48 h at 37°C in a 96-well microtiter plate in the presence of increasing concentrations of GAGs. Parasite growth inhibition, expressed as a percentage of control (medium) values, was determined by counting Pf-iEs in at least 1,000 cells on Giemsa-stained smears, under a light microscope.
Merozoite invasion inhibition assay.
The merozoite invasion inhibition assay with native and modified FucCS, or other GAGs, was performed as described previously (37), with modifications. In short, synchronized late-stage forms (>24 h) of P. falciparum cultures were grown for 24 to 30 h at 37°C in a 96-well microtiter plate with increasing concentrations of GAGs, and parasite invasion inhibition was determined as described for GIAs.
Statistical analysis.
Statistical significance was determined using one-way analysis of variance (ANOVA) or Student's t test for parametric data. Kruskal-Wallis and post hoc tests or the Mann-Whitney U test was used for nonparametric data. All statistical analyses were performed using Prism version 5.02 (GraphPad Software), and values were considered significant at P < 0.05.
RESULTS
Effects of FucCS on P. falciparum cytoadherence and rosette disruption.
Based on the ability of various GAGs, including heparin, to prevent parasite cytoadhesion, we assessed the cytoadherence-inhibiting effect of FucCS using monophenotypic Pf-iEs with HLECs and human placenta cryosections. To evaluate FucCS toxicity, several concentrations of the compound were incubated with HepG2 cells and noninfected erythrocytes (niEs). FucCS significantly impaired hepatic cell growth only at a high concentration of 10,000 μg/ml, and no effect on niEs was observed at any of the tested doses (see Fig. S1A and B in the supplemental material). Thus, we analyzed the ability of FucCS to inhibit P. falciparum cytoadherence to endothelial cells (Fig. 2). Indeed, FucCS abolished cytoadherence in a dose-dependent manner similar to that of heparin. The adhesion of parasites that bind to specific host receptors, such as CSA (Pf-iEsCSA), ICAM-1 (Pf-iEsICAM), and CD36 (Pf-iEsCD36), was inhibited by 99.1, 64.6, and 71.2%, respectively (Fig. 2A to C). Moreover, at a concentration as low as 1 μg/ml, FucCS was more efficient than heparin (Fig. 2A). As natural infections of P. falciparum often involve multiphenotypic parasites (e.g., those binding to different receptors) (68, 69), we examined whether FucCS could block the adhesion of fresh P. falciparum isolates. According to Fig. 2D and E, FucCS also inhibited adhesion of clinical isolates in a dose-dependent manner and was significantly more effective than heparin at several tested concentrations.
FIG 2.

Effects of FucCS (native or modified) on the cytoadhesion of P. falciparum (laboratory strains or clinical isolates) to HLECs. P. falciparum monophenotypic laboratory strains Pf-iEsCSA (A), Pf-iEsICAM (B), and Pf-iEsCD36 (C) and clinical Amazonian isolates PfKA (D) and Pf38A (E) were incubated for 1 h at 37°C on 8-well culture slides containing HLECs, in the presence or absence (control) of different concentrations of FucCS or heparin, or Pf-iEsCSA were incubated for 1 h at 37°C on HLECs in the presence of 100 μg/ml of native FucCS or the modified forms (deFucCS or CRFucCS) or culture medium alone (control) (F). Slides were extensively washed, and bound Pf-iEs were counted after Giemsa staining. Inhibition was determined as a percentage, relative to control values, and results are expressed as the means ± standard deviations (SD) of four wells from 2 independent assays. *, P < 0.05; ***, P < 0.001 (ANOVA).
Next, we investigated the major structural motifs in FucCS involved in the inhibition of parasite adhesion, using two chemically modified polysaccharides. As observed in Fig. 2F, both deFucCS and CRFucCS were significantly less effective than native FucCS in inhibiting Pf-iEsCSA adhesion. CRFucCS blocked parasite adhesion at higher levels than did deFucCS (P < 0.001), and similar effects were noted for Pf-iEsICAM and Pf-iEsCD36 (see Fig. S2A and B in the supplemental material).
Given that CSA is a receptor associated with MiP and that FucCS abolished adhesion of Pf-iEsCSA, we evaluated the capacity of FucCS to inhibit the binding of this parasite to placenta cryosections. As shown in Fig. 3, 100 μg/ml of FucCS abrogated Pf-iEsCSA adherence to the human placenta. We also evaluated the effects of FucCS on parasite adhesion under flow conditions. As shown in Fig. 4A, virtually no Pf-iEs adhered to HLECs under a constant flow pressure of 0.09 Pa in a solution of 100 μg/ml FucCS. Importantly, FucCS, in comparison with the control (medium), removed a significant amount of CSA-adherent parasites, even under conditions of low shear stress (0.09 Pa) (Fig. 4B).
FIG 3.
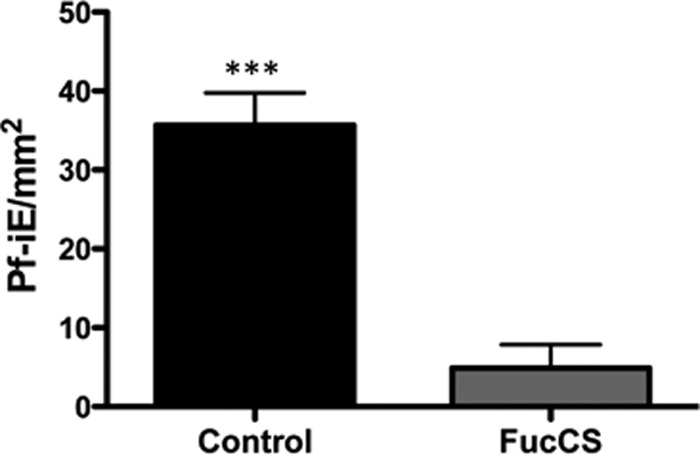
FucCS inhibition of adhesion of CSA-bound parasites to placenta. Cultured P. falciparum parasites selected for CSA binding (Pf-iEsCSA) were incubated at 37°C with placenta cryosections, with FucCS (100 μg/ml) or cytoadhesion medium (control). After 1 h, placental tissue samples were extensively washed, and adhered parasites were Giemsa stained and then visualized and counted using a Nikon microscope at ×100 magnification. The results are expressed as the mean ± SD of the number of bound Pf-iEsCSA/mm2 counted in at least 30 randomly selected fields after Giemsa staining. ***, P < 0.001 (Mann-Whitney test).
