

Abstract
Zinc finger proteins that bind Zn(II) using a Cys2His2 coordination motif within a ββα protein fold are the most abundant DNA binding transcription factor domains in eukaryotic systems. These classic zinc fingers are typically unfolded in the apo state and spontaneously fold into their functional ββα folds upon incorporation of Zn(II). These metal-induced protein folding events obscure the free energy cost of protein folding by coupling the protein folding and metal-ion binding thermodynamics. Herein, we determine the formation constant of a Cys2His2/ββα zinc finger domain, the C-terminal finger of the Wilms’ tumor suppressor protein (WT1-4), for the purposes of determining its free energy cost of protein folding. Measurements of individual conditional dissociation constants, Kd values, at pH values from 5 to 9 were determined using fluorescence spectroscopy by direct or competition titration. Potentiometric titrations of apo-WT1-4 followed by NMR spectroscopy provided the intrinsic pKa values of the Cys2His2 residues, and corresponding potentiometric titrations of Zn(II)–WT1-4 followed by fluorescence spectroscopy yielded the effective pKaeff values of the Cys2His2 ligands bound to Zn(II). The Kd, pKa, and pKaeff values were combined in a minimal, complete equilibrium model to yield the pH-independent formation constant value for Zn(II)–WT1-4, KfML value of 7.5 × 1012 M–1, with a limiting Kd value of 133 fM. This shows that Zn(II) binding to the Cys2His2 site in WT1-4 provides at least −17.6 kcal/mol in driving force to fold the protein scaffold. A comparison of the conditional dissociation constants of Zn(II)–WT1-4 to those from the model peptide Zn(II)–GGG–Cys2His2 over the pH range 5.0 to 9.0 and a comparison of their pH-independent KfML values demonstrates that the free energy cost of protein folding in WT1-4 is less than +2.1 kcal/mol. These results validate our GGG model system for determining the cost of protein folding in natural zinc finger proteins and support the conclusion that the cost of protein folding in most zinc finger proteins is ≤+4.2 kcal/mol, a value that pales in comparison to the free energy contribution of Zn(II) binding, −17.6 kcal/mol.
Short abstract
The formation constant for a natural Cys2His2 ββα zinc finger peptide has been determined using a suite of equilibrium measurements. These data are compared to literature values for Zn(II)−Cys2His2 protein events to assess the free energy cost of protein folding in proteins with metal-induced protein folding events. The data lead to the conclusion that the free energy contribution of Zn(II) binding is far in excess of the minimal free energy cost of protein folding.
Introduction
Zinc finger transcription factors are one of the largest classes of eukaryotic proteins and are characteristic examples of structural Zn(II) proteins.1 While classically recognized for their DNA/RNA binding ability,2 zinc fingers are also involved in mediating protein–protein interactions3 and membrane association4 via lipid binding and may act as biological redox switches.5 One key structure–function relationship in zinc fingers is their metal-induced protein folding events.6 Zinc fingers are typically unstructured in the apo state, and they fold into their biologically active ββα fold upon Zn(II) incorporation. The Zn(II) is bound tightly, sub-nanomolar Kd values at physiological pH, using pseudotetrahedral Cys2His2, Cys3His1, or Cys4 binding sites.7,8 Because Zn(II) binding and protein folding are thermodynamically coupled in the metal-induced protein folding event, the precise free energy contribution of Zn(II) binding toward protein structure, assembly, and function is obscured by the underlying free energy cost of protein folding. Indeed, estimates of the free energy cost of protein folding in zinc fingers in the literature are as high as +16 kcal/mol.9
We have developed a method to separate the free energies of metal-ion binding and protein folding in zinc finger proteins.10 Our method is based on a designed 16 amino acid peptide scaffold, GGG, into which we individually incorporated each of the classic zinc finger metal-ion binding motifs, Cys2His2, Cys3His1, and Cys4. The resulting three peptides, GGG–Cys2His2, GGG–Cys3His1, and GGG–Cys4, each bind Zn(II) in a 1:1 stoichiometry in a pseudotetrahedral coordination motif with tight affinity, sub-nanomolar Kd values at physiological pH.10,11 A suite of detailed equilibrium measurements over the pH range of 4–9 and a complete description of the metal–peptide binding equilibrium were used to measure their formation constant, KfML (or β110) values of 2.5 × 1013, 1.5 × 1015, and 5.6 × 1016 M–1 for GGG–Cys2His2, GGG–Cys3His1, and GGG–Cys4, respectively.9 The KfML values of GGG–Cys2His2, GGG–Cys3His1, and GGG–Cys4 indicate that cysteine thiolates are better ligands than histidine imidazoles12,13 and that Zn(II) binding is favorable by 18.3, 20.7, and 22.8 kcal/mol, respectively. Additionally, these KfML values are attenuated by the ligand pKa values such that the Kd values at physiological pH are nearly identical, i.e. 1–5 pM. Because the GGG peptides have no secondary structure in the apo and holo forms, their free energy cost of protein folding is minimal and, correspondingly, their Zn(II) binding affinities, KfML values, are tighter than natural zinc finger proteins with the same coordination motif. We10 and others14 have compared the free energies of Zn(II) binding to the GGG peptide with the free energy of Zn(II) binding to natural and synthetic zinc finger peptides and proteins with the same coordination motif to deduce the free energy cost of protein folding in the latter at specific pH values using conditional dissociation constant values. These data indicate that the free energy cost of protein folding in most zinc fingers is between 0–4 kcal/mol.10 However, a recent report by Sénèque and Latour,15 which reports substantially tighter Zn(II) affinity in the Consensus Peptide 1 series of designed zinc fingers and slow metal-ion exchange kinetics, indirectly questions this key result.
To date, our conclusions have been based on comparisons of conditional dissociation constants between our GGG model peptide and zinc fingers at individual pH values because of a lack of KfML values for natural zinc fingers. Herein, we explore the effect of pH on the free energy cost of protein folding using a natural Cys2His2 zinc finger, the C-terminal finger of the Wilms’ tumor suppressor protein,16WT1-4. We have determined the formation constant for Zn(II)–WT1-4 to be 7.5 × 1012 M–1 using conditional dissociation constant measurements, EGTA and HEDTA competition constant measurements, and potentiometric titrations of the apo and holo forms. A comparison of the conditional dissociation constants of Zn(II)–WT1-4 to those from Zn(II)–GGG–Cys2His2 over the pH range 5.0 to 9.0 and a comparison of the pH-independent KfML values demonstrate that the free energy cost of protein folding in both is identical within error. These results validate our model system and indicate that the pH effects on the cost of protein folding in peptides with an unstructured apo form are similar.
Experimental Section
Materials
Zinc(II) chloride, trifluoroacetic acid, ethanedithiol, 1-hydroxybenzotriazole, diethyl ether, acetic anhydride, diisopropylethylamine (DIEA), and piperidine were obtained from the Sigma-Aldrich Chemical Co. Aqueous stock solutions of Zn(II) were quantified by atomic absorption spectroscopy. Natural Fmoc-protected amino acids were obtained from Bachem. HBTU, O-(1H-benzotriazole-1-yl)-N,N,N′,N′-tetramethyluronium hexafluorophosphate, was purchased from Qbiogene. All other chemicals and solvents were reagent grade and used without further purification.
Chemical Synthesis of the Peptide
The 27 amino acid C-terminal zinc finger peptide of the Wilms tumor suppressor protein17 (WT1-4) was synthesized using sold-phase peptide synthesis.18 The crude peptide was purified to homogeneity with HPLC, and the identity of the purified peptide was confirmed with mass spectrometry. The sequence is given below with the zinc-binding residues highlighted in bold and the tryptophan residue in italics:
UV–vis Spectroscopy
UV–visible spectra were recorded on either a Varian Cary 100 or a Cary 300 spectrophotometer using anaerobic quartz cells of 1.0 cm path length. Peptide concentrations were determined spectrophotometrically using ε280 of 5600 M–1 cm–1 for Trp.18
Circular Dichroism Spectropolarimetry
CD spectra were recorded on a Aviv Biomedical model 214 CD spectrometer using anaerobic quartz cells of 1.0 cm path length. Peptide concentrations were determined spectrophotometrically using ε280 of 5600 M–1 cm–1 for Trp.18
Fluorescence Spectroscopy
Excitation and emission fluorescence spectra were recorded on a Cary Eclipse fluorimeter using anaerobic quartz cells of 1.0 cm path length. Excitation and emission slit widths of 5.0 and 2.5 nm, respectively, were employed. The excitation wavelength was 280 nm, and the fluorescence emission was collected from 300 to 450 nm. Peptide concentrations were between 1 and 25 μM, as determined spectrophotometrically using ε280 = 5600 M–1 cm–1 for Trp.18
NMR Spectroscopy
A 1.2 mM sample of apo-WT1-4 in water was prepared in an anaerobic glovebox and placed into a NMR tube equipped with a J. Young valve for spectral measurement. The 600 μL sample of apo-WT1-4 contained 10 mM KCl, 0.1 mM DSS, and 10% D2O. The pH was set using microliter aliquots of 0.1 M HNO3 or 0.1 NaOH under strictly anaerobic conditions to prevent cysteine oxidation.
