

Abstract
Chronic psychological stress appears to accelerate biological aging, and oxidative damage is an important potential mediator of this process. However, the mechanisms by which psychological stress promotes oxidative damage are poorly understood. This study investigates the theory that cortisol increases in response to an acutely stressful event have the potential to either enhance or undermine psychobiological resilience to oxidative damage, depending on the body's prior exposure to chronic psychological stress. In order to achieve a range of chronic stress exposure, forty-eight post-menopausal women were recruited in a case-control design that matched women caring for spouses with dementia (a chronic stress model) with similarly aged control women whose spouses were healthy. Participants completed a questionnaire assessing perceived stress over the previous month and provided fasting blood. Three markers of oxidative damage were assessed: 8-iso-prostaglandin F2α (IsoP), lipid peroxidation, 8-hydroxyguanosine (8-OxoG) and 8-hydroxy-2′-deoxyguanosine (8-OHdG), reflecting oxidative damage to RNA/DNA respectively. Within approximately one week, participants completed a standardized acute laboratory stress task while salivary cortisol responses were measured. The increase from 0 to 30 min was defined as “peak” cortisol reactivity, while the increase from 0 to 15 min was defined as “anticipatory” cortisol reactivity, representing a cortisol response that began while preparing for the stress task. Women under chronic stress had higher 8-oxoG, oxidative damage to RNA (p<.01). A moderated mediation model was tested, in which it was hypothesized that heightened anticipatory cortisol reactivity would mediate the relationship between perceived stress and elevated oxidative stress damage, but only among women under chronic stress. Consistent with this model, bootstrapped path analysis found significant indirect paths from perceived stress to 8-OxoG and IsoP (but not 8-OHdG) via anticipatory cortisol reactivity, showing the expected relations among chronically stressed participants (p≤.01.) Intriguingly, among those with low chronic stress exposure, moderate (compared to low) levels of perceived stress were associated with reduced levels of oxidative damage. Hence, this study supports the emerging model that chronic stress exposure promotes oxidative damage through frequent and sustained activation of the Hypothalamic-Pituitary-Adrenal axis. It also supports the less studied model of ‘eustress’ - that manageable levels of life stress may enhance psychobiological resilience to oxidative damage.
Keywords: Oxidative stress, biological aging, chronic stress, acute stress, DNA/RNA damage, cortisol, hypothalamic-pituitary-adrenal axis, eustress, resilience, reactive oxygen species
Introduction
During periods of intense stress, people are sometimes said to have “aged before your eyes.” This popular idiom reflects the widespread cultural belief that chronic psychological stress can accelerate the aging process. Moreover, it is consistent with accumulating evidence that individuals exposed to chronic stress show signs of accelerated biological aging, such as systemic inflammation and shorter telomere length (Epel et al., 2004; Damjanovic et al., 2007; Gouin et al., 2008; Humphreys et al., 2012). Conversely, it implies the coveted possibility that if we could uncover the mechanisms of this process, we might discover the means to postpone the onset of diseases of aging.
The Free Radical Theory of Aging (Harman, 1956) proposed that accumulation of oxidative damage over time causes cellular aging and contributes to the onset of age-related disease. This implies that it might be possible to slow the physical and cognitive decline associated with aging by mitigating oxidative damage. While oxidative damage is not the sole cause of biological aging, it is widely believed to be an important player in the aging process (Muller et al., 2007). Various oxidative damage markers are elevated or involved in age-related diseases such as diabetes (Monnier et al., 2006; Poulsen et al., 2012), cancer (Valko et al., 2006), neurodegenerative diseases (Nunomura et al., 2012; Poulsen et al., 2012) and cardiovascular disease (Gutierrez et al., 2006). Oxidative damage is caused by reactive oxygen species (ROS) unmitigated by antioxidants. Under healthy conditions, ROS production is primarily a byproduct of daily mitochondrial respiration (Gutierrez et al., 2006) that fuels basic metabolic processes. Oxidative stress reflects a state of cellular imbalance, in which ROS production exceeds antioxidant mechanisms that neutralize ROS, resulting in oxidative damage to nearby molecules, such as DNA, RNA and lipids. Some of the most interesting markers in terms of their relevance to disease processes include 8-iso-prostaglandin F2α (IsoP), a marker of lipid peroxidation, 8-hydroxyguanosine (8-OxoG), a marker that primarily reflects damage to RNA, and 8-hydroxy-2′-deoxyguanosine (8-OHdG), a marker of damage to DNA (Poulsen et al., 2012). IsoP and 8-OHdG are among the most commonly reported markers, in part due to their stability in vivo (Akagi et al., 2003; Milne et al., 2007). 8-oxoG is comparatively less common, but increasingly important (Poulsen et al., 2012); for example, urinary 8-oxoG (but not 8-OHdG) predicted long-term mortality from type 2 diabetes in a recent study (Broedbaek et al., 2011). Although all three markers are considered indicators of oxidative stress, they do not always correlate with one another (Watters et al., 2009), and they may differ for several reasons – e.g., DNA, RNA and lipids differ in their proximity to mitochondria, the primarily producer of ROS, they employ different mechanisms of repair or elimination, and each damage signature may have different pathophysiological consequences (Furukawa et al., 2004; Poulsen et al., 2012).
Psychological stress and distress have been associated with higher levels of oxidative damage (Irie et al., 2003; Epel et al., 2004; Forlenza and Miller, 2006; Gidron et al., 2006). For example, pre-menopausal women caring for a chronically ill child (considered a model of chronic stress exposure) who endorsed greater perceived stress had higher oxidative stress (indexed by the ratio of F2-Isoprostanes, or 8-iso-prostaglandin F2α, to vitamin E) and shorter telomere length, a marker of cellular age (Epel et al., 2004). The mechanism remains unclear, and is likely mediated in part by stress-related hormones (e.g., cortisol) as well as health behaviors (Radak et al., 2005; Ballal et al., 2010). If stress-arousal plays an important role, this would suggest that stress-management should be a core component of preventative interventions designed to improve healthy aging.
The glucocorticoid hormone cortisol represents a potentially important mechanism linking chronic stress with accelerated aging. When events are perceived as stressful, and particularly when the stressor evokes negative affect or social threat (Dickerson and Kemeny, 2004), this may induce increased cortisol secretion, which mobilizes the metabolic energy to cope with stressors. One standardized experimental task used to elicit a stress-induced cortisol increase (i.e., “reactivity”) is the Trier Social Stress Task (Kirschbaum et al., 1993). Individuals exhibit relatively stable individual differences in cortisol reactivity to acute stress (Kirschbaum et al., 1995). Hence, individuals with high cortisol reactivity who are exposed to chronic stressors that persistently evoke a reactivity response are more likely to exhibit adverse health effects. 24-hour urinary excretion of cortisol has previously been linked cross-sectionally with elevated markers of DNA and RNA damage in older adults (Joergensen et al., 2011); however, that study did not directly link cortisol or oxidative damage markers with psychological stress or stress-induced cortisol reactivity.
