

Abstract
It has been well established that the circadian clock plays a crucial role in the regulation of almost every physiological process. It also plays a critical role in pathophysiological states including those of obesity and diabetes. Recent evidence has highlighted the potential for targeting the circadian clock as a potential drug target. New studies have also demonstrated the existence of “clock-independent effects” of the circadian proteins, leading to exciting new avenues of research in the circadian clock field in physiology. The goal of this review is to provide an introduction to and overview of the circadian clock in physiology, including mechanisms, targets, and role in disease states. The role of the circadian clocks in the regulation of the cardiovascular system, renal function, metabolism, the endocrine system, immune, and reproductive systems will be discussed.
Keywords: circadian, clock, metabolism, cardiovascular, immune, endocrine, renal, diabetes, obesity
the concept of circadian rhythms was first documented in the 18th century, when the French astronomer de Mairan recorded his observation that the opening and closing of heliotrope plant leaves occurred independently of sunlight (65). Now it is apparent that almost all facets of physiology display rhythmic oscillations from the simplest archaebacteria to humans (79); these rhythms are controlled by an internal circadian clock. The purpose of this review is to provide an overview of the role the circadian clock plays in the regulation of a variety of mammalian physiological functions.
The circadian clock can be divided into two major components: the central clock, residing in the suprachiasmatic nucleus (SCN) of the brain, and the peripheral clocks that are present in nearly every tissue and organ system (3, 4). Light signals and other external factors such as feeding cues entrain the central circadian clock, which in turn synchronize the peripheral clocks (117). Light enters through the retina of the eye, causing electrical signals to pass through the retinal hypothalamic tract, which are converted to chemical signals in the SCN. The peripheral clocks can be entrained independently of the central clock by food cues; however, the nature of the relationship between the central and peripheral clocks is still being investigated (117). A prevailing view of the association between the central and peripheral clocks is that of conductor and orchestra (29). Each peripheral clock can play music independently, but direction by the central clock helps the peripheral clocks play in harmony.
On a molecular level, the circadian clock mechanism can be described by the Transcription Translation Oscillating (TTO) Loop (Fig. 1) (45). The circadian proteins CLOCK and BMAL1 heterodimerize and interact with E-box response elements in the promoters of target genes, which drive the positive transcription arm of the TTO loop. Two of these target genes encode the circadian proteins Period (Per) (homologs: 1, 2, and 3) and Cryptochrome (Cry) (homologs: 1 and 2). These two proteins interact, translocate into the nucleus, and inhibit the activity of CLOCK and BMAL1, which promotes the transcriptional repression arm of the TTO loop. Other accessory proteins include D-site albumin binding protein (DBP), and the nuclear orphan receptors retinoid-related orphan receptor (ROR) and REV-ERB, which modulate the activity of the loop through activation (ROR) or repression (REV-ERBα) of Bmal1 gene expression (reviewed in Ref. 155). Further work has shown the importance of additional transcriptional loops including the role of the Per proteins in repressing their own transcription and that of Cry1/2 (100). Interestingly, a recent compelling study used global chromatin immunoprecipitation (ChIP)-Seq to identify DNA binding interactions of the core clock proteins. This study found that Per (1–2) and Cry (1–2) possessed “CLOCK-independent” interactions, which are interactions that do not involve the CLOCK-BMAL1 complex. These clock-independent interactions are still time dependent but do not appear to involve the full participation of the other core circadian clock proteins. The evidence for these interactions and their possible roles will be discussed in the review. There is also increasing evidence for the role of posttranslational modifications in the regulation of the circadian clock. Regulation of nuclear entry and export of the Per proteins is integral for overall circadian control (149). Casein kinase-1 δ/ε (CK1δ/ε) are involved in the phosphorylation of the Per proteins. CK1δ/ε-mediated phosphorylation results in Per1 nuclear entry (70, 116). Protein phosphatase 1 (PP1) interacts with Per1 and likely dephosphorylates the protein, resulting in cytoplasmic localization of Per1 (70). It is likely that the balance between CK1δ/ε phosphorylation and dephosphorylation by PP1 may be a key regulator of the posttranslational regulation of the Per proteins.
Fig. 1.