All 1D and 2D NMR spectra of the apo-WT1-4 peptide were collected on a Bruker DRX600 NMR spectrometer at 288 K. Water suppression in all 2D experiments was accomplished with the double gradient echo methods of Hwang and Shaka.19 The data were processed using NMRPipe20 and extended using linear prediction and zero filling. The data were analyzed using NMRView.21 All proton resonances for apo-WT1-4 were assigned using the methods developed by Wüthrich.22 Intraresidue proton resonances were assigned using both short (15 ms) and long (80 ms) mixing time TOCSY experiments and a 225 ms mixing time NOESY experiment at 288 K using 640 t1 increments at pH 6.0. The chemical shift values of the δ and ε protons of His23, His24, and His27 as well as the β protons of Cys5 and Cys10 were then recorded from long mixing time (80 ms) TOCSY spectra collected at 17 different pH values under strictly anaerobic conditions.
X-ray Absorption Spectroscopy
Samples of Zn(II)–WT1-4 (∼1–2 mM) were prepared with 20% (v/v) glycerol and loaded in Lucite cuvettes with 6 μm polypropylene windows before rapid freezing in liquid nitrogen. X-ray absorption spectra were measured at the National Synchrotron Light Source (Brookhaven National Lab, Upton, NY), beamline X3B, with a Si(111) double crystal monochromator; harmonic rejection was accomplished using a Ni focusing mirror. Data collection and reduction were accomplished according to published procedures.23 The data presented for Zn(II)–WT1-4 represent the average of the six scans.
Both raw and Fourier filtered EXAFS data were fit utilizing theoretical amplitude and phase functions calculated with FEFF ver. 8.00.24 The Zn–N and Zn–S scale factors and the threshold energy, ΔE0, were calibrated to the experimental spectra of the tetrakis-1-methylimidazole and tetraphenylthiolate complexes of Zn. These calibrated values (SZn–N = 0.78, SZn–S = 0.85, and DE0 = −16 eV, with E0 set to 9675 eV) were held fixed in subsequent fits to Zn(II)–WT1-4 data. First shell fits were then obtained for all reasonable coordination numbers while allowing the absorber–scatterer distance, Ras, and the Debye–Waller factor, σas2, to vary. Fits to unfiltered EXAFS gave identical results. In no case did inclusion of a mixed first shell, with distinct Zn–N and Zn–O scattering contributions, result in either a significant improvement in fit residual or resolvable Zn–N/Zn–O distances. Multiple scattering contributions from coordinated histidines were fit using a set of combined multiple-scattering paths, according to published procedures.25
Isothermal Titration Fluorimetry: Direct Metal-Ion Titrations
Aqueous stock solutions of Zn(II)Cl2 unbuffered at pH 7.0 were added in microliter aliquots to freshly prepared WT1-4 peptide solutions in aqueous buffers under strictly anaerobic conditions in 1.0 cm cuvettes. Samples were allowed to equilibrate for at least 10 min (at least twice their measured equilibration time) before measurement of their fluorescence spectra. The conditional metal–ligand dissociation constants, conditional Kd values, were obtained from fitting a plot of the increase in tryptophan fluorescence at 355 nm against the [Zn(II)]/[WT1-4] ratio to the following 1:1 equilibrium binding model.
| 1 |
| 2 |
The equation used to fit the data is as follows
| 3 |
where Flmeas, the measured fluorescence emission intensity, is a function of Fl0, the fluorescence intensity of the WT1-4 peptide ligand prior to metal binding, Fllim, the limiting emission intensity of the Zn(II)–WT1-4 complex, MT, the total concentration of metal added to peptide solution, LT, the total concentration of the WT1-4 peptide, and Kd, the conditional dissociation constant.
Isothermal Titration Fluorimetry: EGTA and HEDTA Competition Titrations
For pH values above 6.0, conditional equilibrium dissociation constant determination for the Zn(II)–WT1-4 complex necessitated the use of EGTA (ethylene glycol tetraacetic acid) and HEDTA (N-(2-hydroxyethyl)ethylenediamine-N,N′,N′-triacetic acid) competition.26,27 An unbuffered aqueous solution of Zn(II)Cl2 at pH 7.0 was added in microliter aliquots to a buffered solution of 22.1 μM WT1-4 containing 5.0 equiv of competitor (EGTA or HEDTA) under strictly anaerobic conditions. The increase in fluorescence at 355 nm upon the addition of Zn(II) was fit to a competition equilibrium binding model based on eqs 4–8.
| 4 |
| 5 |
| 6 |
| 7 |
| 8 |
Where Flmeas, the measured fluorescence emission intensity, is a function of Fl0, the fluorescence of the WT1-4 peptide prior to metal binding, Fllim, the limiting fluorescence of the Zn(II)–WT1-4 complex, MT, the total concentration of metal added to peptide solution, LT, the total concentration of the WT1-4 ligand, EGTAT, the total concentration of EGTA, and Kcomp, the conditional competition constant.
The Kcomp value, coupled with the conditional equilibrium dissociation constant value of Zn(II)–EGTA, KdZn(II)–EGTA, given by eqs 9–11, gives the conditional equilibrium dissociation constant value for Zn(II)–WT1-4.
| 9 |
| 10 |
| 11 |
Where αL is the mole fraction of fully deprotonated EGTA, KfZn(II)–EGTA is the formation constant of fully deprotonated EGTA for Zn(II), a value of 1013.1, and K(1–4) are the stepwise proton dissociation constants of EGTA; K1 = 1.0, K2 = 3.1 × 10–2, K3 = 1.0 × 10–2, and K4 = 2.2 × 10–3.27
Potentiometric pH Titrations. Apo Peptide
The NMR spectra of WT1-4 support an unfolded apo state because very few interresidues NOEs were observed and chemical shift values of the Ha protons were very close to their random coil values.22,28 The pKa values for the metal-ion ligands in apo-WT1-4 were determined from fits of the graphs of chemical shift value versus pH for each of the Cys and His residues using the equation
| 12 |
where the measured proton chemical shift, δmeas, is a function of protonated amino acid chemical shift, δ0, the change in chemical shift due to deprotonation, Δδ, the solution pH value, and the acid dissociation constant of the amino acid pKa. These four pKa1–4 values establish the speciation of the apo-WT1-4, as shown in Scheme 1.
Scheme 1. Equilibrium Model for Zn(II)–WT1-4 Formation.

Potentiometric pH Titrations. Holo Peptide
Potentiometric pH titrations of Zn(II)–WT1-4 were performed manually using a 1.0 cm path length fluorescence cuvette fitted with a pH electrode under a stream of hydrated nitrogen gas. The pH of a 17 μM Zn(II)–WT1-4 sample in unbuffered water at pH 10 was adjusted by addition of microliter aliquots of 0.1 M HCl. Between each addition, the samples were allowed to equilibrate 3 min prior to measurement of their fluorescence emission spectra. The pH dependence of the fluorescence emission at 355 nm was fit to an equation for two separate protonation events, a cooperative two-proton event with an effective pKa1,2eff value and a cooperative two-proton event with an effective pKa3,4eff value that establishes the speciation of Zn(II)–WT1-4 given in Scheme 1.
| 13 |
Where the fluorescence emission at 355 nm measured at any pH, Flmeas is a function of the initial fluorescence, and Fl0, is the change in fluorescence due to the first and second protonation events, ΔFl1 and ΔFl2, respectively, the solution pH value, and the effective acid dissociation constants of the ligands bound to metal, pKa1,2eff and pKa3,4eff. For Zn(II)–WT1-4, the transition reflected by pKa3,4eff represented ∼65% of the total fluorescence intensity change, with the remainder represented by the pKa1,2eff-based transition.
pH Dependence of Conditional Dissociation Constants
Because of the expected proton dependence of the Zn(II)–WT1-4 conditional dissociation constants, Kd values were measured at varying pH values in order to determine the value of KfML for each metal. The Kd values of Zn(II)–WT1-4 at each pH were determined as above using fluorescence spectroscopy. The resulting plots of −log Kd versus pH is fit to the following equilibrium binding expression for the pH-dependent formation of Zn(II)–WT1-4 from the WT1-4 peptide and {Zn(II)(H2O)6}2+ using the proton binding model in Scheme 1:
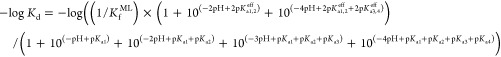 |
14 |
The conditional dissociation constant at any pH, Kd, is a function of the pH-independent formation constant at high pH, KfML, the effective acid dissociation constants of the metal-bound histidines, pKa1,2eff, and cysteines, pKa3,4eff, the acid dissociation constant values for the histidines, pKa1 and pKa2, and cysteines, pKa3 and pKa4, in the apo peptide, and the solution pH.