It is generally assumed that cortisol reactivity is best captured by peak secretion, occurring between 21-40 minutes following stressor onset (Dickerson and Kemeny, 2004). However, converging evidence from psychoneuroendocrinology and systems biology (Aschbacher et al., 2012) suggests that transient stress-arousal responses should also be assessed by other features of the dynamic response. It takes approximately 10 minutes for a stress response initiated in the hypothalamus to promote a detectable increase in peripheral cortisol (Sapolsky et al., 2000), and these kinetics may have clinical importance for health (Aschbacher and Kemeny, 2011; Aschbacher et al., 2012). When the principles of system robustness (Kitano, 2007) are applied to biological stress systems, robustness theory suggests that a heightened response to stress may help the body optimize performance to frequently encountered stressors. This may occur during chronic stress. In animal models, chronic stress exposure reorganizes neural networks regulating neuroendocrine function, providing an anatomical basis for more excitable cortisol responses to stress (Miklos and Kovacs, 2012). Hence, we propose that “anticipatory” cortisol reactivity (i.e., increases during psychological anticipation of the stressor) may be an important marker of a central nervous system that is “primed” for heightened stress-reactivity. Chronically stressed individuals may develop a heightened tendency to anticipate stress and mount a rapid cortisol response, which could increase vulnerability to oxidative stress and accelerate biological aging (Drabant et al., 2011; O'Donovan et al., 2012; Tomiyama et al., 2012).
Although the term “stress” carries a negative connotation, evidence suggests that under certain circumstances, stress exposures may have the potential to enhance an organism's performance and resilience. The first evidence that low-to-moderate doses of stress may have beneficial effects (“eustress”) emerged over a century ago as an “inverted U” relationship between arousal and performance (Yerkes and Dodson, 1908). This psychological principle finds its biological doppelgänger in the myriad examples of inverted U relationships between glucocorticoid actions and various physiological targets (Sapolsky, 1997). For example, cortisol bears an inverted U-shaped relationship with mitochondrial function, a key regulator of oxidative stress (Du et al., 2009). In cell culture models, brief administration of high-dose cortisol resulted in improved mitochondrial function and neuroprotective effects, whereas long-term high-dose cortisol administration dramatically decreased mitochondrial function and promoted cell death (Du et al., 2009). Moreover, at low concentrations, reactive oxygen and lipid species activate cytoprotective pathways that increase antioxidants (Gutierrez et al., 2006). Corticosterone administered at the time of immune activation in doses that mimic a physiological stress response enhances the ensuing immune response, while pharmacological doses or chronic administration are immune-suppressive (Dhabhar and McEwen, 1999).
Extrapolating from these U-shaped relationships, this study hypothesized that greater perceived stress over the previous month and anticipatory cortisol reactivity to an acute stress task will be significantly associated with increased oxidative damage among chronically stressed caregivers. In contrast, among low-stress controls, these same factors either will be not be significantly associated with oxidative damage, or will be associated with decreased oxidative damage, a manifestation of eustress and psychobiological resilience (Figure 1). The conceptual framework was tested simultaneously in a full moderated mediation model with two primary component hypotheses: 1) anticipatory cortisol reactivity will mediate associations between perceived stress and oxidative damage markers, and 2) the indirect path from perceived stress to oxidative damage via cortisol reactivity will be significant among chronically stressed caregivers, but not among age-matched low-stress controls. Secondarily, we explored the “eustress” hypothesis in controls, or the idea that moderate stress may have salutary effects on oxidative damage.
Figure 1. Moderate Stress May Enhance Resilience.

Methods
Participants
This study was conducted as part of a larger study of caregiving stress and biological aging among post-menopausal women. A subset of 48 participants from the larger study participated in an acute laboratory stress task and had data available for analysis in the current study. Twenty-five participants were caregivers of a relative with dementia and twenty-three were age-matched controls with healthy spouses. Caregivers had provided care for 4.7 years on average (range: 8 months-11.42 years). The mean age was 63 years (range: 51-79), the median income was $70,000-79,000, and the racial/ethnic breakdown was 90% (n=43) Caucasian, 2% (n=1) African American, and 8% (n=4) Asian/Pacific Islander. Exclusion criteria included chronic medical conditions (cardiovascular disease, cancer, diabetes, and autoimmune diseases), current smoking, or use of medications known to affect stress hormones. To ensure a healthy control group with no mental illness, controls taking antidepressants were excluded. Participants were deemed healthy by virtue of self-report, having passed a medical exam, and having standard lab values from their baseline fasting blood draw that fell within the normal range (e.g., glucose, liver, kidney function tests and combined blood count). Approximately one week after completing their basal fasting blood draw and questionnaire packet, participants came in for the afternoon acute stress task. The study protocol was approved by the Institutional Review Board of the University of California San Francisco.
Perceived Stress Scale (PSS)
The Perceived Stress Scale-10 (Cohen et al., 1983) is a standard 10-item questionnaire that assesses subjective perceptions of stress over the previous month. The scale has been normed in several large national surveys, and the average PSS scores in 2006 and 2009 among women was approximately 16 (Cohen and Janicki-Deverts, 2012). Moreover, the PSS has been previously shown to relate to measures of oxidative stress and cellular age among pre-menopausal caregiving women (Epel et al., 2004). Items reflect uncontrollability, unpredictability and feeling that demands outweigh coping resources. Response options form a 5-point Likert scale ranging from 0=never to 4=very often. Cronbach's alpha was .91 in two large samples from 2006 and 2009 (Cohen and Janicki-Deverts, 2012), and .94 in the current sample.
Negative Affect
Using the Positive and Negative Affect Schedule (PANAS), participants reported the degree to which they experienced various emotions in the past week (Watson et al., 1988). To explore the potential resilience-building aspects of moderate levels of stress and distress, an a priori decision was made only to use the negative subscale, which assesses feeling scared, troubled, sad, guilty and irritable (Cronbach's alpha = .93 in this sample). This was used as a secondary measure to the PSS.
Acute Stress Task Protocol
The Trier Social Stress Task (TSST)(Kirschbaum et al., 1993) is a gold-standard laboratory stress task used to evoke stress-arousal responses. In this study, because many participants were retired, they did not do a mock job interview, but instead discussed their strengths and weaknesses and completed a math task in front of an evaluative audience. All participants completed the TSST between 1400h and 1500h to control for diurnal rhythm variation. Participants rested for one hour, sitting quietly listening to music, prior to collection of the baseline sample. Salivary samples were collected via the passive drool method in 2 mL SaliCaps tubes (IBL Hamburg, Germany) at multiple time-points during the acute stress task, which permitted quantification of anticipatory and peak cortisol reactivity: 0 (baseline), 15 (after 5 min instructions, 5 min preparation, 5 min speech) and 30 (to capture peak cortisol). This study's a priori hypothesis was that the anticipatory cortisol response, quantified as the change from rest to 15 minutes, might be equally as or more important in predicting health indicators than the peak cortisol response, quantified as change from rest to 30 minutes. This measure was considered anticipatory because neuroendocrine responses would likely have been triggered in the central nervous system immediately upon preparation to give the speech in order to result in a significant cortisol increase within roughly 10 minutes (from the end of the instructions and beginning of preparation). Hence, this may represent increased situational anticipatory anxiety or threat (Drabant et al., 2011). The importance of the anticipatory increase was additionally based upon previous findings (Aschbacher et al., 2012) and kinetic studies showing non-linear association rates of glucocorticoid steroid-receptor complexes (Yeakley et al., 1980).