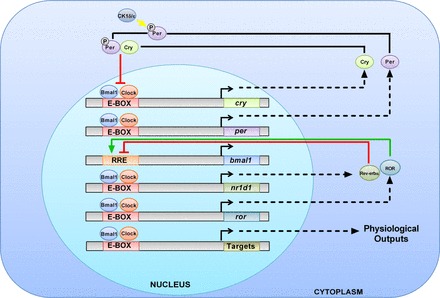
Transcription-Translation Oscillating (TTO) Loop model. In the positive arm of the TTO loop, Clock, and Bmal1 heterodimerize to activate transcription of circadian target genes, including Per (homologs: 1–3), Cry (homologs:1–2), ROR, and Nr1d1(REV-ERB-α). In the negative arm of the TTO loop, Per and Cry are thought to interact and inhibit the action of Bmal1 and Clock, thereby decreasing their own transcription. ROR and REV-ERBα mediate opposing actions on Bmal1 gene expression. Per1 is phosphorylated by the circadian regulatory kinases casein kinase 1δ/ε, which allows it to enter the nucleus.
It has been well established that the circadian clock plays an integral role in the regulation of nearly all physiological processes. This review will cover the most recent experimental evidence supporting a key role for the circadian clock in the regulation of the cardiovascular system and renal function, metabolism, and the endocrine, immune, and reproductive systems. The use of global circadian knockout (KO) mouse models together with human studies has clearly demonstrated the importance of the circadian clock to normal physiology. The increasing use of cell-type and tissue-specific KO models should further aid our understanding of the mechanism underlying the regulation of physiological function by the circadian clock.
Cardiovascular System and the Kidney
The role of the circadian clock in the cardiovascular system is a very active area of investigation with a focus on the regulation of blood pressure. This section will discuss blood pressure phenotypes in circadian clock gene KO models and introduce the concept or circadian blood pressure patterns. The circadian clock in the heart, the vasculature, and the kidney will be considered as well.
Circadian clock proteins and blood pressure.
Multiple circadian KO animals have a blood pressure phenotype. CLOCK KO mice are hypotensive compared with wild-type (WT) mice and display a mild diabetes insipidus (157). This phenotype may be due to decreased levels of 20-HETE in CLOCK KO animals; 20-HETE is a vasoconstrictor of the preglomerular arteries and is involved in raising blood pressure (95). We have recently shown that Per1 KO mice exhibit dramatically lower blood pressure than WT controls (135). One possible explanation for this phenotype is the increased levels of ET-1 in the renal medulla of these animals. Endothelin-1 (ET-1) is a robust inhibitor of epithelial sodium channel (ENaC) activity via an endothelin-B receptor-nitric oxide-dependent pathway (11). However, the effect of ET-1 on blood pressure in these animals remains to be determined. Cry1/2 double KO mice exhibit salt-sensitive hypertension due to increased synthesis of aldosterone (30). This was caused by increased production of 3-β-dehydrogenase-isomerase, an enzyme in the aldosterone synthesis pathway. BMAL1 KO mice have reduced blood pressure during the active phase (23) and thus lack the circadian variation in blood pressure.
It has been established since the 20th century that blood pressure decreases at night by around 10% and rises during the day (2). The role of circadian clock proteins in the “dipping” of blood pressure is not understood. People who do not experience night time dipping have been shown to have increased cardiovascular events (69). These individuals are termed nondippers. The exact mechanism or reasons for nondipping is unknown. However, it is known that Type 2 diabetics (6), hypertensives (salt sensitive or non-salt sensitive) (145) (reviewed in Ref. 119), and people with chronic kidney disease (89) are known to have increased occurrence of nondipping. One contributing mechanism may be renal sodium handling (reviewed in Ref. 134). Evidence suggests that dysregulated sodium reabsorption contributes to nondipping (13, 34, 130, 131) and that treatment with angiotensin receptor blockers or diuretics can convert a nondipper to a dipper (42, 90, 107, 144). Others have proposed that circadian blood pressure rhythms are independent of renal sodium handling [reviewed by Sachdeva and Weder (119)]. To briefly summarize, in normal individuals who at night sleep lying down in a supine position, sodium excretion and blood pressure both dip at night. However, in individuals that are active or upright at night, blood pressure dips, but sodium excretion does not. Supine positioning at night does not appear to be required for rhythmic blood pressure in normotensive males and females (123). It is likely that a combination of factors including sympathetic activity (85, 138), hormone signaling (37), nitric oxide (66), and sodium reabsorption all contribute in some way to circadian blood pressure control. Much work is needed to determine which factors or mechanisms are involved in the circadian control of blood pressure.