Results
Experimental Design
Zinc(II) plays a major role in biochemistry via its ability to stabilize protein structure.29−36 The largest class of metalloproteins encoded in the human genome,37,38 the zinc finger proteins, are typically unstructured in the absence of Zn(II) and fold into their biologically active forms upon Zn(II) incorporation.6 Figure 1 shows the C-terminal zinc finger of the Wilms’ tumor suppressor protein (WT1-4), which folds into the classic zinc finger ββα fold upon binding Zn(II) to its Cys2His2 site.17 Indeed, zinc finger proteins are the prototypical examples of metal-induced protein folding events. Because the free energy of metal-ion binding is used to overcome the unfavorable free energy of protein folding, its actual value is obscured in typical measurements of metal–protein association/dissociation constants.
Figure 1.

(Left) Molecular model of the Zn(II)–WT1-4 complex rendered in PyMOL68 and (Right) free energy diagram of zinc finger protein folding.
In zinc finger proteins (ZFPs) such as WT1-4, the free energy of Zn(II) binding to the Cys2His2 site is coupled to the free energy of protein folding.39 Figure 1 shows that the apo-folded state is higher in energy than the apo-unfolded state of a typical zinc finger, with the difference equal to the free energy of apo-protein folding, ΔGapofolding. There is no direct method to measure the value of ΔGapofolding, which has been estimated to be between 0 and +16 kcal/mol.9 The free energy contribution of metal-ion binding, ΔGML, is the energy difference between the apo-folded and holo-folded states; the structure15 is shown for WT1-4 in Figure 1. ΔGML cannot typically be measured because of the uncertainty in the value of ΔGapofolding. The observed free energy contribution of metal-ion binding, ΔGML–obs, can be directly measured and is weaker than the free energy contribution of metal-ion binding, ΔGML, by the free energy cost of protein folding, ΔGapofolding, as follows:
| 15 |
We have developed a method to tease apart the free energy of metal-ion binding, ΔGML, from the free energy of protein folding, ΔGapofolding, in zinc finger proteins.10 Our method is based on a designed 16 amino acid peptide scaffold, GGG, into which we individually incorporated each of the classic zinc finger metal-ion binding motifs, Cys2His2, Cys3His1, and Cys4. The resulting three peptides, GGG–Cys2His2, GGG–Cys3His1, and GGG–Cys4, each bind Zn(II) in a 1:1 stoichiometry in a pseudotetrahedral coordination motif and have formation constant values of 2.5 × 1013, 1.5 × 1015, and 5.6 × 1016 M–1, respectively, as determined using a suite of detailed equilibrium measurements over the pH range of 5–9.10,11 Because the GGG peptides have no secondary structure in the apo and holo forms, their free energy cost of protein folding is minimal, ΔGapofolding = 0 kcal/mol, and their observed and actual free energy contributions of Zn(II) binding are equivalent. In natural ZFPs, the observed free energy of Zn(II) binding is weaker than the actual free energy because of the positive values of ΔGapofolding. Therefore, the difference in the actual free energy of Zn(II) binding to GGG to the observed free energy of Zn(II) binding to a ZFP can be used to reveal the cost of protein folding in the ZFP. We and others14 have compared the free energy of Zn(II) binding to the GGG peptide to natural and synthetic zinc finger peptides and proteins with the same coordination motif, derived from conditional dissociation constant values in the literature, to the free energy of Zn(II) binding to the GGG peptide with the same Zn(II) binding motif to determine the free energy cost of protein folding in the former.
| 16 |
These comparisons indicate that the free energy cost of protein folding in most zinc fingers is between 0 and +4 kcal/mol, and it does not appear to vary between the classic coordination motifs of Cys2His2, Cys3His1, and Cys4 despite the fact that the KfML values for each motif are distinct.
These conclusions are based on the comparison of the GGG peptide data to natural and synthetic zinc finger dissociation constants at single pH values. Although this has provided considerable insight into the free energy cost of protein folding in zinc fingers, the pH dependence of the conditional dissociation constant values is significant, and a comparison of the pH-independent formation constants, KfML values, is more accurate because it removes effects resulting from changes in the His/Cys pKa values. Because of the lack of KfML values for natural zinc fingers, a comparison of KfML values cannot be done at this time; in addition, any observed changes in conditional dissociation constants may reflect changes in KfML values or the inherent His/Cys pKa values. The closest comparison that can be done is for the Cys3His1 motif based on amino acids 34–51 of the HIV-1 nucleocapsid protein, (34–51)NCp7.40,41 The reported KfML value for (34–51)NCp7 is 2 × 1015 M–1, which is identical, within error, to the 1.5 × 1015 M–1 value reported for GGG–Cys3His1, indicating a similar cost of protein folding between these two peptides. However, any conclusions about zinc finger protein folding are somewhat speculative because the Zn knuckle fold of NCp7 is distinct from the classic zinc finger ββα fold. Additionally, the observed similarities in KfML values between GGG and (34–51)NCp7 may reflect their size similarity, 16 versus 18 amino acids, or structural similarity, lack of helical/sheet structure in the two holo peptides.
Because the literature KfML values for natural zinc fingers do not exist, we chose to evaluate a classic zinc finger with Cys2His2 coordination that adopts the classic ββα fold for this contribution. A single finger domain from the full-length four finger Wilms’ tumor suppressor protein (WT1) was selected, as the full-length protein required for kidney and genitourinary system development42−45 has been structurally characterized by both NMR and X-ray crystallography.17 The single C-terminal Cys2His2/ββα finger zinc finger, WT1-4, was selected for study because it is the only one with the intrinsic fluorescence probe tryptophan in its wild-type sequence. Herein, we present the first intrinsic pH-independent formation constant determination of a natural zinc finger protein with the classic ββα fold. The conditional dissociation constants and overall formation constant for Zn(II)–WT1-4 are determined using a suite of equilibrium methods that include the measurement of the pKa and effective pKa values of the His/Cys ligands, which provide a complete description of the metal–peptide binding equilibrium. These data are used to show that the cost of protein folding in WT1-4 over the pH range of 5–9 is minimal. These data validate our method for determining and evaluating the cost of protein folding across a wide pH range in a natural protein scaffold.
Fluorescence Spectroscopy
Figure 2 shows the steady-state fluorescence emission spectrum of WT1-4 and Zn(II)–WT1-4 at 22 μM concentration in aqueous buffer. The fluorescence emission maximum of Trp7 in apo-WT1-4 at 355 nm indicates that the indole ring is solvent-exposed.46 Zn(II) binding to WT1-4 results in a slight blue-shift of the emission maximum to 350 nm, along with an increase in tryptophan fluorescence emission intensity, which is due, in part, to the elimination of the quenching mechanism of cysteine thiols.47 The observed hypsochromic shift indicates that Trp7 in the metal-bound WT1-4 peptide is slightly protected from solvent, as evidenced in the NMR and X-ray structures of WT1, whereas Trp7 in apo-WT1-4 is more solvent-exposed.17 These data are consistent with an unfolded apo-WT1-4 that folds upon Zn(II) incorporation, and they provide a spectroscopic indicator of metal-ion binding.
Figure 2.

Steady-state fluorescence emission spectra of 22 μM apo-WT1-4 (dotted line) and 22 μM Zn(II)–WT1-4 (solid line) in pH 7.0 buffer (20 mM HEPES, 100 mM KCl). Each sample was excited through a 5 nm slit at 280 nm, the tryptophan λmax value, and the fluorescence emission was collected through a 2.5 nm slit.
Isothermal Titration Fluorimetry
The intrinsic fluorescence of Trp7 was used to follow the incorporation of Zn(II) under a variety of solution pH conditions. The conditional dissociation constants for Zn(II)–WT1-4 were measured over the pH range of 5.0 to 9.0 using fluorescence spectroscopy because of the pH dependence on the Kd values resulting from proton release upon metal-ion binding to apo-WT1-4.