Confirmatory Measures: Task-Related Anticipatory Threat & Effort
Both social evaluative threat and task effort can predict cortisol reactivity to acute stress (Dickerson and Kemeny, 2004). As a secondary analysis, to better clarify the specific role of psychological processes involved in stress anticipation, we included a previously published measure of pre-task emotional and cognitive threat. This measure was composed of two subscales - 3 emotionally focused items (e.g., “How anxious do you feel about the upcoming tasks?” and 3 cognitively focused items (e.g., “The upcoming tasks will be very demanding;” The Cronbach's alphas for this sample were .77 and .86 respectively) (O'Donovan et al., 2012). When conceptualizing reasons for very low threat appraisals, it is possible that certain people did not engage in the stress task. We sought to rule out the possibility that individuals reporting extremely low stress were simply not motivated to put effort into the task. Hence, it could be that trait-like disengagement (i.e., being overly blasé) might account for any apparent “eustress” associations that might be found in the low-stress group (i.e., more disengagement, very low stress, higher oxidative damage). Therefore, we assessed task effort with one item asked immediately after the task, “I put a lot of effort into completing the task successfully.”
Salivary Cortisol Assays
Cortisol samples were frozen at the end of the session and assayed in batch in Germany (laboratory of Clemens Kirschbaum in Dresden, Germany). Salivary free cortisol levels were measured using a commercial chemiluminescence immunoassay (CLIA; IBL Hamburg, Germany). The intra-assay coefficients of variation were 2.9% and 7.7% for high and low levels of cortisol respectively, while the inter-assay coefficients of variation were 5.7% and 9.1% for high and low respectively. All cortisol levels were more than three times larger than the lower limit of detection, which was 0.16 ng/mL. All cortisol values were log-transformed to improve the normality of the distributions.
Oxidative Damage Assays
One week prior to the acute stress task, participants visited the lab for a fasting morning blood draw. A venous forearm catheter was inserted, participants rested for 20 minutes, and blood was drawn between 0730h and 0800h. Blood samples were centrifuged at 1300g for 10 minutes at room temperature, and the serum was aliquotted into cyrovials and stored at -70° C for future analysis. 8-hydroxyguanosine (8-oxoG), 8-hydroxy-2′-deoxyguanosine (8-OHdG) and 8-iso-prostaglandin F2α (IsoP) were measured using the API 4000™ QTRAP® LC/MS/MS System by Kronos Science Laboratory. This system utilizes an Agilent 1200 series liquid chromatography, which interfaces directly with a triple quadrupole/linear ion trap mass spectrometer (Applied Biosystems, Foster City, CA), and is equipped with a TurboIonSpray ionization source. To assess 8-oxoG and 8-OHdG, the serum sample was mixed with an internal standard of O18-8-hydroxy-2′-deoxy-guanosine and an organic solvent was added to precipitate proteins. After centrifugation, the supernatant was evaporated in a vacuum, and the residue was redissolved with LC mobile phase and injected into an HPLC system that separates the analytes from the matrix interference. Under multiple reaction-monitoring (MRM) detection mode, ion pairs of m/z 284/168 and 300/168 were used to quantify 8-OHdG and 8-OxoG respectively. The intra and inter precision of the procedure were monitored by two levels of serum controls. The coefficients of variation for 8-oxoG and 8-OHdG ranged from 4% to 8% (intra) and 8% to 12% (inter). To determine IsoP, serum sample was mixed with an internal standard of 8-isoprostaglandin F2α-d4 (IsoP-d4) and an organic solvent was added to precipitate proteins. After centrifugation, the supernatant was purified through a C18 solid phase extraction (SPE) cartridge on an automated SPE workstation (RapidTrace; Zymark, Hopkinton, MA, USA). The eluent is evaporated in vacuum, and the residue was reconstituted in mobile phase and injected onto HPLC system. Under MRM detection mode, the ion pair of m/z 353/193 was used to quantify IsoP. Recoveries are consistently in excess of 90%. The coefficients of variation ranged from 4% to 7% (intra) and 6% to 11% (inter). In order to rule out the possibility that differences in blood measures of oxidative damage were not likely due to poorer kidney function among caregivers leading to altered clearance rates, plasma creatinine was assayed and group levels were compared.
Health Behaviors & Medical Factors
All participants were characterized on the following health behaviors, which could be related to oxidative damage: physical activity (frequency*duration of moderate or vigorous activity over the past month), alcohol consumption in the past month (yes/no), and having smoked 100 or more cigarettes in one's lifetime. Smoking was an exclusion criterion for the study so there were no current smokers included. Body mass index (BMI) was included as an indicator of overweight, which may contribute to oxidative stress (Furukawa et al., 2004). Medical histories at baseline were assessed using self-reports for the presence of conditions (yes/no): hypertension (n=16), hypercholesterolemia (n=14), chronic pain (n=8) and arthritis/osteoarthritis (n=10). Participants also reported whether or not they currently use any of the following medications or supplements, potentially related to stress system hormones or oxidative stress: non-steroidal anti-inflammatory medications (NSAIDs) (n=11), antidepressants (selective serotonin reuptake inhibitors or tricyclics) (n=7), statins (n=9) and vitamin supplements that might contain antioxidants (n=20).
Data Analysis
All physiological outcomes and proposed mediators were examined for deviations from normality both visually and using the K-S test. Cortisol variables were log-transformed, which normalized their distributions. Based on an a priori decision, outliers greater than 3 standard deviations were winsorized to be 5% greater than the next largest value (one outlier was winsorized for IsoP and for 8-OHdG). A small number of missing data-points (roughly 2%) were imputed using the estimated maximization missing data imputation package in SPSS 20.0. Of the imputed data-points, one point was missing each for: 8-oxoG, 8-OHdG, IsoP, basal cortisol, negative PANAS and PSS (each point missing was from a different participant with the exception that the same participant was missing a datapoint for both 8-oxoG and 8-OHdG). Three and four points respectively were imputed for cortisol at immediately post-speech and 30 min post-stress task onset. The overall pattern of results was similar with and without data imputation. Three individuals declined to state their income, and as these points may have been systematically missing (individuals who decline to state their income may differ from individuals who are comfortable doing so), they were not imputed. A moderated mediation model was used to test whether heightened anticipatory cortisol stress-reactivity mediated the relationship between perceived stress and oxidative damage among individuals exposed to a chronic stressor (i.e., caregivers) (Figure 1). All hypotheses were tested at the same time using a bootstrapped moderated mediation model with the aid of the “Process” macro for SPSS (Hayes, 2012). All variables were normalized prior to model entry to facilitate centering and interpretation of coefficients. Regression analyses were used to identify coefficients for individual paths. As the literature suggests that a broad range of factors could potentially impact oxidative damage, relevant covariates included demographics (i.e., age, BMI, race/ethnicity (white versus non-white), having a college education or better), health behaviors (i.e., being a previous smoker, alcohol use, physical activity), medication use and pre-existing physician-diagnosed medical conditions (see methods above for details). To preserve power given the number of covariates, each one was entered individually into the full model.
Results
Group Comparisons
Caregivers (CG) and controls (NC) did not significantly differ on age, BMI, Caucasian race/ethnicity, education, being a previous smoker, alcohol use, and the presence of any physician-diagnosed medical problem (all p's>.12). There was a non-significant trend for CG to report less exercise than NC (p=.09). CG were significantly more likely to use NSAIDs (9 CG versus 2 NC, p=.04), but reported no differences in the use of statins or vitamins potentially containing antioxidants (all p's>.60). As expected, CG reported significantly greater levels of perceived stress than controls over the previous month (p<.001). CG also had significantly greater anticipatory (15-min) but not peak (30-min) cortisol reactivity than NC (p's=.003 and .16 respectively; Figure 2), even when controlling for demographic, health behavior and medical factors, as reported in a previous analysis (Epel et al., 2010). CGs and NCs did not differ on cortisol levels at baseline (p=.41). Furthermore, CG had significantly higher levels of 8-oxoG (p<.001) and non-significantly higher levels of IsoP and 8-OHdG compared to controls (Table 1). The significant association of caregiving with 8-oxoG, remained when sequentially controlling for all covariates (as specified in the data analysis section). Duration of care was not significantly associated with any cortisol or oxidative damage marker; hence it was not included in subsequent analyses. In the whole sample, greater BMI was also significantly associated with higher IsoP, a marker of damage to lipids (β=.32, t(45)=2.29, p=.03), but not with 8-oxoG, a marker of genomic damage. Plasma creatinine, a marker of kidney function, did not significantly differ between CG (M±se = .847±.028) and NC (M±se = .864±.039) (t(46)= 0.350, p=.728).