FIG 4.
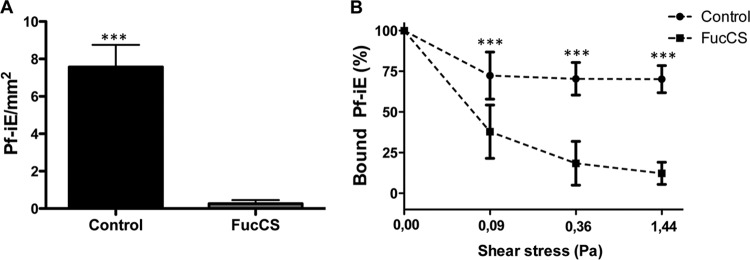
FucCS inhibition, under flow conditions, of Pf-iECSA cytoadhesion and unbound adhered parasites on HLECs. (A) Enriched Pf-iEsCSA, in the presence of FucCS (100 μg/ml) or cytoadhesion medium only (control), were circulated for 10 min at 0.09 Pa on culture slides with HLECs coupled in the flow chamber system. The results are expressed as the number of bound Pf-iEsCSA per square millimeter. (B) Alternatively, cytoadhesion medium (control) or FucCS (100 μg/ml) flowed through single-well microslides containing HLECs with adhered Pf-iEsCSA, under increasing shear stress conditions (0.09, 0.36, and 1.44 Pa). Data are expressed as a percentage, relative to the adhesion at 0.00 Pa (control). In both situations, the remaining bound Pf-iEsCSA were counted in 30 randomly selected fields. ***;, P < 0.001 (Mann-Whitney test).
Having demonstrated the ability of FucCS to prevent parasite binding under static and flow conditions, we investigated the disruptive effects of FucCS on P. falciparum rosettes (FCR3S1.2 strain). As seen in Fig. 5A, FucCS disrupted rosettes in a dose-dependent manner and virtually abolished rosette formation, even at low concentrations such as 10 μg/ml. As observed in cytoadhesion assays, deFucCS proved to be less efficient than the native compound in disrupting rosettes. We then evaluated how quickly FucCS could disrupt rosettes by treating FCR3S1.2 cultures with FucCS or heparin at different time points. As shown in Fig. 5B, FucCS was able to disrupt rosettes at high levels within only 2 min after its addition, similar to heparin.
FIG 5.
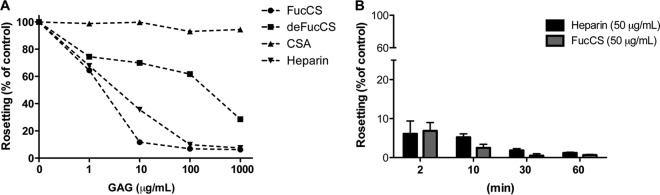
FucCS disruption of P. falciparum rosettes. Rosetting parasite cultures (FCR3S1.2) were incubated for 1 h with or without (control) FucCS, deFucCS, heparin, or CSA at various concentrations (A) or were treated with FucCS or heparin (both at 50 μg/ml) at different time points (2, 10, 30, and 60 min) (B). The percentage of rosetting, relative to control, was expressed as (no. of rosette-forming late-stage Pf-iEs)/(total no. of late-stage Pf-iEs) × 100. Data are from one representative experiment from three independent assays.
Effects of FucCS on P. falciparum growth and erythrocyte invasion.
As previous studies have reported, GAGs, including heparin, have major effects on parasite invasion (37, 40, 70, 71); therefore, we investigated the action of FucCS on parasite development by GIA. P. falciparum blood-stage parasites were incubated with native or modified FucCS, as well as heparin or CSA, for 48 h. As shown in Fig. 6A, similar to heparin, FucCS exhibited dose-dependent inhibitory effects and completely abolished parasite development at concentrations of 100 and 1,000 μg/ml. Importantly, as noted in the cytoadhesion assays, CRFucCS significantly hampered parasite development, whereas deFucCS did not. Additionally, CSA inhibition was observed only at the highest tested dose (1,000 μg/ml). Then, to determine whether FucCS could act on a specific parasite blood stage, we incubated synchronous young-stage (<24 h postinvasion) forms of P. falciparum with native or modified FucCS, as well as heparin or CSA. As seen in Fig. 6B, neither FucCS (native or modified) nor any of the tested GAGs significantly inhibited ring-stage maturation.
FIG 6.

FucCS inhibition of P. falciparum growth (GIA) and merozoite invasion. Ring-synchronized P. falciparum-infected erythrocytes (A and B) or late-stage forms (trophozoites) (C) cultured at 4% parasitemia and hematocrit levels were incubated at 37°C for 48 h (A) or 24 h (B and C) with increasing concentrations of FucCS, heparin, CSA, or defucosylated (100 μg/ml) or carboxyl-reduced (100 μg/ml) modified FucCS. As a control, parasites were grown in RPMI 1640 medium. The percentage of growth inhibition was calculated relative to control values, and results are expressed as the means ± SD of triplicates. *, P < 0.05; **, P < 0.01 (Kruskal-Wallis test).