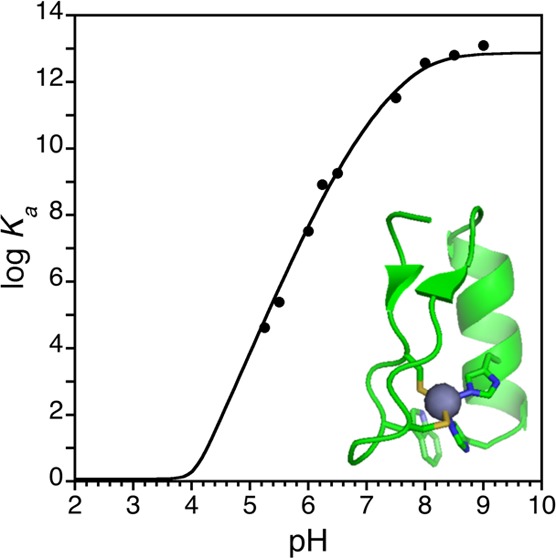
Figure 3 shows the direct titration of Zn(II) into 22 μM WT1-4 at pH 5.25 (20 mM MES, 100 mM KCl). The equilibrium binding isotherm shown in Figure 3 is fit to a 1:1 metal–peptide binding model, eq 3, and demonstrates a conditional dissociation constant value of 22 μM at pH 5.25. At pH values above 6.0, competition titrations with the chelators HEDTA and EGTA were performed to obtain accurate conditional dissociation constant values for Zn(II)–WT1-4. The formation constants for Zn(II)–HEDTA and Zn(II)–EGTA and their respective pKa values were used to calculate their conditional dissociation constants between pH 6.0 and 9.0. Figure 4 shows a typical competition titration of Zn(II) into a buffered pH 9.0 (20 mM Tris, 100 mM KCl) solution containing 22 μM WT1-4 and 110 μM HEDTA. The equilibrium binding isotherm shown in Figure 4 is fit to a competition constant of 3.31. Because the Kd of Zn(II)–HEDTA is 2.01 × 10–14 M at pH 9.0, the conditional dissociation constant of Zn(II)–WT1-4 is 6.6 × 10–14 M at pH 9.0. Kinetic experiments were performed to ensure that samples had reached equilibrium. Figure 5 shows the change in fluorescence intensity as a function of time upon addition of 5 equiv of EDTA to 22 μM Zn(II)–WT1-4 in 20 mM MES, 100 mM KCl, at pH 6.65. The initial intensity, 145.67 au, decreases to 78.45 au in 15 min and remains constant for the following 45 min. Kinetic experiments from direct and competition titrations shows that all samples were equilibrated within 15 min, so measurements were taken 30 min after Zn(II) addition to ensure equilibration.
Figure 3.

Direct titration of Zn(II)Cl2 in unbuffered aqueous solution at pH 7.0 into 22 μM WT1-4 buffered at pH 5.25 (20 mM HEPES, 100 mM KCl) followed by fluorescence spectroscopy. Spectra are shown for the addition of 0.00, 0.22, 0.67, 0.99, 1.81, 2.71, 4.07, 6.78, 8.59, and 12.20 equiv of Zn(II) added, with the others were omitted for clarity. The increase in emission intensity at 355 nm observed upon Zn(II) binding is fit in the inset to a Zn(II)–WT1-4 conditional dissociation constant, Kd, value of 9.9 μM at pH 25.
Figure 4.

Competition titration of Zn(II)Cl2 in unbuffered aqueous solution at pH 7.0 into an aqueous solution containing 22 μM WT1-4 and 110 μM HEDTA buffered at pH 9.0 (20 mM Tris, 100 mM KCl) followed by fluorescence spectroscopy. Spectra are shown for the addition of 0.00, 0.90, 2.03, 2.94, and 4.29 equiv of Zn(II) added, with the others omitted for clarity. Under these conditions, a fit to the plot of fluorescence at 357 nm vs equivalents of Zn(II) added to peptide using eq 6 gives a competition constant value of 3.31 between WT1-4 and HEDTA. Because the Kd of Zn(II)–HEDTA at pH 9.0 is 20.1 fM, the resulting Zn(II)–WT1-4 dissociation constant at pH 9.0 is 66.5 fM.
Figure 5.

Kinetics of Zn(II) removal from 22 μM Zn(II)–WT1-4 by 100 μM EDTA buffered at pH 6.65 (20 mM MES, 100 mM KCl) followed by the decrease in fluorescence emission intensity at 355 nm. The fluorescence emission intensity drops from an initial value of 145.67 au to 78.45 au in 15 min. Under similar conditions, Sénèque and Latour15 measured an equilibration time for Zn(II)–CP1-CCHH of 1600 min.
Potentiometric pH Titrations
The pH-dependent chemical speciation of WT1-4 in the apo state and with Zn(II) bound were investigated using pH titrations followed by NMR and fluorescence spectroscopies, respectively. The chemical shifts of apo-WT1-4 were assigned at pH 6.0. The pH dependence of the Cys and His amino acid chemical shifts was determined using TOCSY. The pKa values of the liganding residues were determined by measuring the proton chemical shifts of Cys5 (Cβ-H), Cys10 (Cβ-H), His23 (Cε1-H, Cδ2-H), and His27 (Cε1-H, Cδ2-H). Figure 6 shows that plots of proton chemical shift versus pH for each resonance were well-fit to single-proton pKa values, eq 12. The data are fit to pKa values of 8.0, 7.9, 6.2, and 6.9 for Cys5, Cys10, His23, and His27, respectively (Table 1). These values are close to the solution values for cysteine and histidine, as expected because of the unfolded nature of apo-WT1-4. In addition, these pKa values establish the apo-peptide speciation in Scheme 1.
Figure 6.

pH titration of the Zn(II) binding residues in WT1-4 followed by NMR spectroscopy. The pH titration curves are fit to single-proton pKa values of 8.0 (○, Cys5), 7.9 (□, Cys10), 6.2 (■, His23), and 6.9 (●, His27).
Table 1. pKa and pKaeff Values of the Zn(II) Binding Residues in apo-WT1-4 and Zn(II)–WT1-4.
| pKa | pKaeff | |
|---|---|---|
| Cys5 | 8.0 | 5.2 |
| Cys10 | 7.9 | 5.2 |
| His23 | 6.2 | 2.6 |
| His27 | 6.9 | 2.6 |
The pKaeff values of Zn(II)–WT1-4 were measured using potentiometric pH titrations followed by fluorescence spectroscopy to determine the appropriate Zn(II) proton competition equilbrium model. Figure 7 shows that titration of 0.1 M HCl into 15 μM Zn(II)–WT1-4 results in a decrease in tryptophan fluorescence. The pH titration of Zn(II)–WT1-4 is best fit to an equilibrium model involving two distinct two-proton protonation events, a coupled two-proton event at a pKa1,2eff value of 2.6 and a cooperative two-proton event at a pKa3,4eff value of 5.2, as shown in Scheme 1. These are assigned to the protonation of the Zn(II)-bound His residues (2.6) and Cys (5.2) residues (Table 1), as shown in eqs 17 and 18.
| 17 |
| 18 |
The measurement of these effective pKaeff values is critical to providing the correct Zn(II)–H+ competition model for the equilibrium presented in Scheme 1 and the speciation diagram for Zn(II)–WT1-4 shown in Figure 8. At pH values greater than 6.5, Zn(II)–WT1-4 is predominantly in the Cys2His2, cysteine thiolate/histidine imidazole, form, as shown in Figure 8. Between pH 3.0 and 5.0, the major species is Zn(II)–WT1-4 in the (CysH+)2His2, cysteine thiol/histidine imidazole, form. Lastly, below pH 2.5, the WT1-4 peptide exists mostly in the (CysH)2His2, cysteine thiol/histidine imidazolium, form without the metal bound.
Figure 7.

pH titration of 17 μM Zn(II)–WT1-4 followed by fluorescence spectroscopy. The decrease in tryptophan fluorescence emission intensity at 355 nm as the pH is lowered by addition of microliter aliquots of 0.1 N HCl is due to protonation/dissociation of the Zn(II) bound thiolate/imidazole ligands. The pH titration data is best fit to an equilibrium model involving two separate protonation events, a two-proton event with a pKa1,2eff value of 5.2 and a cooperative two-proton event with a pKa3,4eff value of 2.6.
Figure 8.

Speciation diagram of the Zn(II)–WT1-4 metal–ligand complex depicting the diimidazole–dithiolate zinc species (solid line), Zn(II)–WT1-4, the diimidiazole–dithiol zinc species (dashed line), Zn(II)–WT1-4–2H+, and the diimidazolium–dithiol species (dotted line), Zn(II)–WT1-4–4H+. The diagram was generated on the basis of the protonation behavior of the Zn(II)–WT1-4 complex in Figure 7.