Figure 2. Effects of Chronic Stress on Salivary Cortisol Reactivity to Acute Stress.
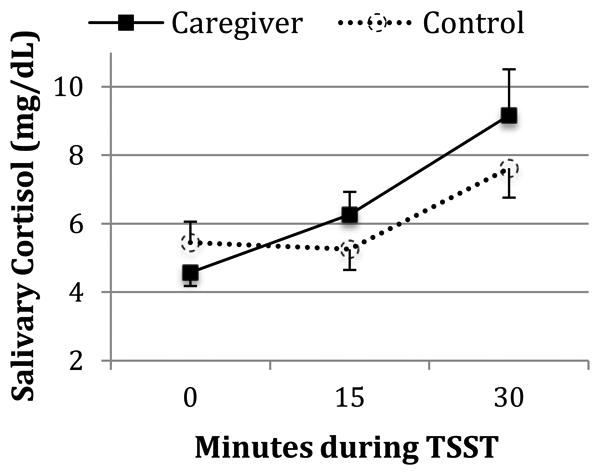
The salivary cortisol values depicted above are untransformed means and their standard errors. The anticipatory increase in cortisol from 0 to 15 minutes is significantly greater among caregivers than controls (p<.01), whereas the peak 30-minute increase does not significantly differ by group (p=.16). The groups did not significantly differ in cortisol levels at the TSST baseline (0 min) (p=.41).
Table 1. Comparison of Caregivers and Controls (Means & Standard Errors).
| Caregivers (n=25) | Controls (n=23) | T-test (df) | P-Value | |
|---|---|---|---|---|
| Perceived Stress | 20.360 (1.353) | 5.804 (0.838) | 9.01 (40.75)‡ | >.001*** |
| Anticipatory Cortisol Reactivity (mg/dL) | 0.100 (0.028) | -0.015 (0.022) | 3.15 (46) | >.01** |
| Peak Cortisol Reactivity mg/dL) | 0.213 (0.050) | 0.117 (0.044) | 1.43 (46) | .16 |
| F2-Isoprostanes (ng/mL) | 0.044 (0.004) | 0.042 (0.003) | 0.35 (46) | .73 |
| Serum 8-oxoG (ng/mL) | 0.036 (0.002) | 0.022 (0.002) | 3.83 (46) | > .001*** |
| Serum 8-OHdG (ng/mL) | 0.249 (0.032) | 0.187 (0.032) | 1.36 (46) | .18 |
p ≤.001,
p ≤.01,
critical alpha = .05.
Unequal variance per significant Levine's test.
Zero Order Pearson Correlations
As the first step in testing the full model, raw Pearson correlations between the key variables in the full model were determined for CGs and NCs separately, in order to set the stage for exploration of potential moderation effects (Table 2). As hypothesized, among chronically stressed CGs, higher perceived stress was significantly associated with greater anticipatory and peak cortisol reactivity (p's ≤.01). Moreover, also among CGs, anticipatory cortisol reactivity was associated with significantly higher levels of 8-oxoG and IsoP (p's ≤.01), but not 8-OHdG. In contrast, peak cortisol was not significantly associated with any oxidative damage marker among CGs. The relationships between PSS and the oxidative damage markers were also not significant among CGs; notably, current experts in meditational analyses assert that it is not necessary for this path to be significant in order for a significant indirect effect to be present (Hayes, 2009; Mackinnon and Fairchild, 2009).
Table 2. Zero Order Correlations among Psychological Stress, Threat, Cortisol Reactivity and Oxidative Damage Markers by Group.
| CG=top-right/NC=bottom-left | Perceived Stress | Negative Affect | Anticipatory Threat | Baseline Cortisol | Anticipatory Cortisol | Peak Cortisol | 8-oxoG | 8-OHdG | IsoP |
|---|---|---|---|---|---|---|---|---|---|
| Perceived Stress | --- | .346† | .286 | .095 | .565** | .514** | .044 | -.224 | .282 |
| Negative Affect | .309 | --- | .274 | .235 | .445* | .218 | .490* | .397* | .435* |
| Anticipatory Threat | .345 | .148 | --- | -.056 | .509** | .220 | .312 | -.099 | .638** |
| Baseline Cortisol | -.182 | -.037 | -.106 | --- | -.201 | -.274 | . 348† | -.101 | .114 |
| Anticipatory Cortisol | .028 | -.127 | .170 | -.272 | --- | .592* | .516** | -.053 | .559** |
| Peak Cortisol | .179 | -.109 | .171 | -.399† | .439* | --- | -.079 | -.071 | .317 |
| 8-oxoG | -.377† | -.019 | -.211 | .017 | .151 | -.140 | --- | .123 | .467* |
| 8-OHdG | -.353† | -.278 | -.290 | -.096 | .110 | -.050 | .093 | --- | -.149 |
| *IsoP | -.425* | .007 | -.442* | .281 | -.225 | -.484* | .220 | .157 | --- |
Note:
p ≤.01,
p ≤.05,
p ≤.10,
critical alpha = .05, N=48, caregivers (CG: n=25), non-caregivers (NC: n=23). Pearson correlations among chronically stressed CG are shown to the top-right of the diagonal, while correlations among NC are shown to the bottom-left. Anticipatory threat refers to the emotional items as specified in the results.
Intriguingly, among NCs only, higher PSS scores were not significantly associated with anticipatory or peak cortisol reactivity using Pearson correlations; however, higher PSS scores were associated with significantly decreased IsoP (Table 2; p = .04) and borderline trends toward decreases in 8-oxoG and 8-OHdG (p's<.10). This relationship with IsoP remained borderline significant (p≤.07), in regression analyses controlling for all covariates. Among low-stress controls, greater peak cortisol reactivity was also significantly related to lower basal levels of IsoP (p=.02), while controlling for all covariates. The pattern of significance for the correlations in Table 2 was not altered when retested in the raw (not imputed) data, except that the relation between PSS and IsoP among controls changed from p=.04 to p=.06.
Full Model: Moderated Mediation
The full model of moderated mediation was tested first without covariates, and secondarily, by entering covariates sequentially to avoid over-fitting. Figure 3 depicts the unadjusted statistical test of moderated mediation. As hypothesized, a significant indirect effect was found (p ≤.01), consistent with a model in which high perceived stress was significantly associated with oxidative damage via anticipatory cortisol reactivity, but only among chronically stressed CGs (Table 3). In contrast, no significant mediating relationship was present among controls (see bootstrapped indirect effect coefficients and confidence intervals in Table 3 and note that this held for critical alphas of .05 and .01). As 8-OHdG was not significantly associated with cortisol reactivity, it did not meet mediation criteria and was not examined in the mediation model. We did not test an alternative model using peak cortisol, because it was not significantly associated with any oxidative damage marker in CGs (Table 2), and therefore did not meet criteria for mediation. Consistent with the primary study hypotheses, the indirect effects for both IsoP and 8-oxoG remained significant when sequentially controlling for all covariates. For the outcome 8-oxoG, the interaction between chronic stress exposure (i.e., caregiver status) and anticipatory cortisol reactivity trended in the same direction as with IsoP, but was of borderline significance (Figure 3).