Assuming that FucCS affects parasite development despite its inability to stop ring-stage maturation, we analyzed the role of FucCS in the invasion of noninfected erythrocytes (niEs) by P. falciparum merozoites. As shown in Fig. 6C, CRFucCS but not deFucCS significantly inhibited parasite invasion at 100 μg/ml, and FucCS or heparin hampered merozoite invasion in a dose-dependent manner, at levels similar to those observed in GIAs, after 48 h of incubation.
DISCUSSION
Here we show that FucCS exhibited marked inhibition of the cytoadhesion of cultured Pf-iEs and clinical isolates to endothelial cells and placenta cryosections under static and flow conditions. FucCS was also highly effective in disrupting rosettes and interfering with parasite invasion. These results suggest that FucCS may prove to be a useful adjunct therapy for severe malaria.
The observed effects of FucCS on P. falciparum cytoadhesion and invasion were very similar to those of heparin in most assays. In fact, heparin displays potent inhibitory effects on Pf-iE cytoadherence and rosette formation in vitro (28, 36, 42, 72–74) and remains the only sulfated glycoconjugate used to treat severe human malaria (75–77). However, despite some success as an adjunct treatment for complications of malaria (77–79), the use of heparin was abandoned due to severe bleeding and the deaths of some children under treatment (47).
When we analyzed the effects of modified compounds derived from native FucCS, we noted marked differences between these compounds (Fig. 2F; also see Fig. S2 in the supplemental material). Although both modified FucCS structures exhibited inhibitory cytoadhesion capacity that was significantly less than that of the native compound, deFucCS was nearly 3-fold less efficient than CRFucCS. These findings demonstrate that the presence of sulfated fucose branches in the native compound is critical for the inhibition of Pf-iEs, and they corroborate previous observations that removal of these branches abolishes the anticoagulant and antithrombotic effects of FucCS (80). The presence of these branches was essential for preserving the inhibitory effects of FucCS in interactions mediated by P- and L-selectin (52), thus indicating that these branches play a key role in supporting the biological effects of FucCS.
Cytoadhesion of P. falciparum to CSA, and possibly other placental receptors, triggers host inflammatory processes, which in turn affect placental function and contribute to the poor clinical outcomes of MiP (18, 19). Based on this fact, CSA has previously been considered for the adjunct treatment of malaria complications during pregnancy (41), due to its inhibitory effects on the in vitro and in vivo cytoadherence of Pf-iEs (41). In this context, we showed that FucCS was very efficient in inhibiting the adhesion of Pf-iEsCSA to HLECs and virtually abolished adhesion to human placenta. Additionally, FucCS was effective in preventing adhesion and removing Pf-iEsCSA from HLECs under flow conditions.
Apart from CSA (42, 72), rosettes can be disrupted by other sulfated glycoconjugates, such as heparin (36, 38, 42), dextran sulfate, and fucoidan (38, 72). Here we have shown that FucCS has effects that are similar to those of heparin, including the speed of action for rosette disruption (Fig. 5). This action was dependent on the sulfated fucose branches in the native structure of FucCS, as deFucCS proved to be less effective than the native compound.
Studies have shown that heparin and other sulfated polysaccharides (e.g., pentosan polysulfate, dextran sulfate, Curdlan sulfate, and fucoidan) inhibit the progression of P. falciparum through its blood stages (40, 43, 71, 81, 82). Moreover, it has been demonstrated that heparin and other GAGs, but not CSA (40, 42), efficiently inhibit the invasion of erythrocytes by the merozoites of different strains of P. falciparum (37, 40, 42, 43). Corroborating these findings, we showed that FucCS inhibited continuous cultivation of P. falciparum in vitro (Fig. 6A), most likely by interfering with merozoite invasion (Fig. 6C) rather than by blocking parasite development from the young- to late-stage forms, as assessed in cultures of 24 and 48 h (Fig. 6A and B). In contrast, CSA did not show any significant inhibitory effects on merozoite invasion (Fig. 6C) and parasite development (Fig. 6A and B) except at the highest dose tested (1,000 mg/ml), which was most likely due to a nonspecific effect of an excess of the compound in the medium. These results agree with previous studies in which CSA had no significant inhibitory effects on reinvasion, especially at lower doses, with only marginal effects at high doses (40, 42).
Some studies have attempted to explain the mechanisms involved in the inhibition of invasion by sulfated polysaccharides. Clark and colleagues showed that only densely sulfated polysaccharides (heparin, dextran sulfate, and fucoidan) were effective in inhibiting parasite invasion (43), suggesting that inhibition was the result of not only nonspecific ionic interactions but also a particular conformation of anions present in the compounds. Boyle and colleagues demonstrated that the level and pattern of sulfation are crucial for the inhibitory activity of heparin and similar compounds (83).
The effect of FucCS seems to be similar to that of heparin, which inhibits parasite invasion, adhesion, and rosetting by binding to regions of PfEMP-1 and merozoite surface protein-1 (MSP-1) (83–85). Based on these findings, we hypothesized that FucCS could block interactions of adhesion and invasion proteins with host receptors that are glycoproteins expressed constitutively in erythrocytes, most likely by binding to conserved regions of these proteins, such as region RII (86). Docking analyses are under way to examine this hypothesis.
In addition to the effects described in this study, FucCS does not require fractionation and/or chemical modification after purification (87) and is not derived from mammals (88), in contrast to heparin and CSA, thus reducing the risk of contamination by pathogens. Moreover, FucCS occurs at high concentrations in sea cucumber and can be isolated with relatively high yields (approximately 1% of the dry weight) (87). Also, it has weaker anticoagulant and antithrombotic effects than heparin (48, 89) and acts, in vitro and in vivo, at concentrations lower than those required to trigger its anticoagulant effects (52). FucCS can also be administered orally, which is another advantage in comparison with other sulfated polysaccharides (90). Importantly, no toxic or cumulative effects in tissue were observed after administration of FucCS to animals at a daily dose of 50 mg/kg of body weight for 30 days (M. S. G. Pavão and E. O. Kozlowski, unpublished data).