Equilibrium Binding Model
The minimal complete equilibrium binding model for Zn(II)–WT1-4 established by the measured intrinsic pKa and effective pKaeff values is presented in Scheme 1. The apo-state pKa values indicated that each His and Cys ligand individually deprotonates, resulting in five species, generically LH4 to L4–, as expected. The slope of the potentiometric curves used to determine the holo-state pKaeff values show different behavior, namely, the cysteine thiols deprotonate as a pair, and the histidine imidazoliums deprotonate as a pair. The observed cooperativity in Cys and His deprotonation results in only three holo-state species, MLH42+, MLH2, and ML2–, where MLH42+ dissociates into M(II) and LH4 under the conditions of our experiment. The speciation observed in the pKa and pKaeff values is used to derive the expression for the pH dependence of the formation constant, eq 14. Therefore, measuring both the pKa and pKaeff values is critical to establishing the speciation model used to determine the formation constant.
EXAFS Spectroscopy
To verify the coordination sphere of Zn(II) in complex with WT1-4 further, the k-edge EXAFS of the metal-bound peptide was examined. As can be seen in Figure 9 and Table 2, the data are best modeled with a Cys2His2 coordination sphere. As anticipated, the Zn–S scattering is substantially stronger than the Zn–N scattering, and modeling the first shell with all low-Z scatterers leads to a more than 2-fold higher fit residual than a model that contains only sulfur (compare the first two fits in Table 2). The mixed first shell of two nitrogen/oxygen and two sulfur donors gives a nearly 10-fold lower residual than the all nitrogen fit, which is also one-third that of the all-sulfur model. To better elucidate the best model for the coordination sphere, a percent improvement (Pi) versus composition plot (inset in Figure 9), where Pi is defined as Pi = (F2S+2S – Fi)/(F4S) × 100%, was constructed.48 Comparison of the Zn(II)–WT1-4 fits (black line) with the curves previously reported for the Zn(II)–GGG–Cys2His2, Zn(II)–GGG–Cys3His, and Zn(II)–GGG–Cys4 peptides10 shows that the data for Zn(II)–WT1-4 is most consistent with that of the Zn(II)–GGG–Cys2His2 peptide. Multiple scattering fits, which have the lowest fit residual, further support Zn binding in a Cys2His2 site, with the outer shell scattering amplitude most consistent with the presence of two histidyl imidazoles in the primary coordination sphere of Zn(II). In addition, the distances of Zn–S (2.28 Å) and Zn–N (2.10 Å) in the 2S+2N (His) model are identical to the one that are previous reported for Zn(II)–GGG–Cys2His2 peptide10 and are similar to those observed in the NMR structure of full-length WT1 bound to DNA (Zn–S 2.29/2.30 Å and Zn–N 2.06/2.07 Å; PDB ID: 2JPG(17)). The corresponding distances in the X-ray structure of full-length WT1 bound to DNA (Zn–S 2.44/2.79 Å and Zn–N 2.21/2.29 Å; PDB ID: 2PRT(17)) are clearly different due to the moderate resolution of the X-ray structure, 3.15 Å. Overall, the EXAFS analysis indicates that the structure of the bound Zn(II) is the same in a single-finger (WT1-4) or the natural four-finger (WT1) construct.
Figure 9.

EXAFS Fourier transform for Zn(II)–WT1-4 (solid line) and best fit (open diamonds), modeled as a Cys2His2 coordination sphere. Inset: Percent improvement (Pi) vs composition plot for Zn(II)–WT1-4 (open symbols, black line) compared to those for characterized model peptides (gray lines, as labeled).
Table 2. EXAFS Curve Fitting Results for Zn(II)–WT1-4a.
| model | Zn–S | Zn–N | Zn–Hisb | Rfc | Ru |
|---|---|---|---|---|---|
| 4 N | 2.07 (6.2) | 238 | 420 | ||
| 4 S | 2.26 (11) | 75 | 183 | ||
| 2 S + 2 S | 2.19 (6.6), 2.32 (5.0) | 61 | 176 | ||
| 2 S + 2 N | 2.28 (2.8) | 2.10 (2.4) | 25 | 128 | |
| 2 S + 2 N (2 His) | 2.28 (2.8) | 2.10 (2.6) | 2.85 (18), 3.37 (2.2), 4.17 (10), 4.38 (16) | 23 | 66 |
Distances (Å) and disorder parameters (in parentheses, σ2 (10–3 Å2)) shown derive from fits to filtered EXAFS data. Δk = 1.5–13.2 Å–1; ΔR = 0.7–2.3 Å for first shell fits; ΔR = 0.1–4.5 Å for multiple scattering fits.
Multiple scattering paths represent combined scattering paths described in Experimental Section.
Goodness of fit (Rf for fits to filtered data; Ru for fits to unfiltered data) is defined as 1000 × (∑i = 1N{[Re(χicalc)]2 + [Im(χicalc)]2})/(∑i = 1{[Re(χiobs)]2 + [Im(χiobs)]2}), where N is the number of data points.
Discussion
The formation constant for a natural Cys2His2/ββα zinc finger has been determined using a suite of equilibrium measurements and the minimal complete equilibrium model for metal–peptide binding. The data describe the metal-ion affinity and solution speciation of the C-terminal zinc finger domain in the Wilms’ tumor suppressor protein, WT1-4, which undergoes a metal-induced protein folding event. The data demonstrate the solution speciation of Zn(II)–WT1-4 and indicate a Zn(II)–WT1-4 formation constant of 7.5 × 1012 M–1, limiting Kd value of 133 fM, which demonstrates that Zn(II) binding provides up to 17.6 kcal/mol of free energy to facilitate protein folding and assembly. These results are identical within error to those of the GGG–Cys2His2 model peptide,10 formation constant of 2.5 × 1013 M–1, limiting Kd value of 40 fM, 18.3 kcal/mol, indicating a similar, minimal cost of protein folding in the two peptides. The similarity between Zn(II)–WT1-4 and GGG–Cys2His2 extends to their pH-dependent conditional dissociation constants because of their similar Cys/His pKa and metal-bound Cys/His effective pKa values. The conditional Kd values of Zn(II)–WT1-4 are further compared with other zinc finger proteins from the literature at various pH values to reveal the cost of protein folding in the latter. These data indicate that most Cys2His2 zinc finger proteins possess a minimal cost of protein folding, <+5 kcal/mol, relative to the −17.6 kcal/mol contribution of Zn(II) binding. Furthermore, these results validate our use of the GGG–Cys2His2 model peptide to deduce the cost of protein folding in natural zinc finger proteins.
Scheme 1 shows the minimal complete equilibrium binding model for Zn(II)–WT1-4 that was established using the measured intrinsic pKa and effective pKaeff values and used to determine formation constant value, KfML (β110), of 7.5 × 1012 M–1. Figure 10 shows the conditional association constants of Zn(II)–WT1-4 as a function of solution pH fit to eq 14 derived from our minimal complete equilibrium model. The plateau at basic pH yields the Zn(II)–WT1-4 formation constant value, which is attenuated at pH values below the apo-state pKa values because of proton competition for Zn(II) binding. The slope of the attenuation approaches 4.0, as expected on the basis of the Cys2His2 coordination sphere, and plateaus below the pKaeff values to yield the Zn(II)–WT1-4-4H+ formation constant value, KfMLH4 (β114), of 1.2 M–1. The complete model allows the independently measured conditional binding constants, pKa and pKaeff values, to be validated against each other. Notably, the −18.3 kcal/mol free energy difference between the intrinsic pKa and effective pKaeff values, derived from ΔΔG = 2.303RT(∑ pKaeff – ∑ pKa), must be equivalent within error to the −17.7 kcal/mol free energy difference between the MLH42+ and ML2– formation constant values, derived from ΔΔG = −2.303RT(log KfML – log KfMLH4) by definition. This provides a stringent check of the data and a validation of the equilbrium model used.
Figure 10.

pH dependence of the conditional dissociation constant of Zn(II) complexation by WT1-4, shown as a plot of the logarithm of the association constant vs solution pH. The equilibrium binding model employed to fit the data yields a pH-independent formation constant, KfML value, of 7.5 × 1012 M–1, or a limiting dissociation constant of 133 fM, which corresponds to a reaction free energy of −17.6 kcal mol–1.