The circadian clock and the heart: metabolism, function, and disease.
In recent years, the role of the circadian clock in the heart has been investigated by Young and colleagues. For more detailed information on the role of the circadian clock in the heart, readers are directed to one of his excellent reviews (33). Below, we focus on three major areas that have come to the forefront of circadian research in the heart: metabolism, cardiac function, and disease.
One of the first major advances in the heart circadian clock field was the generation of a cardiomyocyte cell-specific CLOCK mutant (CCM) mouse (32). With the use of this animal model, multiple genes that played important roles in metabolism were identified, such as adiponutrin, peroxisome proliferator-activated receptor-α (PPARα), PPARγ coactivator-1α (PGC1α), among others (reviewed in Ref. 33). This is not surprising, as these genes have also been shown to be circadian targets in the liver as well (133) (reviewed in Ref. 9).
Heart rate decreases at night and rises during the day (reviewed in Ref. 33). CCM mice have reduced heart rate, specifically during the active phase, meaning they do not experience the expected surge in heart rate during the active phase. Interestingly, CCM mice also exhibit an “athlete's heart” phenotype, which is characterized by decreased heart rate, increased heart muscle mass, and a larger ejection fraction. This phenotype appears to be due to elevated AKT signaling and inactivation of glycogen synthase kinase (reviewed in Ref. 33). The exact mechanism for the circadian control of heart rate, however, is not fully understood. PPARγ (152) has been implicated, via an unknown mechanism involving BMAL1 in the endothelium. It is also known that the expression of multiple intracellular calcium channels, critical for heart rate regulation, are controlled by the circadian clock (61). Myocardial repolarization appears to be controlled by the circadian clock as well, through a mechanism involving regulation of Krüppel-like factor 15 (Klf15) (55). Klf15 is a transcription factor that regulates expression of potassium channels that are responsible for the generation of polarization. Further evidence for the importance of the clock in the heart comes from a recent study showing that BMAL1 KO mice develop dilated cardiomyopathy with age, in which the heart becomes weakened and enlarged, resulting in decreased cardiac output (73).
It is well known that adverse cardiovascular events, including heart attack and stroke, peak in the early morning. This has been attributed to the steep rise in blood pressure and heart rate in the morning, which results in increased oxygen demand for the heart (reviewed in Ref. 139). Expression of the plasminogen activator inhibitor 1 (PAI-1) is a circadian target in rodents and humans (92, 121) and is downregulated in the CLOCK mutant mouse (Δ19) (153). This mutation prevents circadian transcriptional activity of the CLOCK protein, but because CLOCK can still interact with BMAL1 and DNA, the mutation has a dominant negative effect. PAI-1 is involved in thrombus formation. This is important as PAI-1 downregulates tissue-plasminogen activator, which is involved in the opening of occluded vessels (reviewed in Ref. 139). With the use of a thrombotic vascular occlusion model, in which the femoral artery is occluded, it was demonstrated that the time to vascular occlusion exhibited a circadian variation (153). Other studies, which used an ischemia-reperfusion (I/R) model on WT and CCM mice, showed that WT mice who had I/R during the inactive period had larger infarct sizes and greater fibrosis than mice who had I/R at the start of the active period (31). These data suggest that the magnitude of ischemic injury also exhibits a circadian pattern. This effect was lost in CCM mice, demonstrating a role for the circadian clock in the regulation of this process. This effect was mirrored in two human studies (39, 115), which showed that infarct size and myocardial injury displayed circadian oscillations, with highest injury occurring during the early morning hours (5:00–6:00 AM). Hypoxia-inducible factor-1α (HIF-1α) is involved in enhancing the ischemic adaptation of the heart, through the enrichment of the glycolytic capacity of the heart. There may be a role for Per2 in this process as it has been recently shown that HIF-1α is stabilized through adenosine receptor A2b-mediated stability of Per2 (36).
