

Abstract
The standard drug development model uses reductionist approaches to discover small molecules targeting one pathway. Although systems biology analyzes multiple pathways, the approach is often used to develop a small molecule interacting at only one pathway in the system. Similar to that in physics where a departure from the old reductionist “Copenhagen View” of quantum physics to a new and predictive systems based, collective model has emerged yielding new breakthroughs such as the LASER, a new model is emerging in biology where systems biology is used to develop a new technology acting at multiple pathways called “systems therapeutics.”
Diseases are not a simple consequence of abnormality in genes, but reflect the perturbations of the complex system of intracellular networks acted on by complex environmental regulators. The vast majority of previous efforts to elucidate disease and drug response traits have focused on single dimensions of the system. Achieving a more comprehensive, predictive view of common human diseases requires examining living systems in multiple dimensions and at multiple scales. The individual components of complex systems are so tightly coupled that the components cannot be analyzed in isolation, rendering traditional reductionist approaches to science irrelevant, obscuring causal relationships, and lacking an ability to distinguish complexity from mere complication. Biological complexity is an extreme example of complexity, and arises from the inclusion of active, plastic components, nested feedback loops, component multifunctionality, and multiple layers of system dynamics developed through evolutionary processes that are, at least partially, driven through environmental regulators. Systems biology is an emerging approach that seeks to understand the foundations of biology not only at the reductionist molecular level but also by incorporating those reductionist data sets into a larger framework that describes the dynamic interactions of the various levels of information into an emergent, systems level understanding of structure, function, and behaviors.
Our current drug development model is in trouble. Through the recent dominant approach of developing small chemical entities that interact with one specific target and often relying on data predominantly at one level, i.e., genomics, the cost of drug development has risen sharply, while approval rates have fallen.
Even with the introduction of systems biology to the fields of biology and therapeutic development, the mindset in therapeutic development has often remained one of using systems biology for finding the one pathway, or the one target, that is best perturbed to develop the therapeutic. “Finding the magic bullet” is a common phrase that describes this common problem. Instead, the correct thinking needs to shift to one of finding the minimum set of pathways, or the minimum set of targets, using the “minimum molecule set” to perturb in order to best develop a therapeutic. That is, biology is a system, and a particular disease state is the result of multiple perturbations in that system, not just one perturbation. Therefore, only through a thorough understanding of biocircuits in normal and disease states and using computationally intensive biological design-build-test-analyze cycle, with therapeutic molecule production batches based on this process, can we hope to develop safe and efficacious therapeutics through a multitargeted, “systems therapeutic” approach. The approach then is to use a reductionist set (system) of molecules, the minimum molecule set (MMS), that is not overly reductionist so as to be ineffective, but instead use the least number of necessary molecules that are sufficient to realize a safe and efficacious therapeutic. The notion that human diseases are the result of complex interactions among networks has significant implications for drug discovery, leading to the design of molecule combinations that impact entire network states rather than designing drugs that target specific disease associated genes.
We all understand the premise of upward causation, that human diseases and drug response are complex traits that involve entire networks of changes at the molecular level driven by genetic and environmental perturbations. We also understand that changes at the molecular level can induce changes in biochemical processes or broader molecular networks that affect cell behavior and that changes in cell behavior can affect normal tissue or whole organ function, eventually leading to pathophysiological states at the organism level that we associate with disease. Consideration that downward causation exists in these networks is not usually accounted for in our models, nor is the notion that there is no one network level of causation that is privileged.1 That is, an understanding of disease is not likely to arise from a simple understanding of the genome in a disease state, rather multiple levels of understanding are likely needed, such as protein networks, lipid networks, metabolic networks, tissue networks, and environmental networks and that causation flows in all directions among the different networks. For example, recent studies suggest that environmental regulators of retrotransposons are involved in schizophrenia,2 and behavioral effects have profound consequences on cancer.3 As Noble has stated, no one level of causation is privileged, and causation can be considered at many levels, including from the middle out.4 When we ask the age old question, “what came first, the chicken or the egg?” the answer is that the two coevolved through multiple levels of causation, both upward and downward.
Using a mathematical point of view, every component of a biological system can be described in kinetic terms as shown in the Motif of Figure 1. While the level and scale, for example, ion flux through an ion channel on the millisecond scale versus plaque development on a scale of months, will be different for each component, the Motif in each case will be of similar form. Considering the Motif at the level of DNA, a difference in DNA sequence may have a wide variety of possible phenotypic effects, including no effect at all, until the boundary conditions are set, including the actions of many other genes, the metabolic and other states of the cell or organism, and the environment in which the organism exists. Therefore, consideration of a disease state with data at only one level, including DNA sequence alone without the necessary boundary conditions, will result in no predictability of the disease. Further, in analogy with differential equations, knowing only the differences at a given level, for example, as in genome-wide association studies, will poorly predict outcomes. Instead one must integrate the data to understand the necessary and sufficient components and functions. Thus, a differential equation alone has an infinite set of solutions until those possibilities are constrained by the downward causation of boundary conditions. This means that multiple levels of causation are in play, including downward causation. Incorporation of all these data sets into a predictable model is a classic big data problem.
Figure 1.