The formation constant value, KfML, of 7.5 × 1012 M–1 measured for Zn(II)–WT1-4 is identical within the 10-fold (1.4 kcal/mol) error to the 2.5 × 1013 M–1 value previously reported for the Zn(II)–GGG–Cys2His2 model peptide.10 In each case, Zn(II) binding to the Cys2His2 site contributes about −18 kcal/mol toward protein folding and metalloprotein stability. Thus, the KfML data show that the cost of protein folding in the two peptide scaffolds is identical within error, i.e. ΔGapofolding–WT1-4 = ΔGapofolding–GGG. Because we assume that the value of ΔGapofolding–GGG is close to 0 kcal/mol as a result of the lack of secondary structure in its apo-unfolded and apo-folded states, the data indicate that the cost of protein folding in WT1-4 is also close to 0 kcal/mol. One might expect the longer sequence of WT1-4 compared to that of GGG to result in a larger cost of protein folding in the former because of the greater loss of conformational entropy. However, because the value of ΔGapofolding–WT1-4 reflects the difference between the apo-unfolded and apo-folded states, it encompasses contributions from both favorable and unfavorable changes in enthalpy (ΔH) and entropy (ΔS). Restriction of the apo-unfolded ensemble to a structured apo-folded state involves an unfavorable ΔS term from the loss of conformational entropy and a favorable ΔH contribution from secondary structure formation and hydrophobic core packing in the apo-ββα structure. This situation where the unfavorable contributions to the folding free energy are offset by favorable terms appears to be the case also for the Cys3His1 site in the zinc knuckle fold of HIV-1 nucleocapsid protein (34–51)NCp7.40,41 The KfML values for (34–51)NCp7 and the corresponding GGG–Cys3His1 model peptide are 2 × 1015 and 1.5 × 1015 M–1, respectively, indicating an identical cost of protein folding between these two peptides and thus a balance between the favorable and unfavorable free energy terms between the apo-unfolded state and the apo-folded state. Lastly, a comparison of the KfML values from these Cys2His2 and Cys3His1 sites indicates that the Cys3His1site possesses higher affinity, bolstering our previous conclusion that cysteine thiolate of Cys is a better ligand than the imidazole of His for Zn(II).10
The similarity in the KfML values of Zn(II)–WT1-4 and GGG–Cys2His2 does not necessarily indicate equivalent conditional dissociation constants, Kd values, because of the influence of the ligand pKa values, both intrinsic and effective. In the case of Zn(II)–WT1-4 and GGG–Cys2His2, their Cys/His pKa values are similar, likely due to the unfolded nature of their apo states. The similarity in the KfML and the intrinsic Cys/His pKa values between the two peptides results in similar Zn(II)–Cys/Zn(II)–His effective pKaeff values, by definition. The measured Kd values for Zn(II)–WT1-4 and GGG–Cys2His2 are all within 1.5 kcal/mol of each other at an individual pH values. These data indicate that the cost of protein folding in WT1-4 is also close to 0 kcal/mol over the pH range 5–9. A close inspection of the data shows a trend in that at pH values higher than 7.4, GGG–Cys2His2 has up to a 1.0 kcal/mol tighter affinity for Zn(II), whereas Zn(II)–WT1-4 has up to a 1.5 kcal/mol tighter affinity at pH values below 7.0 because of the ligand pKa and pKaeff values. The His pKa values (6.9/6.5 in GGG vs 6.9/6.2 in WT1-4) and the pKaeff values of the Zn(II)–Cys residues (5.6 vs 5.4) are nearly identical, but the Cys pKa values (9.1/8.8 vs 8.0/7.9) and the pKaeff values of the Zn(II)–His residues (3.2/3.0 vs 2.7) show differences that lead to the observed changes in the Kd relative values. The Cys5 and Cys10 pKa values of WT1-4 are likely lower (more acidic) than those in GGG because of the presence of local cationic amino acid side chains, Arg6, Lys12, and Lys13. These pKa and pKaeff changes reflect slight differences in proton management that effect the conditional dissociation constants while not affecting the overall formation constant KfML values, which indicates the importance of determining both the pKa and pKaeff values.
Table 3 shows the values of the free energy cost of protein folding, ΔGapofolding, derived for other Cys2His2 zinc proteins using either the GGG or WT1-4 data sets. The data show that the cost of protein folding values derived are slightly different depending on whether GGG or WT1-4Kd values are used. For example, the ΔGapofolding value of ZFY,49 a Cys2His2/ββα zinc finger derived from the human Y-encoded protein, is either +2.3 or +3.7 kcal/mol at pH 6.5, based on the GGG or WT1-4 data, respectively. Similarly, the cost of folding the third Cys2His2/ββα zinc finger of the Wilms’ tumor suppressor protein, WT1-p,50 is either −0.7 or +0.7 kcal/mol at pH 6.5 based on GGG or WT1-4 data, respectively. The values of ΔGapofolding change by up to 2.1 kcal/mol (Ros8751 at pH 6.8) depending on the model data set used and the solution pH, but this does not alter our prior conclusion that the cost of protein folding in most Cys2His2 zinc fingers is minimal, ≤ 4.2 kcal/mol. Our previous conclusion that the cost of protein folding in most zinc fingers regardless of zinc coordination motif is minimal is supported by Wilcox’s recent report14 on the physiological Zn(II) affinity of three natural proteins, Sp1-3, MyT1-2, and GR-2, which contain the canonical Cys2His2, Cys3His1, and Cys4 coordination motifs, respectively.
Table 3. Cost of Protein Folding in Zn(II)–Cys2His2 Proteins.
| protein | pH | protein Kd (log Ka) | protein ΔGML-obs (kcal/mol) | GGG ΔGML-obs (kcal/mol) | ΔGapofoldingGGG (kcal/mol) | WT1-4 ΔGML–obs (kcal/mol) | ΔGapofoldingWT-1 (kcal/mol) | protein fold |
|---|---|---|---|---|---|---|---|---|
| SUP3766 | 6.4 | 21 nM (7.7) | –10.5 | –10.9 | +0.4 | –12.3 | +1.8 | ββα |
| WT1-p50 | 6.5 | 1.9 nM (8.7) | –11.9 | –11.2 | –0.7 | –12.6 | +0.7 | ββα |
| ZFY49 | 6.5 | 300 nM (6.5) | –8.9 | –11.2 | +2.3 | –12.6 | +3.7 | ββα |
| Ros8751 | 6.8 | 36 nM (7.4) | –10.2 | –11.8 | +1.6 | –13.9 | +3.7 | βββαα |
| CP159 | 7.0 | 8.5 pM (11.1) | –15.1 | –14.0 | –1.1 | –14.5 | –0.6 | ββα |
| CP115 | 7.0 | 1.2 fM (14.9) | –20.3 | –14.0 | –6.3 | –14.5 | –5.8 | ββα |
| SP1-367 | 7.0 | 250 pM (9.6) | –13.1 | –14.0 | +1.0 | –14.5 | +1.4 | ββα |
| SP1-314 | 7.4 | 20 nM (7.7) | –10.5 | –16.3 | +5.8 | –15.6 | +5.1 | ββα |
| HCCHp56 | 7.4 | 54 nM (7.3) | –9.9 | –16.3 | +6.4 | –15.6 | +5.7 | NDa |
| Ant-F57 | 7.5 | 12 nM (7.9) | –10.8 | –16.5 | +5.7 | –15.7 | +4.9 | ββα |
Not determined.
Several other lines of evidence in the literature support these conclusions. First, the successful redesign of a ββα zinc finger to be folded in the apo state did not increase the Zn(II) affinity dramatically, as would be expected if the cost of protein folding was large.52,53 Second, molecular dynamics simulations of a zinc finger domain suggest that the ensemble of apo-unfolded states corresponds to the native state in an average sense.54 Third, the successful computational redesign of a zinc finger into a stable ββα fold without the Zn(II) or the ligands indicates that hydrophobic interactions are sufficient to compensate for the loss of Zn(II) binding.55 Lastly, the MI3 and MI4 proteins, natural homologues of the prokaryotic Cys2His2/βββαα zinc finger protein, Ros87, lack Zn(II) yet achieve the same functional fold.56
There are at least two examples of zinc finger proteins with higher costs of protein folding in the literature, i.e. 6 kcal/mol. A +5.7 kcal/mol cost of protein folding, based on GGG and shown in Table 3, is observed for the artificial “antennafinger” Ant-F57 at pH 7.5. The observed higher cost of protein folding is ascribed to the fact that Zn(II) binding induces a conformational change between two distinct folded states, a helical apo state and a less helical holo state. Thus, the free energy associated with Zn(II) binding is used to reorganize the helical apo-state conformation that is incommensurate with Zn(II) binding into the holo state. A similar situation may be observed for HCCHp,58 a Cys2His2 zinc fingerlike peptide from the HIV-1 virion infectivity factor, which is reported to have some secondary structure in the apo state. The reported Kd value of 54 nM at pH 7.4 for HCCHp suggests a + 6.4 kcal/mol cost of protein folding compared to GGG–Cys2His2.