Figure 3. Anticipatory Cortisol Reactivity to Acute Stress Mediating the Relationship between Perceived Stress and Oxidative Damage in High versus Low Stress Groups.

*** p≤.001, ** p≤.01, * p≤.05, † p≤.10, N=48. Note: Standardized regression coefficients and significance levels for each path are given, showing F2-Isoprostanes first with 8-oxoG (in parentheses). The mediating path analytic models with regression coefficients are shown separately for the chronically stressed caregivers (top) and low-stress controls (bottom). Note that the relationship from perceived stress to cortisol remains the same regardless of which oxidative damage marker is used as the final outcome of the mediation model.
Table 3. Tests of moderated mediation: Bootstrapped indirect effects of perceived stress on oxidative damage via anticipatory cortisol reactivity by group.
| F2-Isoprostanes | 8-oxoG | |||||||
|---|---|---|---|---|---|---|---|---|
| Status | ß | SE | LL | UL | ß | SE | LL | UL |
| Chronic Stress Group | 0.732* | 0.331 | 0.071 | 1.927 | 0.645* | 0.267 | 0.120 | 1.585 |
| Low-Moderate Stress Group | -0.247 | 0.233 | -1.028 | 0.279 | -0.200 | 0.243 | -0.428 | 1.009 |
Note:
p ≤ .01 for the indirect effect;
N = 5000 Bootstrapping resamples; LL and UL = Lower level and upper level of the bias-corrected, accelerated confidence interval at critical alpha of .01.
Confirmatory Analyses
In order to better understand whether the relationship between stress levels and lower oxidative damage among NCs was a likely indication of eustress or a random occurrence, we additionally examined two secondary indicators of stress: PANAS negative affect and anticipatory cognitive & emotional threat. These results (Table 2) reveal that, among chronically stressed individuals, higher negative affect and anticipatory emotional threat are associated with greater anticipatory cortisol reactivity and oxidative damage, primarily IsoP). Intriguingly, the opposite pattern is present in low-stress individuals (e.g., lower anticipatory emotional threat is significantly associated with greater oxidative damage per IsoP). Cognitive threat was not related to oxidative damage in either group. Next, we examined whether the “paradoxical” associations that were significant in the low stress group between psychological factors (PSS and anticipatory emotional/cognitive threat) and oxidative damage became nonsignificant when controlling for task effort. In every case, significant associations remained significant in the same direction. Moreover, when controlling for task effort, the association between PSS and 8-oxoG, which was borderline in uncontrolled analyses, became significant (β= -.45, t(19)= -2.18, p=.04), while effort was not a significant covariate in any of these analyses.
Discussion
Psychological stress and its biological mediators are increasingly recognized as important factors that influence the rate of biological aging. However, the field is still in the process of defining a road map to clarify under what circumstances psychological stress activates “eustress” responses and psychobiological resilience mechanisms, or alternatively, promotes premature aging and morbidity (Dhabhar et al., 2012). Whereas substantially more is known with regards to the beneficial and adverse effects of stress and stress hormones on the immune system (Dhabhar, 2009; Dhabhar et al., 2012), comparatively little is known about how they impact the oxidative milieu and markers of oxidative damage. We tested the idea that, in the sample exposed to chronic stress, the typical assumption regarding a pattern of ‘bad stress’ would emerge – in other words, indices of stress (perception, cortisol) would be associated with elevated markers of oxidative stress. In contrast, in the low stress sample, the pattern of ‘good stress,’ or eustress, would be observed, where those with low-to-moderate stress would have lower markers of oxidative damage than those with very low stress.
A consistent pattern emerged whereby chronically stressed caregivers who reported greater perceived stress, negative affect and threat had significantly greater cortisol increases while anticipating a stressful event. However, intriguingly, chronically stressed women exhibited no differences from age-matched low-stress women in the peak cortisol increase during the actual stressful event itself. In turn, heightened anticipatory cortisol responses to stress were associated with higher levels of two different markers of oxidative damage, 8-oxoG (a marker that primarily indicates RNA damage) and the isoprostane 8-iso-prostaglandin F2α (a marker of lipid peroxidation), but not with 8-OHdG (a marker of DNA damage). Oxidative stress appears to contribute to biological aging (Muller et al., 2007), and the biological signatures of oxidative damage are elevated in age-related diseases, such as neurodegenerative, metabolic, cardiovascular conditions and cancer (Gutierrez et al., 2006; Monnier et al., 2006; Valko et al., 2006; Nunomura et al., 2012; Poulsen et al., 2012). Hence, heightened cortisol release during anticipation of stress may be an important mechanism to explain how psychological stress promotes aging processes and increases risk for diseases among at-risk individuals. Though cross-sectional, these data illuminate our understanding of the likely psychobiological pathways involved in resilience versus vulnerability to stress-related biological aging.
These findings are particularly striking because they highlight the importance of psychobiological processes that take place during anticipation of an upcoming stressor, such as expectations, thoughts and emotions experienced in preparing for a stressor. Intriguingly, it was anticipatory rather than peak cortisol reactivity, which best explained the relationship between perceived stress and heightened oxidative damage among chronically stressed caregivers. Moreover, in the chronically stressed group, anticipatory emotional threat (a psychological self-report measure) was also significantly associated with anticipatory cortisol responses and IsoP, but not with peak cortisol. We suggest that the anticipatory neuroendocrine response was activated during the time period when the instructions were read and the preparation for the stress task began, but before individuals actually began the public speaking task or received any evaluative feedback. From a neurobiological perspective, the brain circuitry activated during anticipation of a stressful event differs from that which is activated during the event itself (Drabant, 2011). The stress individuals experience as they anticipate a future event arises from their emotional and cognitive representations of what they believe or fear they will experience, rather than what they actually experience during the event. Moreover, some evidence suggests that chronic stress exposure may change the morphology of the brain regions that regulate neuroendocrine activity, promoting increased excitability of CRH neurons during stress (Miklos and Kovacs, 2012). Hence, chronic stress may predispose individuals to anticipate stress before the event occurs and mount a stronger corresponding cortisol response. These data, building upon previous findings in this sample (O'Donovan et al., 2012), underscore the important role of stress anticipation in relation to accelerated biological aging. Whereas this study employed brief subscales to assess anticipatory emotional threat, future studies of this nature may consider employing more in-depth assessment tools of anticipatory threat and challenge, such as the Primary Appraisal Secondary Appraisal scale (Gaab et al., 2005; Juster et al., 2012), which have been psychometrically validated against neuroendocrine acute stress reactivity to the TSST.
Chronic stress-related alterations in biomarkers are often interpreted as a manifestation of “wear and tear” on the body or allostatic load (McEwen, 2004), as though the system were a rubber band that lost elasticity. However, an alternative and nonexclusive interpretation is that these changes may represent not so much a loss of resources, but a reallocation. Chronic stress reorganizes the structure of the central nervous system, providing an anatomical basis for changes in the dynamics (e.g., anticipatory reactivity) of stress hormones (Miklos and Kovacs, 2012). These changes may constitute an “intended” optimization of neuroendocrine system function, which has unfortunate “side effects”– i.e., the optimization increases short-term capacity for efficient responses to stress, while increasing long-term vulnerability to age-related disease. Seen from this light, heightened cortisol reactivity may not be a dysfunction or failure of the system per se, but an adaptation to cope with a demanding environment, which may have short term benefits to survival but longer term adverse consequences for healthy aging. We did not, however, investigate the role of delayed cortisol recovery, which might provide more support for the wear and tear model.