In conclusion, the observed similarities in the inhibitory effects of FucCS and heparin on Pf-iE cytoadhesion, rosette formation, and blockage of parasite invasion, as well as the unique sulfated composition of FucCS, which confers advantages over other compounds that have already been tested, raise hopes for the use of FucCS in human trials as an adjunct therapy for the treatment of several complications that characterize severe malaria.
Supplementary Material
ACKNOWLEDGMENTS
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). M.F.B., L.A., S.C.P.L., and B.C.C. were sponsored by FAPESP fellowships. Y.C.B. was supported by a CNPq fellowship. F.T.M.C., C.C.W., M.U.F., P.A.S.M., G.W., and M.S.G.P. are CNPq research fellows. C.R.F.M. was supported by FAPESP and CNPq.
Many thanks go to Maria José Menezes (Department of Parasitology, Instituto de Ciências Biomédicas, Universidade de São Paulo, San Paulo, Brazil) for technical support.
Footnotes
Published ahead of print 6 January 2014
Supplemental material for this article may be found at http://dx.doi.org/10.1128/AAC.00686-13.
REFERENCES
- 1.Snow RW, Guerra CA, Noor AM, Myint HY, Hay SI. 2005. The global distribution of clinical episodes of Plasmodium falciparum malaria. Nature 434:214–217. 10.1038/nature03342 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dellicour S, Tatem AJ, Guerra CA, Snow RW, ter Kuile FO. 2010. Quantifying the number of pregnancies at risk of malaria in 2007: a demographic study. PLoS Med. 7:e1000221. 10.1371/journal.pmed.1000221 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Steketee RW, Nahlen BL, Parise ME, Menendez C. 2001. The burden of malaria in pregnancy in malaria-endemic areas. Am. J. Trop. Med. Hyg. 64:28–35 [DOI] [PubMed] [Google Scholar]
- 4.World Health Organization, Communicable Diseases Cluster. 2000. Severe falciparum malaria. Trans. R. Soc. Trop. Med. Hyg. 94(Suppl 1):S1–S90 [PubMed] [Google Scholar]
- 5.Dondorp A, Nosten F, Stepniewska K, Day N, White N. 2005. Artesunate versus quinine for treatment of severe falciparum malaria: a randomised trial. Lancet 366:717–725. 10.1016/S0140-6736(05)67176-0 [DOI] [PubMed] [Google Scholar]
- 6.Menendez C. 2006. Malaria during pregnancy. Curr. Mol. Med. 6:269–273. 10.2174/156652406776055186 [DOI] [PubMed] [Google Scholar]
- 7.Clark IA, Rockett KA. 1994. The cytokine theory of human cerebral malaria. Parasitol. Today 10:410–412. 10.1016/0169-4758(94)90237-2 [DOI] [PubMed] [Google Scholar]
- 8.Clark IA, Rockett KA, Cowden WB. 1991. Role of TNF in cerebral malaria. Lancet 337:302–303 [DOI] [PubMed] [Google Scholar]
- 9.Fried M, Muga RO, Misore AO, Duffy PE. 1998. Malaria elicits type 1 cytokines in the human placenta: IFN-γ and TNF-α associated with pregnancy outcomes. J. Immunol. 160:2523–2530 [PubMed] [Google Scholar]
- 10.Suguitan AL, Jr, Leke RG, Fouda G, Zhou A, Thuita L, Metenou S, Fogako J, Megnekou R, Taylor DW. 2003. Changes in the levels of chemokines and cytokines in the placentas of women with Plasmodium falciparum malaria. J. Infect. Dis. 188:1074–1082. 10.1086/378500 [DOI] [PubMed] [Google Scholar]
- 11.Fievet N, Moussa M, Tami G, Maubert B, Cot M, Deloron P, Chaouat G. 2001. Plasmodium falciparum induces a Th1/Th2 disequilibrium, favoring the Th1-type pathway, in the human placenta. J. Infect. Dis. 183:1530–1534. 10.1086/320201 [DOI] [PubMed] [Google Scholar]
- 12.Taylor TE, Fu WJ, Carr RA, Whitten RO, Mueller JS, Fosiko NG, Lewallen S, Liomba NG, Molyneux ME. 2004. Differentiating the pathologies of cerebral malaria by postmortem parasite counts. Nat. Med. 10:143–145. 10.1038/nm986 [DOI] [PubMed] [Google Scholar]
- 13.MacPherson GG, Warrell MJ, White NJ, Looareesuwan S, Warrell DA. 1985. Human cerebral malaria: a quantitative ultrastructural analysis of parasitized erythrocyte sequestration. Am. J. Pathol. 119:385–401 [PMC free article] [PubMed] [Google Scholar]
- 14.Beeson JG, Amin N, Kanjala M, Rogerson SJ. 2002. Selective accumulation of mature asexual stages of Plasmodium falciparum-infected erythrocytes in the placenta. Infect. Immun. 70:5412–5415. 10.1128/IAI.70.10.5412-5415.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Fried M, Duffy PE. 1996. Adherence of Plasmodium falciparum to chondroitin sulfate A in the human placenta. Science 272:1502–1504. 10.1126/science.272.5267.1502 [DOI] [PubMed] [Google Scholar]
- 16.Bray RS, Sinden RE. 1979. The sequestration of Plasmodium falciparum infected erythrocytes in the placenta. Trans. R. Soc. Trop. Med. Hyg. 73:716–719. 10.1016/0035-9203(79)90028-2 [DOI] [PubMed] [Google Scholar]