In our original report, we noted that Berg’s designed Cys2His2/ββα zinc finger Consensus Peptide 159,60 (CP1-CCHH) possessed a Zn(II) affinity that was slightly tighter than GGG and therefore had a ΔGapofolding value of −1.3 kcal/mol, which is within the error of the Kd measurements. However, a recent report by Sénèque and Latour15 reevaluates both the zinc affinity of CP1-CCHH and the metal-ion exchange kinetics, and their findings differ significantly from Berg’s initial reports. Sénèque and Latour report a KfML (or β110) value of 3.2 × 1017 M–1 for Zn(II)–CP1-CCHH and a Kd value at pH 7.0 of 1.25 fM. These values are significantly tighter than our measured Zn(II)–GGG–Cys2His2 and Zn(II)–WT1-4 values, KfML values of 2.5 × 1013 and 7.5 × 1012 M–1, and our measured Kd values at pH 7.0 of 56 and 22 pM, respectively, as well as Berg’s reported Zn(II)–CP1-CCHHKd value at pH 7.0 of 5.7 pM.55 Sénèque and Latour15 also observe slow metal-exchange kinetics (teq > 24 h at pH 6.65) when using EDTA competition titrations to determine the Kd values of Zn(II)–CP1-CCHH, whereas we observe rapid kinetics for metal-ion exchange between Zn(II)–WT1-4 and EDTA (teq < 15 min, at pH 6.65, shown in Figure 5) and Zn(II)–GGG–Cys2His2 (teq < 3 min, at pH 6.5), and Berg61 measured rapid displacement of Co(II) by Zn(II) in CP1-CCHH. Lastly, Sénèque and Latour observe that Cys to His substitutions yield Zn(II) affinities in the order CP1-CCCH > CP1-CCCC > CP1-CCHH, whereas we observe the trend GGG–Cys4 > GGG–Cys3His > GGG–Cys2His2. This difference likely reflects the geometic restrictions imposed by the folded holo state of the CP1 peptides.
The discrepancy between the CP1-CCHH data of Sénèque and Latour and Berg may reflect the different experimental conditions used to determine the Zn(II)–CP1-CCHHKd values. Sénèque and Latour studied Zn(II) binding using UV (Zn–S charge transfer band, ε220 = 8700 M–1cm–1) and CD (protein folding) spectroscopy in phosphate buffer in the presence of the water-soluble reductant TCEP, tris(2-carboxyethyl)phosphine, whereas Berg determined the Zn(II)–CP1-CCHHKd values with UV–vis spectroscopy by Co(II) competition under strictly anaerobic conditions without TCEP or phosphate buffer. In our own experiments, we avoided the use of phosphate buffer and TCEP because of their interactions with Zn(II), Zn3[PO4]2KSP value of 9.0 × 10–33 and TCEP KfML of Zn(II)–TCEP is 813,62 and we utilized strictly anaerobic conditions. Our measured Kd values for Zn(II)–WT1-4 and Zn(II)–GGG9 agree with the thermodynamic studies of CP1-CCHH and its variants by Berg and others.9,60,63,64 Additionally, we observe that the equilibria of metal-ion binding and metal-ion removal by EDTA are established within 15 min for both WT1-4 and GGG, consistent with the metal-ion substitution kinetics of CP1-CCHH. Furthermore, our results agree with other thermodynamic studies on natural zinc fingers in the literature, as shown in Table 3.
While resolution of the differences observed in Zn(II)–CP1-CCHHKd values is beyond the scope of this report, the reported KfML (or β110) value may have a significant impact on our method for deducing the cost of protein folding in zinc finger peptides and proteins. Sénèque and Latour15 report that CP1-CCHH binds Zn(II) 13 900-fold, or 5.6 kcal/mol, tighter than GGG–Cys2His2 and 42 700-fold, or 6.3 kcal/mol, tighter than WT1-4 based on the KfML values. This finding suggests that it costs +5.6 kcal/mol to fold GGG–Cys2His2 and therefore +5.6 kcal/mol should be added to all of the ΔGapofolding values derived from GGG–Cys2His2. In order to validate our approach to determining the cost of protein folding in Zn(II) proteins further, we are currently determining the KfML value of human transcription factor IIB (TFIIB), a Cys3His1 zinc ribbon protein that is folded into the same structure in both the apo and holo states, as shown by NMR spectroscopy.65 Thus, we expect the measured KfML value to reflect the zero cost of protein folding in TFIIB.
Conclusions
In the present work, we have evaluated the formation constant for a natural zinc finger protein domain, the C-terminal zinc finger of Wilms’ tumor suppressor, in an effort to validate our protein folding free energy calculation methodology with a natural zinc finger domain over a wide pH range. A comparison of the formation constants and the condition dissociation constants at physiological pH demonstrate that the Cys2His2 zinc-binding sites in WT1-4 and GGG have comparable affinities, within the 1.4 kcal/mol (10-fold) error of the measurements. This fact leads to the conclusion that the free energy cost of protein folding in the natural zinc finger, WT1-4, and the simple peptide, GGG, is comparable. Furthermore, given the lack of secondary structure in GGG, we assert that the free energy cost of protein folding in GGG is close to 0 kcal/mol. It then follows that the cost of protein folding in WT1-4 is also close to 0 kcal/mol.
Acknowledgments
This work was supported by NIH grant SC3-GM089634. The National Synchrotron Light Source is supported by the U.S. Department of Energy.
The authors declare no competing financial interest.
Funding Statement
National Institutes of Health, United States
References
- Klug A. Q. Rev. Biophys. 2010, 43, 1–21. [DOI] [PubMed] [Google Scholar]
- Burdach J.; O’Connell M. R.; Mackay J. P.; Crossley M. Trends Biochem. Sci. 2012, 37, 199–205. [DOI] [PubMed] [Google Scholar]
- Paul I.; Cui J.; Maynard E. L. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 18475–18480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stenmark H.; Aasland R. J. Cell Sci. 1999, 112, 4175–4183. [DOI] [PubMed] [Google Scholar]
- Kröncke K. D.; Klotz L. O. Antiox. Redox Signaling 2009, 11, 1015–1027. [DOI] [PubMed] [Google Scholar]
- Frankel A. D.; Berg J. M.; Pabo C. O. Proc. Natl. Acad. Sci. U.S.A. 1987, 84, 4841–4845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berg J. M. Curr. Opin. Struct. Biol. 1993, 3, 11–16. [Google Scholar]
- Magyar J. S.; Godwin H. A. Anal. Biochem. 2003, 320, 39–54. [DOI] [PubMed] [Google Scholar]
- Blasie C. A.; Berg J. M. Biochemistry 2002, 41, 15068–15073. [DOI] [PubMed] [Google Scholar]
- Reddi A. R.; Guzman T.; Breece R. M.; Tierney D. L.; Gibney B. R. J. Am. Chem. Soc. 2007, 129, 12815–12827. [DOI] [PubMed] [Google Scholar]
- Reddi A. R.; Gibney B. R. Biochemistry 2007, 46, 3745–3758. [DOI] [PubMed] [Google Scholar]
- Vallee B. L.; Auld D. S. Acc. Chem. Res. 1993, 26, 543–551. [Google Scholar]
- Gockel P.; Vahrenkamp H.; Züberbuhler A. D. Helv. Chim. Acta 1993, 76, 511–520. [Google Scholar]
- Rich A. M.; Bombarda E.; Schenk A. D.; Lee P. D.; Cox E. H.; Spuches A. M.; Hudson L. D.; Kieffer B.; Wilcox D. E. J. Am. Chem. Soc. 2012, 134, 10405–10418. [DOI] [PubMed] [Google Scholar]
- Sénèque O.; Latour J.-M. J. Am. Chem. Soc. 2010, 132, 17760–17774. [DOI] [PubMed] [Google Scholar]