Intriguingly, these data also paint the encouraging picture that low to moderate stress may have beneficial or resilience-enhancing effects among post-menopausal women. Specifically, among control women whose levels of chronic perceived stress were well below the 2009 national norms for women (i.e., a PSS score of approximately 16) (Cohen and Janicki-Deverts, 2012), greater perceived stress and anticipatory threat were associated with significantly reduced levels of oxidative damage. This finding was not due to lower self-reported task effort in the very low stress women. Although seemingly paradoxical, this finding was replicated using several indicators of stress and distress. Moreover, the finding is consistent with the Yerkes-Dodson theory of stress and performance (Yerkes and Dodson, 1908) and the well-documented non-linear effects of cortisol on many target tissues (Sapolsky, 1997). However, it should be noted that overall levels of oxidative damage, as measured by 8-oxoG, were still significantly higher in chronically stressed individuals. Hence, as a whole these results suggest that there is a non-linear relationship between stress and markers of an adverse biochemical milieu, whereby at the extreme end of the spectrum (i.e., very low chronic stress and threat responses), biological systems may not be functioning at optimal fitness, but the profile is still “healthier” than in a chronically stressed system. The implication is that maximal resilience may be found at the nadir of a U- or J-shaped curve. Hence, there may be a “happy medium” of life stress.
Although cross-sectional, these data suggest that cortisol is a plausible mediator of oxidative damage among chronically stressed individuals, whereas unidentified counter-regulatory mechanisms likely contribute to the existence of a non-linear relationship and the contrary pattern of results in non-chronically stressed individuals (Sapolsky, 1997). Cortisol reactivity may also be an indicator of the magnitude of the amygdalar response to threat, particularly in the chronically stressed group. Certain types of physiological stress, such as periodic exercise, are known to promote long-term decreases in circulating indicators of oxidative damage, apparently due to upregulation of antioxidant and other cytoprotective mechanisms (Radak et al., 2005). While purely speculative, it is conceivable that low-to-moderate episodic psychological stress exposure could improve psychobiological resilience to oxidative damage in a manner similar to exercise. Future research should include a more thorough assessment of relevant counter-regulatory mechanisms such as indicators of the nitric oxide pathway, anti-oxidants, and cellular surveillance mechanisms.
This study has a number of limitations. While these data are consistent with a moderated mediation model, they are cross-sectional, and establishing true mediation will require replication in a longitudinal design. This study was not designed to disentangle the effects of caregiving as an objective exposure factor from the subjective perception of stress, and we did not recruit caregivers low in perceived stress or high-stress NCs. Hence the differences in the relation between perceived stress and oxidative stress may reflect not only a different functional relationship of moderate versus extreme perceived stress, but also the chronically stressful experience of caregiving. Moreover, it is possible that relations interpreted herein as eustress may instead reflect a suppressive coping style, in which individuals who report very low levels of stress and threat response exhibit higher oxidative damage because they are in fact suppressing or are unwilling to disclose their true negative emotions. However, if this were true, we might expect to see greater physiological arousal (e.g., cortisol) in the very low stress participants as a result of emotional suppression, which we did not find. Furthermore, future studies should explore anxiety, avoidant coping styles or negative affectivity as personality traits that may influence anticipatory reactivity and possibly oxidative stress levels. This study conceptualized resilience in relation to a specific chronic stressor: dementia caregiving; however, psychobiological resilience in relation to a traumatic stressor or the accumulation of various types of stressful life events over time might yield a different pattern of system adaptation or optimization.
The presence of medication use and medical conditions in this sample is a limitation; however, when recruiting older adults, excluding all such factors could lead to a biased sample of highly resilient individuals, hence some statistical controls such as those used herein may represent a preferable alternative. Specifically, more caregivers were taking antidepressants, which have been previously associated with decreases in cortisol (Knorr et al., 2012). Given the direction of that effect, the more likely concern is that it may have contributed to an underestimation of the reported mediation effects; however future studies should consider this factor in the research design. The accumulation of oxidative damage over time could potentially impact trajectories of biological aging; however, this would likely require an extended longitudinal study. We did not statistically correct for multiple measures of oxidative damage because each represents unique effects on different molecules (RNA/DNA/lipids), and the existing literature is insufficient to hypothesize a priori which damage markers are most likely to be affected by stress. We find few relations with 8-OHdG except the positive association with negative affect among caregivers (Table 2), and notably, 8-OHdG was not associated with IsoP (as in a previous study by Watters et al. (2009)) or 8-oxoG. This could be because DNA has more repair mechanisms than RNA (Poulsen et al., 2012), and agents that damage RNA do not necessarily damage DNA and vice versa (Fimognari et al., 2008).
The timing of the stress response is an under-appreciated factor in many studies of stress and health. This study demonstrates the importance of conceptualizing timing at many levels, including the chronicity of stress exposure, psychological determinants of acute neuroendocrine reactivity, and the biological mechanisms that determine how quickly a cascade of molecular actions can occur. It is likely the combination of factors occurring on different time-scales and non-linear relationships that give rise to “moderated mediation” effects, such as those illustrated herein. It is therefore important that psychobiological researchers go beyond these statistical terms to understand what processes give rise to the underlying system behavior described by these statistical models (Sapolsky, 1997; Aschbacher et al., 2012). It is only by gaining this deeper understanding of the processes underlying stress-health associations that the field can utilize this knowledge to propel translational applications. Although this study focused on anticipatory stress arousal, recovery is another equally important dimension of stress system dynamics and allostasis (Karatsoreos and McEwen, 2011; Juster et al., 2012), which should be explored in future work of this nature.
Anticipatory stress arousal may accelerate the accumulation of oxidative damage and biological aging, particularly among chronically stressed individuals. In contrast, a strong stress-arousal response may be associated with a beneficial or resilient biological signature (i.e., reduced oxidative stress), among individuals whose stress was episodic and of lower intensity than the national average. In sum, these data support both the toxic stress and eustress models, in which chronic repeated psychological stress exposure may promote damage and increase risk for disease, whereas manageable stress exposure may promote resilience and resistance to disease (Dienstbier, 1989).
Acknowledgments
We thank Aric Prather, Shamini Jain and the Samueli Institute, Jean Tillie and Wendy Wolfson for their technical and intellectual contributions to the study.
Funding Sources: This research was supported in part by funding from the NIH/NIA grant R01 AG030424-01A2, as well as support for the first author from the NIH/NHLBI grant K23 HL112955 and The Institute for Integrative Health, Baltimore, MD. The CTSI CCRC and the Core Immunology Lab were supported by NIH/NCRR UCSF-CTSI Grant No. UL1 RR024131. The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Footnotes
Contributors: KA wrote the first draft of the paper, and EE, AO, OW, FD and TS provided assistance in editing, framing, literature review and interpretation. KA conducted statistical analyses. EE designed the study and developed the protocol. YS conducted the oxidative stress assays, and wrote key methods portions. All authors contributed to and have approved the final manuscript.