- 17.Hanson J, Lam SW, Mahanta KC, Pattnaik R, Alam S, Mohanty S, Hasan MU, Hossain A, Charunwatthana P, Chotivanich K, Maude RJ, Kingston H, Day NP, Mishra S, White NJ, Dondorp AM. 2012. Relative contributions of macrovascular and microvascular dysfunction to disease severity in falciparum malaria. J. Infect. Dis. 206:571–579. 10.1093/infdis/jis400 [DOI] [PubMed] [Google Scholar]
- 18.Beeson JG, Reeder JC, Rogerson SJ, Brown GV. 2001. Parasite adhesion and immune evasion in placental malaria. Trends Parasitol. 17:331–337. 10.1016/S1471-4922(01)01917-1 [DOI] [PubMed] [Google Scholar]
- 19.Duffy PE, Fried M. 2005. Malaria in the pregnant woman. Curr. Top. Microbiol. Immunol. 295:169–200 [DOI] [PubMed] [Google Scholar]
- 20.Miller LH. 1969. Distribution of mature trophozoites and schizonts of Plasmodium falciparum in the organs of Aotus trivirgatus, the night monkey. Am. J. Trop. Med. Hyg. 18:860–865 [DOI] [PubMed] [Google Scholar]
- 21.Luse SA, Miller LH. 1971. Plasmodium falciparum malaria: ultrastructure of parasitized erythrocytes in cardiac vessels. Am. J. Trop. Med. Hyg. 20:655–660 [PubMed] [Google Scholar]
- 22.Pouvelle B, Buffet PA, Lepolard C, Scherf A, Gysin J. 2000. Cytoadhesion of Plasmodium falciparum ring-stage-infected erythrocytes. Nat. Med. 6:1264–1268. 10.1038/81374 [DOI] [PubMed] [Google Scholar]
- 23.Doumbo OK, Thera MA, Kone AK, Raza A, Tempest LJ, Lyke KE, Plowe CV, Rowe JA. 2009. High levels of Plasmodium falciparum rosetting in all clinical forms of severe malaria in African children. Am. J. Trop. Med. Hyg. 81:987–993. 10.4269/ajtmh.2009.09-0406 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Udomsangpetch R, Wahlin B, Carlson J, Berzins K, Torii M, Aikawa M, Perlmann P, Wahlgren M. 1989. Plasmodium falciparum-infected erythrocytes form spontaneous erythrocyte rosettes. J. Exp. Med. 169:1835–1840. 10.1084/jem.169.5.1835 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Handunnetti SM, David PH, Perera KL, Mendis KN. 1989. Uninfected erythrocytes form “rosettes” around Plasmodium falciparum infected erythrocytes. Am. J. Trop. Med. Hyg. 40:115–118 [DOI] [PubMed] [Google Scholar]
- 26.Carlson J, Helmby H, Hill AV, Brewster D, Greenwood BM, Wahlgren M. 1990. Human cerebral malaria: association with erythrocyte rosetting and lack of anti-rosetting antibodies. Lancet 336:1457–1460. 10.1016/0140-6736(90)93174-N [DOI] [PubMed] [Google Scholar]
- 27.Treutiger CJ, Hedlund I, Helmby H, Carlson J, Jepson A, Twumasi P, Kwiatkowski D, Greenwood BM, Wahlgren M. 1992. Rosette formation in Plasmodium falciparum isolates and anti-rosette activity of sera from Gambians with cerebral or uncomplicated malaria. Am. J. Trop. Med. Hyg. 46:503–510 [DOI] [PubMed] [Google Scholar]
- 28.Rowe A, Obeiro J, Newbold CI, Marsh K. 1995. Plasmodium falciparum rosetting is associated with malaria severity in Kenya. Infect. Immun. 63:2323–2326 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.al-Yaman F, Genton B, Mokela D, Raiko A, Kati S, Rogerson S, Reeder J, Alpers M. 1995. Human cerebral malaria: lack of significant association between erythrocyte rosetting and disease severity. Trans. R. Soc. Trop. Med. Hyg. 89:55–58. 10.1016/0035-9203(95)90658-4 [DOI] [PubMed] [Google Scholar]
- 30.Turner L, Lavstsen T, Berger SS, Wang CW, Petersen JEV, Avril M, Brazier AJ, Freeth J, Jespersen JS, Nielsen MA, Magistrado P, Lusingu J, Smith JD, Higgins MK, Theander TG. 2013. Severe malaria is associated with parasite binding to endothelial protein C receptor. Nature 498:502–505. 10.1038/nature12216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ockenhouse CF, Tandon NN, Magowan C, Jamieson GA, Chulay JD. 1989. Identification of a platelet membrane glycoprotein as a falciparum malaria sequestration receptor. Science 243:1469–1471. 10.1126/science.2467377 [DOI] [PubMed] [Google Scholar]
- 32.Berendt AR, Simmons DL, Tansey J, Newbold CI, Marsh K. 1989. Intercellular adhesion molecule-1 is an endothelial cell adhesion receptor for Plasmodium falciparum. Nature 341:57–59. 10.1038/341057a0 [DOI] [PubMed] [Google Scholar]
- 33.Rogerson SJ, Chaiyaroj SC, Ng K, Reeder JC, Brown GV. 1995. Chondroitin sulfate A is a cell surface receptor for Plasmodium falciparum-infected erythrocytes. J. Exp. Med. 182:15–20. 10.1084/jem.182.1.15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Robert C, Pouvelle B, Meyer P, Muanza K, Fujioka H, Aikawa M, Scherf A, Gysin J. 1995. Chondroitin-4-sulphate (proteoglycan), a receptor for Plasmodium falciparum-infected erythrocyte adherence on brain microvascular endothelial cells. Res. Immunol. 146:383–393. 10.1016/0923-2494(96)81042-X [DOI] [PubMed] [Google Scholar]
- 35.Fried M, Domingo GJ, Gowda CD, Mutabingwa TK, Duffy PE. 2006. Plasmodium falciparum: chondroitin sulfate A is the major receptor for adhesion of parasitized erythrocytes in the placenta. Exp. Parasitol. 113:36–42. 10.1016/j.exppara.2005.12.003 [DOI] [PubMed] [Google Scholar]