- Yang L.; Han Y.; Suarez Saiz F.; Minden M. D. Leukemia 2007, 21, 868–876. [DOI] [PubMed] [Google Scholar]
- Stoll R.; Lee B. M.; Debler E. W.; Laity J. H.; Wilson I. A.; Dyson H. J.; Wright P. E. J. Mol. Bio. 2007, 372, 1227–1245. [DOI] [PubMed] [Google Scholar]
- Solid-Phase Synthesis: A Practical Guide; Kates S. A., Albericio F., Eds.; Marcel Dekker: New York, 2000. [Google Scholar]
- Hwang T.-L.; Shaka A. J. J. Magn. Reson. 1998, 135, 280–287. [DOI] [PubMed] [Google Scholar]
- Delaglio F.; Grzesiek S.; Vuister G. W.; Zhu G.; Pfeifer J.; Bax A. J. Biomol. NMR 1995, 6, 277–293. [DOI] [PubMed] [Google Scholar]
- Johnson B. A.; Blevins R. A. J. Biomol. NMR 1994, 4, 603–614. [DOI] [PubMed] [Google Scholar]
- Wüthrich K.NMR of Proteins and Nucleic Acids; John Wiley and Sons: New York, 1986; p 320. [Google Scholar]
- Thomas P. W.; Stone E. M.; Costello A. L.; Tierney D. L.; Fast W. Biochemistry 2005, 44, 7559–7569. [DOI] [PubMed] [Google Scholar]
- Ankudinov A. L.; Ravel B.; Rehr J. J.; Conradson S. D. Phys. Rev. B 1998, 58, 7565–7576. [Google Scholar]
- Costello A. L.; Periyannan G.; Yang K. W.; Crowder M. W.; Tierney D. L. J. Biol. Inorg. Chem. 2006, 11, 351–358. [DOI] [PubMed] [Google Scholar]
- Smith R. M.; Martell A. E.; Motekaitis R. J.. NIST Critically Selected Stability Constants of Metal Complexes; National Institutes of Standards and Technology: Gaithersburg, MD, 2001; Vol. 46. [Google Scholar]
- Martell A. E.; Smith R. M.. Critical Stability Constants; Plenum Press: New York, 1974; Vol. 1. [Google Scholar]
- Cavanagh J.; Fairbrother W. J.; Palmer A. G. III; Skelton N. J.. Protein NMR Spectroscopy: Principles and Practice; Academic Press: San Diego, CA, 1996. [Google Scholar]
- Razin S. V.; Borunova V. V.; Maksimenko O. G.; Kantiddze O. L. Biochemistry (Moscow) 2012, 77, 217–226. [DOI] [PubMed] [Google Scholar]
- Carroll D. Curr. Gene Ther. 2011, 11, 2–10. [DOI] [PubMed] [Google Scholar]
- Auld D. S. BioMetals 2001, 14, 271–313. [DOI] [PubMed] [Google Scholar]
- Maret W.; Li Y. Chem. Rev. 2009, 109, 4682–4707. [DOI] [PubMed] [Google Scholar]
- Maret W. Adv. Nutr. 2013, 4, 82–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andreini C.; Bertini I.; Cavallaro G. PLoS One 2011, 6, e26325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berg J. M. Curr. Opin. Struct. Biol. 1993, 3, 11–16. [Google Scholar]
- Laity J. H.; Lee B. M.; Wright P. E. Curr. Opin. Struct. Biol. 2001, 11, 39–46. [DOI] [PubMed] [Google Scholar]
- Andreini C.; Banci L.; Bertini I.; Rosato A. J. Proteome Res. 2006, 5, 196–201. [DOI] [PubMed] [Google Scholar]
- Andreini C.; Bertini I. J. Inorg. Biochem. 2012, 111, 150–6. [DOI] [PubMed] [Google Scholar]
- Gibney B. R.Metallopeptides as Tools To Understand Metalloprotein Folding and Stability. In Protein Folding and Metal Ions: Mechanisms, Biology and Disease; Gomes C., Wittung-Stafshede P., Eds.; CRC Press: Boca Raton, FL, 2011; pp 227–245. [Google Scholar]
- Mély Y.; De Rocquigny H.; Morellet N.; Roques B. P.; Gérard D. Biochemistry 1996, 35, 5175–5182. [DOI] [PubMed] [Google Scholar]
- Mély Y.; Cornille F.; Fournie-Zaluski M.; Darlix J.; Roques B. P.; Gérard D. Biopolymers 1991, 31, 899–906. [DOI] [PubMed] [Google Scholar]
- Pelletier J.; Bruening W.; Kastan C. E.; Mauer S. M.; Manivel J. C.; Striegel J. E. Cell 1991, 67, 437–447. [DOI] [PubMed] [Google Scholar]
- Drummond I. A.; Rupprecht H. D.; Rohwer-Nutter P.; Lopez-Guisa J. M.; Madden S. L.; Rauscher F. J. III; Sukhatme V. P. Mol. Cell. Biol. 1994, 14, 3800–3809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coppes M. J.; Campbell C. E.; Williams B. R. G. FASEB J. 1993, 7, 886–894. [DOI] [PubMed] [Google Scholar]
- Haber D. A.; Timers T. H.; Pelletier J.; Sharp P. A.; Housman D. E. Proc. Natl. Acad. Sci. U.S.A. 1992, 89, 6010–6014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lakowicz J. R.Principles of Fluorescence Spectroscopy; Plenum Press: New York, 1983. [Google Scholar]
- Harris H. L.; Hudson B. S. Biochemistry 1990, 29, 5276–5285. [DOI] [PubMed] [Google Scholar]
- Clark-Baldwin K.; Tierney D. L.; Govindaswamy N.; Gruff E. S.; Kim C.; Berg J.; Koch S. A.; Penner-Hahn J. E. J. Am. Chem. Soc. 1998, 120, 8401–8409. [Google Scholar]
- Lachenmann M. J.; Ladbury J. E.; Phillips N. B.; Narayana N.; Qian X.; Weiss M. A. J. Mol. Biol. 2002, 316, 969–989. [DOI] [PubMed] [Google Scholar]
- Lachenmann M. J.; Ladbury J. E.; Dong J.; Huang K.; Carey P.; Weiss M. A. Biochemistry 2004, 43, 13910–13925. [DOI] [PubMed] [Google Scholar]
- Palmieri M.; Malgieri G.; Russo L.; Baglivo I.; Esposito S.; Netti F.; Del Gatto A.; de Paola I.; Zaccaro L.; Pedone P. V.; Isernia C.; Milardi D.; Fattorusso R. J. Am. Chem. Soc. 2013, 135, 5220–5228. [DOI] [PubMed] [Google Scholar]
- Struthers M. D.; Cheng R. P.; Imperiali B. J. Am. Chem. Soc. 1996, 118, 3073–3081. [Google Scholar]
- Walkup G. K.; Imperiali B. J. Am. Chem. Soc. 1997, 119, 3443–3450. [Google Scholar]
- Zagrovic B.; Snow C. D.; Khaliq S.; Shirts M. R.; Pande V. S. J. Mol. Biol. 2002, 323, 153–164. [DOI] [PubMed] [Google Scholar]
- Dahiyat B. I.; Mayo S. L. Science 1997, 278, 82–87. [DOI] [PubMed] [Google Scholar]
- Baglivo I.; Palmieri M.; Rivellino A.; Netti F.; Russo L.; Esposito S.; Iacovino R.; Farina B.; Isernia C.; Fattorusso R.; Pedone P. V.; Malgieri G. Biochim. Biophys. Acta 2014, 1844, 497–504. [DOI] [PubMed] [Google Scholar]
- Hori Y.; Sugiura Y. J. Am. Chem. Soc. 2002, 124, 9362–9363. [DOI] [PubMed] [Google Scholar]
- Giri K.; Scott R. A.; Maynard E. L. Biochemistry 2009, 48, 7969–7978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krizek B. A.; Amann B. T.; Kilfoil V. J.; Merkle D. L.; Berg J. M. J. Am. Chem. Soc. 1991, 113, 4518–4523. [Google Scholar]
- Krizek B. A.; Merkle D. L.; Berg J. M. Inorg. Chem. 1993, 32, 937–940. [Google Scholar]
- Buchsbaum J. C.; Berg J. M. Inorg. Chim. Acta 2000, 297, 217–219. [Google Scholar]
- Krezel A.; Latajka R.; Bujacz G. D.; Bal W. Inorg. Chem. 2003, 42, 1994–2003. [DOI] [PubMed] [Google Scholar]
- Shi Y.; Beger R. D.; Berg J. M. Biophys. J. 1993, 64, 749–753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berezovskaya Y.; Armstrong C. T.; Boyle A. L.; Porrini M.; Woolfson D. N.; Barran P. E. Chem. Commun. 2010, 47, 412–414. [DOI] [PubMed] [Google Scholar]
- Ghosh M.; Elsby L. M.; Mal T. K.; Gooding J. M.; Roberts S. G. E.; Ikura M. Biochem. J. 2004, 378, 317–324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isernia C.; Bucci E.; Leone M.; Zaccaro L.; Di Lello P.; Digilio G.; Esposito S.; Saviano M.; De Blasio B.; Pedone C.; Pedone P. V.; Fattorusso R. ChemBioChem. 2003, 4, 171–180. [DOI] [PubMed] [Google Scholar]
- Posewitz M. C.; Wilcox D. E. Chem. Res. Toxicol. 1995, 8, 1020–1028. [DOI] [PubMed] [Google Scholar]
- The PyMOL Molecular Graphics System, version 1.5.0.4; Schrödinger, LLC.