Conflict of Interest: No authors have conflicts of interest to declare.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Akagi S, Nagake Y, Kasahara J, Sarai A, Kihara T, Morimoto H, Yano A, Nakao K, Nanba K, Ichikawa H, Makino H. Significance of 8-hydroxy-2'-deoxyguanosine levels in patients with chronic renal failure. Nephrology. 2003;8:192–195. doi: 10.1046/j.1440-1797.2003.00163.x. [DOI] [PubMed] [Google Scholar]
- Aschbacher K, Adam E, Crofford LJ, Kemeny ME, Demitrack MA, Ben-Zvi A. Linking disease symptoms and subtypes with personalized systems-based phenotypes: A proof of concept study. Brain Behav Immun. 2012;26:1047–1056. doi: 10.1016/j.bbi.2012.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aschbacher K, Kemeny ME. New directions in linking the dynamics of affective and stress-arousal systems. Brain Behav Immun. 2011;25:230–231. doi: 10.1016/j.bbi.2010.10.025. [DOI] [PubMed] [Google Scholar]
- Ballal K, Wilson CR, Harmancey R, Taegtmeyer H. Obesogenic high fat western diet induces oxidative stress and apoptosis in rat heart. Mol Cell Biochem. 2010;344:221–230. doi: 10.1007/s11010-010-0546-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broedbaek K, Siersma V, Henriksen T, Weimann A, Petersen M, Andersen JT, Jimenez-Solem E, Stovgaard ES, Hansen LJ, Henriksen JE, Bonnema SJ, Olivarius Nde F, Poulsen HE. Urinary markers of nucleic acid oxidation and long-term mortality of newly diagnosed type 2 diabetic patients. Diabetes Care. 2011;34:2594–2596. doi: 10.2337/dc11-1620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen S, Janicki-Deverts D. Who's Stressed? Distributions of psychological stress in the United States in probability samples from 1983, 2006, and 2009. J Appl Soc Psychol. 2012;42:1320–1334. [Google Scholar]
- Cohen S, Kamarck T, Mermelstein R. A global measure of perceived stress. J Health Soc Behav. 1983;24:385–396. [PubMed] [Google Scholar]
- Damjanovic AK, Yang Y, Glaser R, Kiecolt-Glaser JK, Nguyen H, Laskowski B, Zou Y, Beversdorf DQ, Weng NP. Accelerated telomere erosion is associated with a declining immune function of caregivers of Alzheimer's disease patients. J Immunol. 2007;179:4249–4254. doi: 10.4049/jimmunol.179.6.4249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhabhar FS. Enhancing versus suppressive effects of stress on immune function: implications for immunoprotection and immunopathology. Ann Ny Acad Sci. 2009;16:300–317. doi: 10.1159/000216188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhabhar FS, Malarkey WB, Neri E, McEwen BS. Stress-induced redistribution of immune cells-From barracks to boulevards to battlefields: A tale of three hormones - Curt Richter Award Winner. Psychoneuroendocrinology. 2012;37:1345–1368. doi: 10.1016/j.psyneuen.2012.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhabhar FS, McEwen BS. Enhancing versus suppressive effects of stress hormones on skin immune function. PNAS. 1999;96:1059–1064. doi: 10.1073/pnas.96.3.1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickerson SS, Kemeny ME. Acute stressors and cortisol responses: a theoretical integration and synthesis of laboratory research. Psychol Bull. 2004;130:355–391. doi: 10.1037/0033-2909.130.3.355. [DOI] [PubMed] [Google Scholar]
- Dienstbier RA. Arousal and physiological toughness: implications for mental and physical health. Psychol Rev. 1989;96:84–100. doi: 10.1037/0033-295x.96.1.84. [DOI] [PubMed] [Google Scholar]
- Drabant EM, Kuo JR, Ramel W, Blechert J, Edge MD, Cooper JR, Goldin PR, Hariri AR, Gross JJ. Experiential, autonomic, and neural responses during threat anticipation vary as a function of threat intensity and neuroticism. NeuroImage. 2011;55:401–410. doi: 10.1016/j.neuroimage.2010.11.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du J, Wang Y, Hunter R, Wei Y, Blumenthal R, Falke C, Khairova R, Zhou R, Yuan P, Machado-Vieira R, McEwen BS, Manji HK. Dynamic regulation of mitochondrial function by glucocorticoids. PNAS. 2009;106:3543–3548. doi: 10.1073/pnas.0812671106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epel ES, Blackburn EH, Lin J, Dhabhar FS, Adler NE, Morrow JD, Cawthon RM. Accelerated telomere shortening in response to life stress. PNAS. 2004;101:17312–17315. doi: 10.1073/pnas.0407162101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epel ES, Lin J, Dhabhar FS, Wolkowitz OM, Puterman E, Karan L, Blackburn EH. Dynamics of telomerase activity in response to acute psychological stress. Brain Behav Immun. 2010;24:531–539. doi: 10.1016/j.bbi.2009.11.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fimognari C, Sestili P, Lenzi M, Bucchini A, Cantelli-Forti G, Hrelia P. RNA as a new target for toxic and protective agents. Mutat Res. 2008;648:15–22. doi: 10.1016/j.mrfmmm.2008.09.003. [DOI] [PubMed] [Google Scholar]
- Forlenza MJ, Miller GE. Increased serum levels of 8-hydroxy-2'-deoxyguanosine in clinical depression. Psychosom Med. 2006;68:1–7. doi: 10.1097/01.psy.0000195780.37277.2a. [DOI] [PubMed] [Google Scholar]
- Furukawa S, Fujita T, Shimabukuro M, Iwaki M, Yamada Y, Nakajima Y, Nakayama O, Makishima M, Matsuda M, Shimomura I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J Clin Invest. 2004;114:1752–1761. doi: 10.1172/JCI21625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaab J, Rohleder N, Nater UM, Ehlert U. Psychological determinants of the cortisol stress response: the role of anticipatory cognitive appraisal. Psychoneuroendocrinology. 2005;30:599–610. doi: 10.1016/j.psyneuen.2005.02.001. [DOI] [PubMed] [Google Scholar]
- Gidron Y, Russ K, Tissarchondou H, Warner J. The relation between psychological factors and DNA-damage: a critical review. Biol Psychol. 2006;72:291–304. doi: 10.1016/j.biopsycho.2005.11.011. [DOI] [PubMed] [Google Scholar]
- Gouin JP, Hantsoo L, Kiecolt-Glaser JK. Immune dysregulation and chronic stress among older adults: a review. Ann Ny Acad Sci. 2008;15:251–259. doi: 10.1159/000156468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutierrez J, Ballinger SW, Darley-Usmar VM, Landar A. Free radicals, mitochondria, and oxidized lipids: the emerging role in signal transduction in vascular cells. Circ Res. 2006;99:924–932. doi: 10.1161/01.RES.0000248212.86638.e9. [DOI] [PubMed] [Google Scholar]
- Harman D. Aging: a theory based on free radical and radiation chemistry. J Gerontol. 1956;11:298–300. doi: 10.1093/geronj/11.3.298. [DOI] [PubMed] [Google Scholar]
- Hayes AF. Beyond Baron and Kenny: Statistical Mediation Analysis in the New Millennium. Commun Monogr. 2009;76:408–420. [Google Scholar]
- Hayes AF. PROCESS: A versatile computational tool for observed variable mediation, moderation, and conditional process modeling [White paper] 2012 Retrieved from http://www.afhayes.com/public/process2012.pdf.