Several studies in rodents and humans have demonstrated the importance of the circadian clock in cardiovascular disease, through observation of cardiovascular phenotypes associated with circadian disruption or desynchronization. One study used a mouse transverse aortic constriction model, which causes pressure overload-induced cardiac hypertrophy. In this model, a 10-h/10-h light-dark cycle was used to disrupt the circadian clock in mice, which led to increased cardiac hypertrophy and reduced contractility compared with mice on a normal 12-h/12-h light-dark cycle (86). Furthermore, in this same model, it was shown that nonblood pressure-dependent antihypertrophic benefits of the angiotensin-converting enzyme (ACE) inhibitor captopril were increased more during the inactive phase than the active phase (87). Interestingly, using the same model, it was demonstrated that multiple genes involved in cardiac remodeling, glycolysis, and fatty acid metabolism were significantly altered. These genes exhibited differences in light and dark expression, with significant elevations of multiple genes, including kinesin family member 5B and matrix metallopeptidase 2, during the light phase (142). This study highlights the importance of cardiac remodeling that occurs during sleep (reviewed in Ref. 33). In humans, desynchronization leads to an increased risk for cardiovascular events. Desynchronization can be caused by rotating shift work or chronic jet lag. Multiple studies have demonstrated that shift workers who have rotating schedules have increased risk for cardiovascular events. Japanese males with rotating shift work had a 2.3-fold higher risk of death due to ischemia of the heart (41). Another study showed that rotational shift-working African-American women displayed an increased hazard ratio for cardiovascular events (75). These observations highlight the importance of maintaining healthy circadian function for cardiovascular health.
Circadian clock and the vasculature: endothelial dysfunction, nitric oxide, and role of BMAL1.
It was first shown during the mid-1900s that blood flow in humans decreases during the night and increases during the day (58). Vascular functions such as sympathetic and vascular tones, adrenergic receptor agonist response, forearm vascular resistance, and flow-mediated dilation exhibit circadian variations as well (53, 101).
Multiple studies done by Rudic and colleagues have demonstrated the importance of the clock, specifically the circadian protein BMAL1, in the vascular endothelium. It was demonstrated that BMAL1 KO mice exhibit endothelial dysfunction due to nitric oxide uncoupling, which lead to increased superoxide formation (5). This was at least partially due to dysregulation of biopterins, which are involved in nitric oxide formation. Rudic and colleagues (17) also demonstrated that BMAL1 KO or Per2/3 double KO mice developed arteriosclerotic disease. Another study has shown similar effects in Per2 KO mice in that aortic rings from Per2 KO mice exhibited impaired endothelium-dependent relaxation from acetylcholine (150). An interesting study demonstrated that nitric oxide can activate Per1 gene expression through a cAMP response element binding protein (cREB) and CLOCK/BMAL1-dependent mechanism (66). In the same study it was also demonstrated that age-dependent decreases in nitric oxide were related to impaired circadian rhythmicity. It remains to be determined whether or not vascular defects occur in other circadian KO mouse models.
Circadian clock and the kidney: role of coordinate regulation of sodium reabsorption by Per1.
Multiple renal functions exhibit a circadian rhythm including renal blood flow, glomerular filtration rate, potassium excretion, and sodium excretion (reviewed in Ref. 134). CLOCK KO mice have a blunting of the 24-h urinary sodium excretion pattern (157). The Na+/H+ exchanger 3, involved in sodium reabsorption, was shown to exhibit circadian oscillations in the mouse renal medulla (96, 120).
Our lab has shown that Per1 is involved in the basal and aldosterone-induced regulation of the α-subunit of the epithelial sodium channel (ENaC) in the renal collecting duct (47, 48, 116). ENaC is the key mediator of sodium reabsorption in the renal collecting duct and is responsible for the fine-tuning of sodium reabsorption and blood volume control by the kidney. In addition to ENaC, we have shown that Per1 coordinately regulates multiple genes involved in sodium reabsorption (135). These include the positive modulation of Fxyd5, a positive regulator of Na-K-ATPase (80). Fxyd5, a γ-like subunit of the sodium potassium ATPase, increases Na-K-ATPase activity. Per1 was shown to negatively regulate multiple genes involved in negative regulation of ENaC including Ube2e3, caveolin-1, and endothelin-1. Ube2e3 is an ubiquitin ligase that has been shown to inhibit ENaC by ubiquitination, targeting ENaC to the proteasome (27). Caveolin-1 is a lipid raft protein involved in retrieval of ENaC from the membrane (71). Increased caveolin-1 levels result in lower levels of ENaC in the membrane. Endothelin-1 is discussed above. Taken together, these results predict that Per1 KO would result in decreased sodium reabsorption and decreased blood pressure. As mentioned previously, Per1 KO mice have significantly lower blood pressure compared with WT controls. Interestingly, one human study showed that Per1 was upregulated 1.7-fold in the renal medulla of kidneys from hypertensive patients compared with normotensive controls, suggesting a role for Per1 in blood pressure control in humans (84).