- 36.Carlson J, Wahlgren M. 1992. Plasmodium falciparum erythrocyte rosetting is mediated by promiscuous lectin-like interactions. J. Exp. Med. 176:1311–1317. 10.1084/jem.176.5.1311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kulane A, Ekre HP, Perlmann P, Rombo L, Wahlgren M, Wahlin B. 1992. Effect of different fractions of heparin on Plasmodium falciparum merozoite invasion of red blood cells in vitro. Am. J. Trop. Med. Hyg. 46:589–594 [DOI] [PubMed] [Google Scholar]
- 38.Rowe A, Berendt AR, Marsh K, Newbold CI. 1994. Plasmodium falciparum: a family of sulphated glycoconjugates disrupts erythrocyte rosettes. Exp. Parasitol. 79:506–516. 10.1006/expr.1994.1111 [DOI] [PubMed] [Google Scholar]
- 39.Rogerson SJ, Reeder JC, al-Yaman F, Brown GV. 1994. Sulfated glycoconjugates as disrupters of Plasmodium falciparum erythrocyte rosettes. Am. J. Trop. Med. Hyg. 51:198–203 [DOI] [PubMed] [Google Scholar]
- 40.Xiao L, Yang C, Patterson PS, Udhayakumar V, Lal AA. 1996. Sulfated polyanions inhibit invasion of erythrocytes by plasmodial merozoites and cytoadherence of endothelial cells to parasitized erythrocytes. Infect. Immun. 64:1373–1378 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pouvelle B, Meyer P, Robert C, Bardel L, Gysin J. 1997. Chondroitin-4-sulfate impairs in vitro and in vivo cytoadherence of Plasmodium falciparum infected erythrocytes. Mol. Med. 3:508–518 [PMC free article] [PubMed] [Google Scholar]
- 42.Vogt AM, Pettersson F, Moll K, Jonsson C, Normark J, Ribacke U, Egwang TG, Ekre HP, Spillmann D, Chen Q, Wahlgren M. 2006. Release of sequestered malaria parasites upon injection of a glycosaminoglycan. PLoS Pathog. 2:e100. 10.1371/journal.ppat.0020100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Clark DL, Su S, Davidson EA. 1997. Saccharide anions as inhibitors of the malaria parasite. Glycoconj. J. 14:473–479. 10.1023/A:1018551518610 [DOI] [PubMed] [Google Scholar]
- 44.Andrews KT, Klatt N, Adams Y, Mischnick P, Schwartz-Albiez R. 2005. Inhibition of chondroitin-4-sulfate-specific adhesion of Plasmodium falciparum-infected erythrocytes by sulfated polysaccharides. Infect. Immun. 73:4288–4294. 10.1128/IAI.73.7.4288-4294.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kyriacou HM, Steen KE, Raza A, Arman M, Warimwe G, Bull Havlik PCI, Rowe JA. 2007. In vitro inhibition of Plasmodium falciparum rosette formation by Curdlan sulfate. Antimicrob. Agents Chemother. 51:1321–1326. 10.1128/AAC.01216-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.John CC, Kutamba E, Mugarura K, Opoka RO. 2010. Adjunctive therapy for cerebral malaria and other severe forms of Plasmodium falciparum malaria. Expert Rev. Anti Infect. Ther. 8:997–1008. 10.1586/eri.10.90 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.World Health Organization, Malaria Action Programme. 1986. Severe and complicated malaria. Trans. R. Soc. Trop. Med. Hyg. 80(Suppl):3–50 [PubMed] [Google Scholar]
- 48.Mourao PA, Pereira MS, Pavao MS, Mulloy B, Tollefsen DM, Mowinckel MC, Abildgaard U. 1996. Structure and anticoagulant activity of a fucosylated chondroitin sulfate from echinoderm: sulfated fucose branches on the polysaccharide account for its high anticoagulant action. J. Biol. Chem. 271:23973–23984 [DOI] [PubMed] [Google Scholar]
- 49.Vieira RP, Mourao PA. 1988. Occurrence of a unique fucose-branched chondroitin sulfate in the body wall of a sea cucumber. J. Biol. Chem. 263:18176–18183 [PubMed] [Google Scholar]
- 50.Vieira RP, Mulloy B, Mourao PA. 1991. Structure of a fucose-branched chondroitin sulfate from sea cucumber: evidence for the presence of 3-O-sulfo-β-d-glucuronosyl residues. J. Biol. Chem. 266:13530–13536 [PubMed] [Google Scholar]
- 51.Mourao PA, Giumaraes B, Mulloy B, Thomas S, Gray E. 1998. Antithrombotic activity of a fucosylated chondroitin sulphate from echinoderm: sulphated fucose branches on the polysaccharide account for its antithrombotic action. Br. J. Haematol. 101:647–652. 10.1046/j.1365-2141.1998.00769.x [DOI] [PubMed] [Google Scholar]
- 52.Borsig L, Wang L, Cavalcante MC, Cardilo-Reis L, Ferreira PL, Mourao PA, Esko JD, Pavao MS. 2007. Selectin blocking activity of a fucosylated chondroitin sulfate glycosaminoglycan from sea cucumber: effect on tumor metastasis and neutrophil recruitment. J. Biol. Chem. 282:14984–14991. 10.1074/jbc.M610560200 [DOI] [PubMed] [Google Scholar]
- 53.Taylor RL, Shively JE, Conrad HE. 1976. Stoichiometric reduction of uronic acid carboxyl groups in polysaccharides. Methods Carbohydr. Chem. 7:149–151 [Google Scholar]
- 54.Avril M, Traore B, Costa FT, Lepolard C, Gysin J. 2004. Placenta cryosections for study of the adhesion of Plasmodium falciparum-infected erythrocytes to chondroitin sulfate A in flow conditions. Microbes Infect. 6:249–255. 10.1016/j.micinf.2003.12.007 [DOI] [PubMed] [Google Scholar]