- Humphreys J, Epel ES, Cooper BA, Lin J, Blackburn EH, Lee KA. Telomere shortening in formerly abused and never abused women. Biol Res Nurs. 2012;14:115–123. doi: 10.1177/1099800411398479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Irie M, Asami S, Ikeda M, Kasai H. Depressive state relates to female oxidative DNA damage via neutrophil activation. Biochem Biophys Res Commun. 2003;311:1014–1018. doi: 10.1016/j.bbrc.2003.10.105. [DOI] [PubMed] [Google Scholar]
- Joergensen A, Broedbaek K, Weimann A, Semba RD, Ferrucci L, Joergensen MB, Poulsen HE. Association between urinary excretion of cortisol and markers of oxidatively damaged DNA and RNA in humans. PLoS ONE. 2011;6:e20795. doi: 10.1371/journal.pone.0020795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juster RP, Perna A, Marin MF, Sindi S, Lupien SJ. Timing is everything: anticipatory stress dynamics among cortisol and blood pressure reactivity and recovery in healthy adults. Stress. 2012;15:569–577. doi: 10.3109/10253890.2012.661494. [DOI] [PubMed] [Google Scholar]
- Karatsoreos IN, McEwen BS. Psychobiological allostasis: resistance, resilience and vulnerability. Trends Cogn Sci. 2011;15:576–584. doi: 10.1016/j.tics.2011.10.005. [DOI] [PubMed] [Google Scholar]
- Kirschbaum C, Pirke KM, Hellhammer DH. The ‘Trier Social Stress Test’--a tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology. 1993;28:76–81. doi: 10.1159/000119004. [DOI] [PubMed] [Google Scholar]
- Kirschbaum C, Prussner JC, Stone AA, Federenko I, Gaab J, Lintz D, Schommer N, Hellhammer DH. Persistent high cortisol responses to repeated psychological stress in a subpopulation of healthy men. Psychosom Med. 1995;57:468–474. doi: 10.1097/00006842-199509000-00009. [DOI] [PubMed] [Google Scholar]
- Kitano H. Towards a theory of biological robustness. Mol Syst Biol. 2007;3:1–7. doi: 10.1038/msb4100179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knorr U, Vinberg M, Gether U, Winkel P, Gluud C, Wetterslev J, Kessing LV. The effect of escitalopram versus placebo on perceived stress and salivary cortisol in healthy first-degree relatives of patients with depression-A randomised trial. Psychiatry Res. 2012;200:354–360. doi: 10.1016/j.psychres.2012.05.015. [DOI] [PubMed] [Google Scholar]
- Mackinnon DP, Fairchild AJ. Current Directions in Mediation Analysis. Curr Dir Psychol Sci. 2009;18:16. doi: 10.1111/j.1467-8721.2009.01598.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McEwen BS. Protection and damage from acute and chronic stress: allostasis and allostatic overload and relevance to the pathophysiology of psychiatric disorders. Ann N Y Acad Sci. 2004;1032:1–7. doi: 10.1196/annals.1314.001. [DOI] [PubMed] [Google Scholar]
- Miklos IH, Kovacs KJ. Reorganization of synaptic inputs to the hypothalamic paraventricular nucleus during chronic psychogenic stress in rats. Biol Psychiatry. 2012;71:301–308. doi: 10.1016/j.biopsych.2011.10.027. [DOI] [PubMed] [Google Scholar]
- Milne GL, Sanchez SC, Musiek ES, Morrow JD. Quantification of F2-isoprostanes as a biomarker of oxidative stress. Nat Protoc. 2007;2:221–226. doi: 10.1038/nprot.2006.375. [DOI] [PubMed] [Google Scholar]
- Monnier L, Mas E, Ginet C, Michel F, Villon L, Cristol JP, Colette C. Activation of oxidative stress by acute glucose fluctuations compared with sustained chronic hyperglycemia in patients with type 2 diabetes. JAMA. 2006;295:1681–1687. doi: 10.1001/jama.295.14.1681. [DOI] [PubMed] [Google Scholar]
- Muller FL, Lustgarten MS, Jang Y, Richardson A, Van Remmen H. Trends in oxidative aging theories. Free Radic Biol Med. 2007;43:477–503. doi: 10.1016/j.freeradbiomed.2007.03.034. [DOI] [PubMed] [Google Scholar]
- Nunomura A, Tamaoki T, Motohashi N, Nakamura M, McKeel DW, Jr, Tabaton M, Lee HG, Smith MA, Perry G, Zhu X. The earliest stage of cognitive impairment in transition from normal aging to Alzheimer disease is marked by prominent RNA oxidation in vulnerable neurons. J Neuropathol Exp Neurol. 2012;71:233–241. doi: 10.1097/NEN.0b013e318248e614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Donovan A, Tomiyama AJ, Lin J, Puterman E, Adler NE, Kemeny M, Wolkowitz OM, Blackburn EH, Epel ES. Stress appraisals and cellular aging: a key role for anticipatory threat in the relationship between psychological stress and telomere length. Brain Behav Immun. 2012;26:573–579. doi: 10.1016/j.bbi.2012.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poulsen HE, Specht E, Broedbaek K, Henriksen T, Ellervik C, Mandrup-Poulsen T, Tonnesen M, Nielsen PE, Andersen HU, Weimann A. RNA modifications by oxidation: a novel disease mechanism? Free Radic Biol Med. 2012;52:1353–1361. doi: 10.1016/j.freeradbiomed.2012.01.009. [DOI] [PubMed] [Google Scholar]
- Radak Z, Chung HY, Goto S. Exercise and hormesis: oxidative stress-related adaptation for successful aging. Biogerontology. 2005;6:71–75. doi: 10.1007/s10522-004-7386-7. [DOI] [PubMed] [Google Scholar]
- Sapolsky RM. McEwen-Induced Modulation of Endocrine History: A Partial Review. Stress. 1997;2:1–12. doi: 10.3109/10253899709014733. [DOI] [PubMed] [Google Scholar]
- Sapolsky RM, Romero LM, Munck AU. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev. 2000;21:55–89. doi: 10.1210/edrv.21.1.0389. [DOI] [PubMed] [Google Scholar]
- Tomiyama AJ, O'Donovan A, Lin J, Puterman E, Lazaro A, Chan J, Dhabhar FS, Wolkowitz O, Kirschbaum C, Blackburn E, Epel E. Does cellular aging relate to patterns of allostasis? An examination of basal and stress reactive HPA axis activity and telomere length. Physiol Behav. 2012;106:40–45. doi: 10.1016/j.physbeh.2011.11.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem Biol Interact. 2006;160:1–40. doi: 10.1016/j.cbi.2005.12.009. [DOI] [PubMed] [Google Scholar]
- Watson D, Clark LA, Tellegen A. Development and validation of brief measures of positive and negative affect: the PANAS scales. J Pers Soc Psychol. 1988;54:1063–1070. doi: 10.1037//0022-3514.54.6.1063. [DOI] [PubMed] [Google Scholar]
- Watters JL, Satia JA, da Costa KA, Boysen G, Collins LB, Morrow JD, Milne GL, Swenberg JA. Comparison of three oxidative stress biomarkers in a sample of healthy adults. Biomarkers. 2009;14:587–595. doi: 10.3109/13547500903183954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeakley JM, Balasubramanian K, Harrison RW. Comparison of glucocorticoid-receptor binding kinetics with predictions from a biomolecular model. J Biol Chem. 1980;255:4182–4188. [PubMed] [Google Scholar]
- Yerkes RM, Dodson JD. The relation of strength of stimulus to rapidity of habit-formation. J Comp Neurol Psychol. 1908;18:459–482. [Google Scholar]