Metabolism
The role of the circadian clock in metabolism has been studied extensively. Feeding cues can synchronize the peripheral clocks of the liver, heart, pancreas, and kidney (26, 154). The exact molecular mechanism is not fully understood, but evidence links the role of Sirtuin-1 (SIRT1), a NAD-dependent deacetylase, and the metabolite NAD, presumably through interactions of SIRT1 with CLOCK-BMAL1 (110, 118) (reviewed in Ref. 54). There is also evidence that the leptin-sensitive neurons of the arcuate nucleus (in the hypothalamus of the brain) are important for these cues (74). In this study, leptin receptor-B expressing neurons in the arcuate nucleus were targeted with a leptin-conjugated saporin toxin. Mice that were targeted with toxin rapidly became obese and experienced attenuation of feeding rhythms. Interestingly, SCN-ablated mice, in which the central clock is removed, exhibited arrhythmia in total darkness but did not become obese during the study. This study highlights the importance of the leptin-sensitive neurons of the arcuate nucleus not only in generation of feeding rhythms but also in obesity. It has been shown by transcriptomic analysis that a multitude of rate-limiting enzymes involved in metabolism exhibit circadian oscillations (133). Two studies, one in humans (24) and one in mice (35), show that an average of 25% of metabolites display circadian oscillations. The role of the circadian clock and metabolism has been heavily investigated in the last decade. The readers are directed to these two reviews for more information (9, 117). Recent work has focused on the role of the circadian clock in the pathophysiological conditions of obesity and diabetes.
Circadian clock in obesity: story of REV-ERB and CLOCK.
One of the first experimental models to highlight the importance of the circadian clock in metabolism was the CLOCK mutant mouse (Δ19). CLOCK (Δ19) mutant animals exhibit metabolic syndrome with obesity and hyperlipidemia (143). Period 1/2/3 triple KO or Per3 single KO mice gain significantly more weight on normal or high-fat diets than WT mice (25). One study demonstrated that BMAL1 KO animals are obese and exhibit hyperlipidemia (49). However, another group reported that BMAL1 KO animals do not have this phenotype, and that the lack of weight gain was attributed to the aging phenotype seen in these animals (51). Additional studies are needed to resolve the role of BMAL1 in these disorders. Overweight women with a single-nucleotide-polymorphism (SNP) in the Clock gene locus (3111T/C) exhibited decreased activity, altered sleep patterns, and abnormal circadian rhythmicity (8). It remains to be seen whether this SNP in the Clock gene causes a propensity for obesity.
Two recent studies showed that restriction of food intake to certain times of day can attenuate the weight gain that occurs due to a high-fat diet. One study restricted feeding in mice to the inactive period (day), thereby reversing the feeding rhythm without changing total caloric intake. This study showed that the time-of-day restricted animals had attenuated weight gain, lower adipogenesis, lower cholesterol, lower corticosterone, and were more insulin sensitive than control mice that had free access to food over the 24-h cycle (124). The increased level of these factors was attributed to the high-fat diet. Another study restricted feeding to the mouse active period (night). The time-of-day restricted mice experienced attenuated weight gain, higher oxygen consumption as a measure of metabolic rate, increased leptin, and insulin sensitivity (50). Thus, in both cases, food restriction to either the active or inactive period led to attenuation of high-fat diet-induced obesity. In the case of humans, these results could be applicable to people with night-time eating syndrome, in which sufferers eat constantly night or day, leading to a greater risk of metabolic syndrome (reviewed in Ref. 44).