- 55.Lekana Douki JB, Traore B, Costa FT, Fusai T, Pouvelle B, Sterkers Y, Scherf A, Gysin J. 2002. Sequestration of Plasmodium falciparum-infected erythrocytes to chondroitin sulfate A, a receptor for maternal malaria: monoclonal antibodies against the native parasite ligand reveal pan-reactive epitopes in placental isolates. Blood 100:1478–1483. 10.1182/blood-2002-01-0315 [DOI] [PubMed] [Google Scholar]
- 56.Fernandez V, Treutiger CJ, Nash GB, Wahlgren M. 1998. Multiple adhesive phenotypes linked to rosetting binding of erythrocytes in Plasmodium falciparum malaria. Infect. Immun. 66:2969–2975 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Moll K, Ljungström I, Perlmann P, Scherf A, Wahlgren M (ed) 2008. Methods in malaria research, 5th ed. MR4/ATCC, Manassas, VA: http://www.mr4.org/Publications/MethodsinMalariaResearch/tabid/333/Default.aspx [Google Scholar]
- 58.Carvalho BO, Lopes SC, Nogueira PA, Orlandi PP, Bargieri DY, Blanco YC, Mamoni R, Leite JA, Rodrigues MM, Soares IS, Oliveira TR, Wunderlich G, Lacerda MV, del Portillo HA, Araujo MO, Russell B, Suwanarusk R, Snounou G, Renia L, Costa FT. 2010. On the cytoadhesion of Plasmodium vivax-infected erythrocytes. J. Infect. Dis. 202:638–647. 10.1086/654815 [DOI] [PubMed] [Google Scholar]
- 59.Muanza K, Gay F, Behr C, Scherf A. 1996. Primary culture of human lung microvessel endothelial cells: a useful in vitro model for studying Plasmodium falciparum-infected erythrocyte cytoadherence. Res. Immunol. 147:149–163. 10.1016/0923-2494(96)83167-1 [DOI] [PubMed] [Google Scholar]
- 60.Hasler T, Albrecht GR, Van Schravendijk MR, Aguiar JC, Morehead KE, Pasloske BL, Ma C, Barnwell JW, Greenwood B, Howard RJ. 1993. An improved microassay for Plasmodium falciparum cytoadherence using stable transformants of Chinese hamster ovary cells expressing CD36 or intercellular adhesion molecule-1. Am. J. Trop. Med. Hyg. 48:332–347 [DOI] [PubMed] [Google Scholar]
- 61.Gay F, Robert C, Pouvelle B, Peyrol S, Scherf A, Gysin J. 1995. Isolation and characterization of brain microvascular endothelial cells from Saimiri monkeys: an in vitro model for sequestration of Plasmodium falciparum-infected erythrocytes. J. Immunol. Methods 184:15–28 [DOI] [PubMed] [Google Scholar]
- 62.Pouvelle B, Fusai T, Gysin J. 1998. Plasmodium falciparum and chondroitin-4-sulfate: the new key couple in sequestration. Med. Trop. (Mars) 58:187–198 (In French.) [PubMed] [Google Scholar]
- 63.Noviyanti R, Brown GV, Wickham ME, Duffy MF, Cowman AF, Reeder JC. 2001. Multiple var gene transcripts are expressed in Plasmodium falciparum infected erythrocytes selected for adhesion. Mol. Biochem. Parasitol. 114:227–237. 10.1016/S0166-6851(01)00266-3 [DOI] [PubMed] [Google Scholar]
- 64.Salanti A, Staalsoe T, Lavstsen T, Jensen AT, Sowa MP, Arnot DE, Hviid L, Theander TG. 2003. Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol. Microbiol. 49:179–191. 10.1046/j.1365-2958.2003.03570.x [DOI] [PubMed] [Google Scholar]
- 65.Costa FT, Fusai T, Parzy D, Sterkers Y, Torrentino M, Douki JB, Traore B, Petres S, Scherf A, Gysin J. 2003. Immunization with recombinant duffy binding-like-γ3 induces pan-reactive and adhesion-blocking antibodies against placental chondroitin sulfate A-binding Plasmodium falciparum parasites. J. Infect. Dis. 188:153–164. 10.1086/375800 [DOI] [PubMed] [Google Scholar]
- 66.Pouvelle B, Traore B, Nogueira PA, Pradines B, LePolard C, Gysin J. 2003. Modeling of Plasmodium falciparum-infected erythrocyte cytoadhesion in microvascular conditions: chondroitin-4-sulfate binding, a competitive phenotype. J. Infect. Dis. 187:292–302. 10.1086/346050 [DOI] [PubMed] [Google Scholar]
- 67.Rieckmann KH, Campbell GH, Sax LJ, Mrema JE. 1978. Drug sensitivity of Plasmodium falciparum: an in-vitro microtechnique. Lancet 1(8054):22–23 [DOI] [PubMed] [Google Scholar]
- 68.Newbold C, Warn P, Black G, Berendt A, Craig A, Snow B, Msobo M, Peshu N, Marsh K. 1997. Receptor-specific adhesion and clinical disease in Plasmodium falciparum. Am. J. Trop. Med. Hyg. 57:389–398 [DOI] [PubMed] [Google Scholar]
- 69.Rogerson SJ, Tembenu R, Dobano C, Plitt S, Taylor TE, Molyneux ME. 1999. Cytoadherence characteristics of Plasmodium falciparum-infected erythrocytes from Malawian children with severe and uncomplicated malaria. Am. J. Trop. Med. Hyg. 61:467–472 [DOI] [PubMed] [Google Scholar]
- 70.Sivaraman CA, Rai Chowdhuri AN. 1983. Effect of heparin sodium on in vitro development of Plasmodium falciparum. Indian J. Exp. Biol. 21:247–250 [PubMed] [Google Scholar]
- 71.Butcher GA, Parish CR, Cowden WB. 1988. Inhibition of growth in vitro of Plasmodium falciparum by complex polysaccharides. Trans. R. Soc. Trop. Med. Hyg. 82:558–559. 10.1016/0035-9203(88)90504-4 [DOI] [PubMed] [Google Scholar]
