
Figure 2.
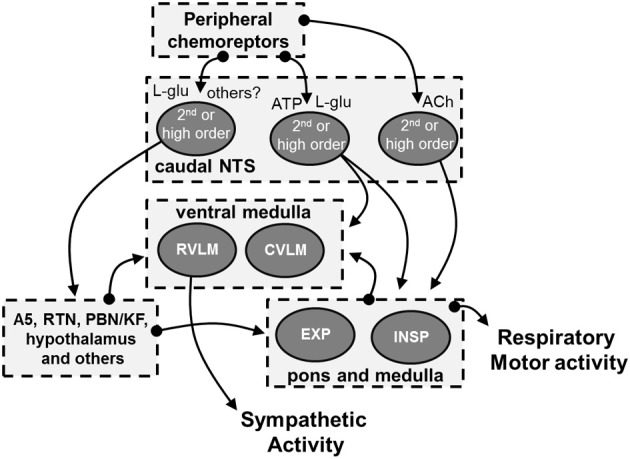
Schematic drawing showing the possible cellular and neurochemical mechanisms of the cNTS mediating the processing of peripheral chemoreceptors inputs. The stimulation of peripheral chemoreceptors evokes the release of L-glutamate in the cNTS (Mifflin, 1992; Andresen and Kunze, 1994). Experimental evidence indicates the L-glutamate, in association with ATP, is essential for the processing of sympatho-excitatory response of the peripheral chemoreflex in the cNTS (Machado and Bonagamba, 2005; Braga et al., 2007). It is suggested that both glutamatergic and purinergic systems interact and activate cNTS neurons that send projections to pre-sympathetic neurons of the RVLM (Accorsi-Mendonca et al., 2009, 2013). L-glutamate is also proposed to mediate the activation of other neural pathways that are important for the sympatho-excitatory component of peripheral chemoreflex, including those to the A5 region, retrotrapezoid nucleus (RTN), parabrachial nucleus/Kölliker-Fuse complex (PBN/KF) and hypothalamus (Koshiya and Guyenet, 1994; Olivan et al., 2001; Haibara et al., 2002; Reddy et al., 2005; Takakura et al., 2006; Queiroz et al., 2011; Song et al., 2011; Taxini et al., 2011; King et al., 2012). In addition to L-glutamate and ATP, our recent studies suggest that ACh in the cNTS significantly contributes to the tachypnea and the patterning of sympathetic response of peripheral chemoreflex (Furuya et al., 2014). We hypothesize that ACh activates cNTS neurons that send projections to respiratory neurons of the ventral medulla that, in turn, promotes the stimulation of inspiratory motor activity and the patterning of sympathetic activity. The latter may involve the modulation of the neurons of RVLM and of the caudal ventrolateral medulla (CVLM) (Mandel and Schreihofer, 2009; Moraes et al., 2012c). However, it remains to be elucidated the sources of ACh in the cNTS as well as the efferent pathways activated by ACh in response to peripheral chemoreflex activation.