In recent years, focus has turned to the circadian proteins CLOCK and REV-ERB-α in the circadian control of metabolism. As mentioned above, CLOCK (Δ19) mutant display a metabolic syndrome phenotype. Recent studies showed that REV-ERB-α KO animals have increased adiposity and deregulated fatty acid/glucose utilization (28). Another interesting study, using genome-wise cistromic analysis to identify target sites in mouse liver, demonstrated that REV-ERB-α and REV-ERB-β share 50% common DNA binding sites and display significant overlap with BMAL1 binding sites, including those critical for metabolism (18). As mentioned above, it has been shown that REV-ERB-α plays a role in inhibition of BMAL1 expression. This study presents an important role for REV-ERB-β and suggests a novel mechanism for REV-ERB-α/β regulation of metabolic genes. These researchers also demonstrated that REV-ERB-α/β double KO mice display deregulated lipid metabolism. Another study showed that synthetic REV-ERB-α/β agonists led to attenuation of weight gain, cholesterol levels, triglyceride levels, and lower fat mass in the obese ob/ob mice (127). Both of these studies implicate REV-ERB-α in the regulation of metabolism and highlight the possibility of targeting REV-ERB-α for the treatment of obesity.
Circadian clock and diabetes: glucose-stimulated insulin secretion, uncoupling protein 2, and Cry 1/2.
Studies in circadian clock gene KO mice suggest a role for the circadian clock proteins in diabetes. BMAL1 and CLOCK KO mouse are diabetic with reduced plasma insulin levels (83). Circadian desychronization caused faster onset of diabetes in the Type 1 diabetic HIP rat model; these rats overexpress the human islet amyloid polypeptide, which leads to pancreatic β cell failure (43). One study demonstrated that two SNPs in the Bmal1 gene were associated with an increased risk for gestational diabetes mellitus in Greek women (102). Transcriptomic analysis of pancreatic islets from healthy and Type 2 diabetics demonstrated that Per2–3 and Cry2 were downregulated in people with Type 2 diabetes (132).
BMAL1 KO mice have impaired glucose-stimulated insulin secretion (GSIS), which may explain the reduced plasma insulin levels in these animals. The reduced GSIS may be due to upregulation of the mitochondrial uncoupling protein 2 (UCP2) (72). Interestingly, UCP2 KO animals exhibit endothelial dysfunction (141). This is similar to BMAL1 KO animals, but conversely, UCP2 is upregulated in BMAL1 KO. The full relationship between UCP2 and BMAL1 remains to be explored and will be of great interest. Pancreas-specific BMAL1 KO mice, in which CRE recombinase is driven by the PDX1 promoter, exhibited severe hyperglycemia (83). This study also demonstrated that a defect in insulin exocytosis was a potential cause for the decreased insulin in these animals. Knockdown of REV-ERB-α in mouse islet cells resulted in impaired GSIS along with decreased fatty acid synthase and decreased sterol regulatory element-binding protein 1-c (SREBP-1c) (148). Both of these proteins are critical for fat metabolism.
Several studies have looked at the effect of central clock loss on insulin resistance by ablating the SCN in mice and rats. SCN ablation in rats resulted in the loss of the circadian rhythm of glucose uptake and insulin sensitivity (67). Constant light exposure, which will disrupt the circadian clock (57, 97), resulted in abolishment of the circadian rhythmicity of insulin sensitivity in mice (21). SCN ablation in mice led to a similar disruption in the rhythm of insulin sensitivity and led to hepatic insulin resistance (22). These studies demonstrate the importance of the central clock in the control of insulin resistance.
Recent work has illustrated a role for Cry1 and Cry2 in diabetes. Cry1/2 KO mice were hyperglycemic (140). Furthermore, overexpression of Cry1 in diabetic db/db mice resulted in lowering of blood glucose and increased insulin sensitivity (156). Another study, using a synthetic Cry1/2 agonist in mouse primary hepatocytes, showed that activation of Cry1/2 led to increased glucose production. It was demonstrated that this result was most likely due to increased synthesis of two rate-limiting gluconeogenic enzymes Pck1 and G6pc (52). It will be interesting to see if Cry1/2 is involved in the regulation of GSIS, but as mentioned above, it has recently been shown in genome wide ChIP-seq studies that Cry1/2 has a multitude of “CLOCK independent interactions” with response elements outside of the classical E-box/CLOCK/BMAL1 complex (62). Further investigation into the physiological significance of these interactions should help determine the feasibility of targeting the clock clinically for the treatment of diabetes and other metabolic disorders.