- 72.Barragan A, Spillmann D, Kremsner PG, Wahlgren M, Carlson J. 1999. Plasmodium falciparum: molecular background to strain-specific rosette disruption by glycosaminoglycans and sulfated glycoconjugates. Exp. Parasitol. 91:133–143. 10.1006/expr.1998.4349 [DOI] [PubMed] [Google Scholar]
- 73.Chen Q, Barragan A, Fernandez V, Sundstrom A, Schlichtherle M, Sahlen A, Carlson J, Datta S, Wahlgren M. 1998. Identification of Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) as the rosetting ligand of the malaria parasite P. falciparum. J. Exp. Med. 187:15–23. 10.1084/jem.187.1.15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Carlson J, Nash GB, Gabutti V, al-Yaman F, Wahlgren M. 1994. Natural protection against severe Plasmodium falciparum malaria due to impaired rosette formation. Blood 84:3909–3914 [PubMed] [Google Scholar]
- 75.Sheehy TW, Reba RC. 1967. Complications of falciparum malaria and their treatment. Ann. Intern. Med. 66:807–809. 10.7326/0003-4819-66-4-807 [DOI] [PubMed] [Google Scholar]
- 76.Munir M, Tjandra H, Rampengan TH, Mustadjab I, Wulur FH. 1980. Heparin in the treatment of cerebral malaria. Paediatr. Indones. 20:47–50 [PubMed] [Google Scholar]
- 77.Smitskamp H, Wolthuis FH. 1971. New concepts in treatment of malignant tertian malaria with cerebral involvement. Br. Med. J. 1(5751):714–716. 10.1136/bmj.1.5751.714 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Rampengan TH. 1991. Cerebral malaria in children: comparative study between heparin, dexamethasone and placebo. Paediatr. Indones. 31:59–66 [PubMed] [Google Scholar]
- 79.Jaroonvesama N. 1972. Intravascular coagulation in falciparum malaria. Lancet 299:221–223. 10.1016/S0140-6736(72)90621-6 [DOI] [PubMed] [Google Scholar]
- 80.Zancan P, Mourao PA. 2004. Venous and arterial thrombosis in rat models: dissociation of the antithrombotic effects of glycosaminoglycans. Blood Coagul. Fibrinolysis 15:45–54. 10.1097/00001721-200401000-00008 [DOI] [PubMed] [Google Scholar]
- 81.Evans SG, Morrison D, Kaneko Y, Havlik I. 1998. The effect of Curdlan sulphate on development in vitro of Plasmodium falciparum. Trans. R. Soc. Trop. Med. Hyg. 92:87–89. 10.1016/S0035-9203(98)90969-5 [DOI] [PubMed] [Google Scholar]
- 82.Havlik I, Rovelli S, Kaneko Y. 1994. The effect of Curdlan sulphate on in vitro growth of Plasmodium falciparum. Trans. R. Soc. Trop. Med. Hyg. 88:686–687. 10.1016/0035-9203(94)90230-5 [DOI] [PubMed] [Google Scholar]
- 83.Boyle MJ, Richards JS, Gilson PR, Chai W, Beeson JG. 2010. Interactions with heparin-like molecules during erythrocyte invasion by Plasmodium falciparum merozoites. Blood 115:4559–4568. 10.1182/blood-2009-09-243725 [DOI] [PubMed] [Google Scholar]
- 84.Valle-Delgado JJ, Urban P, Fernandez-Busquets X. 2013. Demonstration of specific binding of heparin to Plasmodium falciparum-infected vs. non-infected red blood cells by single-molecule force spectroscopy. Nanoscale 5:3673–3680. 10.1039/c2nr32821f [DOI] [PubMed] [Google Scholar]
- 85.Juillerat A, Lewit-Bentley A, Guillotte M, Gangnard S, Hessel A, Baron B, Vigan-Womas I, England P, Mercereau-Puijalon O, Bentley GA. 2011. Structure of a Plasmodium falciparum PfEMP1 rosetting domain reveals a role for the N-terminal segment in heparin-mediated rosette inhibition. Proc. Natl. Acad. Sci. U. S. A. 108:5243–5248. 10.1073/pnas.1018692108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Ranjan A, Chitnis CE. 1999. Mapping regions containing binding residues within functional domains of Plasmodium vivax and Plasmodium knowlesi erythrocyte-binding proteins. Proc. Natl. Acad. Sci. U. S. A. 96:14067–14072. 10.1073/pnas.96.24.14067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Glauser BF, Pereira MS, Monteiro RQ, Mourao PA. 2008. Serpin-independent anticoagulant activity of a fucosylated chondroitin sulfate. Thromb. Haemost. 100:420–428. 10.1160/TH08-04-0210 [DOI] [PubMed] [Google Scholar]
- 88.Pacheco RG, Vicente CP, Zancan P, Mourao PA. 2000. Different antithrombotic mechanisms among glycosaminoglycans revealed with a new fucosylated chondroitin sulfate from an echinoderm. Blood Coagul. Fibrinolysis 11:563–573. 10.1097/00001721-200009000-00009 [DOI] [PubMed] [Google Scholar]
- 89.Mourao PA, Boisson-Vidal C, Tapon-Bretaudiere J, Drouet B, Bros A, Fischer A. 2001. Inactivation of thrombin by a fucosylated chondroitin sulfate from echinoderm. Thromb. Res. 102:167–176. 10.1016/S0049-3848(01)00230-4 [DOI] [PubMed] [Google Scholar]
- 90.Fonseca RJ, Mourao PA. 2006. Fucosylated chondroitin sulfate as a new oral antithrombotic agent. Thromb. Haemost. 96:822–829. 10.1160/TH06-06-0304 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.