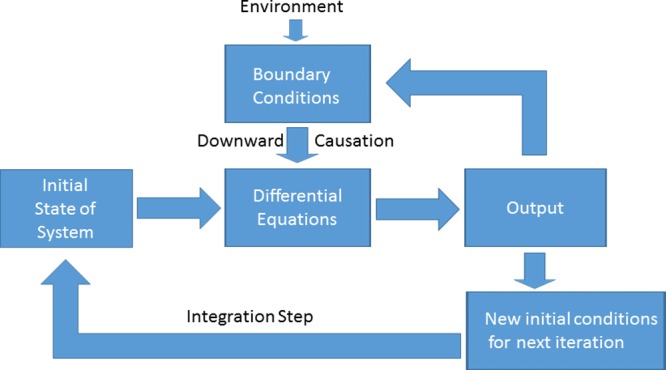
Integration motif. If we consider one pathway in a system, a simple mathematical motif can be described as above. This motif will be only one pathway considered when defining the system, and indeed multiple motifs will be present when considering a disease state, with all of the motifs interacting. (Modified from ref (4). Copyright 2011 The Physiological Society.) For each disease state, multiple motifs at different scales and different levels will be present. For example, in heart attack, one necessary motif to analyze may be ion channel flux represented by protein function on a millisecond scale. Another motif may be plaque formation represented by lipid function on a scale of months. Many other motifs will necessarily be described in heart attack.
We are now at an inflection point in drug development. Success will be achieved through a synergistic interplay between biology and mathematical techniques, especially from network analysis, probability, and statistics studied at multiple levels and scales. Over the past several decades, beginning with early systems biology studies such as those by Hodgkin and Huxley5 on nerve impulse propagation and Noble’s first computer studies of electrical excitation,6 the role of computation in studying biology has grown enormously, and computation has now become an indispensable part of the intellectual mix. To make matters even more computationally intensive, data sets involving different levels, scales, and semantics derived from the clinic, and behavioral and ethological evaluations, may be necessary.7
Attributed to Murray Gell-Mann, and derived from English law, is “That which is not forbidden is mandatory.”8 By this is meant that any allowable process may occur, and therefore, one must try to identify the universality classes (i.e., categories) of relevant phenomena, and then identify and define the likely realizations of those classes. This pattern of discovery is a relatively recent one, arising during the emergence and maturation of quantum electronics started by the Nobel Laureate Charles Townes invention of the MASER, leading to the LASER,9 where laser action is the result of perfect alignment of the crests and troughs of myriad waves of light. Their location and momentum must be theoretically knowable, violating Bohr and Heisenberg’s reductionist “Copenhagen Theory.” Like physics, but lagging behind in time, biology and therapeutic development must move beyond reductionism and simple cataloging to a collective, systems approach to understand emergent, or systems level processes where the therapeutic will likely act at not only complex intramotif (modules) levels but also at the level of the collective of motifs. This viewpoint is meant to be a beginning to the formalized dialogue to realize a systems framework for systems therapeutic development and begin the dialogue “for that which is not forbidden is mandatory” to explore possible computational, systems biology approaches to systems therapeutic development. This means that we explore how systems therapeutics act at multiple targets in the system and will likely involve targets at multiple levels of the system, including genomic, epigenomic, proteomics, cellular, tissue, and even behavioral levels. Considering our mathematical metaphor, using differential equations to describe the system and develop systems therapeutics are necessary. Of equal importance, and a necessity in developing safe, efficacious systems therapeutics, is the use of integration with imposed boundary conditions. Thus, to determine the best MMS, two basic operations must be performed: (1) all relevant data sets from multiple levels should be incorporated into a network analysis to determine the perturbations in the network relevant to the malady, and (2) iterative tests of potential MMSs must be analyzed in the network until the optimal MMS is found that regulates the network and ameliorates the malady. An important example of an emerging systems therapeutics is the stem cell released molecules (SRM), the many types of molecules released from adult stem cells, that are under investigation by many laboratories given their high efficacy and safety profiles.10
Views expressed in this editorial are those of the author and not necessarily the views of the ACS.
The authors declare the following competing financial interest(s): Dr. Maguire is co-owner of BioRegenerative Sciences, Inc.
References
- Noble D. A theory of biological relativity: no privileged level of causation. Interface Focus 2012, 2, 55–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bundo M.; Toyoshima M.; Okada Y.; Akamatsu W.; Ueda J.; Nemoto-Miyauchi T.; Sunaga F.; Toritsuka M.; Ikawa D.; Kakita A.; Kato M.; Kasai K.; Kishimoto T.; Nawa H.; Okano H.; Yoshikawa T.; Kato T.; Iwamoto K. Increased L1 retrotransposition in the neuronal genome in schizophrenia. Neuron 2014, 81, 306–313. [DOI] [PubMed] [Google Scholar]
- Waters D. J. Aging research 2011: exploring the pet dog paradigm. ILAR J. 2011, 52, 97–105. [DOI] [PubMed] [Google Scholar]
- Noble D. Neo-Darwinism, the modern synthesis and selfish genes: are they of use in physiology?. J. Physiol. 2011, 589, 1007–1015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hodgkin A. L.; Huxley A. F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nobel D. Cardiac action and pacemaker potentials based on the Hodgkin–Huxley equations. Nature 1960, 188, 495–497. [DOI] [PubMed] [Google Scholar]
- Maguire G. Using a systems-based approach to overcome reductionist strategies in the development of diagnostics. Expert Rev. Mol. Diagn. 2013, 13, 895–905. [DOI] [PubMed] [Google Scholar]
- Goldenfeld N.; Woese C. Life is physics: evolution as a collective phenomenon far from equilibrium. Ann. Rev. Condens. Matter Phys. 2011, 2, 375–399. [Google Scholar]
- Mead C.Collective Electrodynamics; MIT Press: Cambridge, MA, 2006. [Google Scholar]
- Maguire G. Stem cell therapy without the cells. Commun. Integr. Biol. 2013, 10.4161/cib.26631. [DOI] [PMC free article] [PubMed] [Google Scholar]