Endocrine System
The circadian clock plays a key role in the regulation of the endocrine system, but in turn, the endocrine system plays an important role in the synchronization and regulation of the peripheral clocks (discussed in Ref. 117). Glucocorticoids and melatonin have been implicated in the circadian clock mechanism, both in the regulation of the endocrine system and the peripheral clocks.
Circadian clock and glucocorticoids: circadian synchronizer?
The glucocorticoids, a group of steroid hormones synthesized in the adrenal cortex, are involved in mediating anti-inflammation, the stress response, metabolism, cardiovascular, and neurological function (a more in-depth discussion of the role of these steroids in Ref. 20). One of the proposed roles of the glucocorticoids is in synchronization of the circadian clock. Multiple studies have shown that treatment with the synthetic glucocorticoid dexamethasone (DEX) can synchronize cell cultures in vitro through the synchronization of circadian gene expression (7, 128). DEX can also synchronize isolated cultured mice atrium (146). It was also shown that hydrocortisone, a glucocorticoid, can synchronize circadian clock gene expression in ex vivo mouse SCN, lung, pineal gland, salivary gland, cornea, and liver (105). Adrenalectomized rats maintain rhythmic oscillation of most clock genes in the liver and kidney, except Per1 (137). The fact that the other clock genes still oscillate suggests that glucocorticoids are not the only synchronization mechanism for the circadian clock. It is not surprising that Per1 is affected by adrenalectomy as it has been shown that Per1 is highly sensitive to glucocorticoid signaling (113). However, on the note of increased clock gene expression, several studies performed in multiple cell types showed changes in clock gene expression with DEX including: increased Per1–3 and Bmal1 expression in osteoblast cells (63), increased Per1 in neutrophils, increased Per1–2 in lymphocytes (94), and increased Per1 in renal inner medullary collecting duct cells (136). Mechanistically, one convincing study showed that synchronization in the liver due to DEX was at least partially mediated by the transcription factor hepatocyte nuclear factor 4α (HNF4α) (112).
Glucocorticoids interact with the nuclear hormone receptor, the glucocorticoid receptor (GR). A recent study showed that Cry1/2 interacted with and repressed the actions of GR (68). It was also shown that Cry1/2 are involved in the modulation of DEX-controlled genes, presumably through repression of GR. As mentioned above, Cry1/2 appear to have “CLOCK-independent” interactions, which are interactions that do not involve the CLOCK-BMAL1 complex. Interestingly, a multitude of these seem to be binding events with hormone response elements, including the glucocorticoid response element, which is the binding site of the glucocorticoid receptor (62).
Circadian clock and melatonin: diabetes connection.
Melatonin is a hormone secreted from the pineal gland and other tissues, which is controlled by light. Melatonin is only secreted in darkness (reviewed in Ref. 14). Its levels are not affected by activity and are secreted at darkness for both diurnal and nocturnal animals. It behaves as a feedback mechanism for the central clock in the SCN by inhibiting neuron firing (76) (reviewed in Ref. 108). It also has been implicated in synchronization of the central and perhaps peripheral clocks (106, 114) (reviewed in Ref. 14). Melatonin acts through its two receptors, melatonin receptor 1 and 2, which were recently shown to be expressed in a circadian pattern and to be regulated by melatonin levels (147).
There appears to be a role for melatonin-mediated inhibition of insulin secretion, with a suggested role for melatonin in the control of GSIS (reviewed in Ref. 104). Recent work has begun to determine if there is a connection between melatonin signaling and diabetes. Per1 KO mice exhibit higher plasma and pineal melatonin concentrations during the active phase (19). It will be of great interest to see if Per1 KO mice display reduced insulin secretion due to increased melatonin. It was also shown that removal of the melatonin 2 receptor results in decreased Per1 and Cry1 expression in the SCN (106). Whether these two phenomena are connected remains to be determined. One study showed that melatonin treatment of human prostate cancer cells resulted in increased expression of Per1 and CLOCK, but decreased BMAL1 levels (56). However, the effect of melatonin treatment on circadian protein levels in a noncancerous tissue has not been studied; this is important given that melatonin may have anticarcinogenic effects (reviewed in Ref. 12). If CLOCK or BMAL1 knockdown resulted in upregulation of melatonin, it would provide evidence that the inhibitory actions of melatonin could participate in the lowered insulin levels in these KO animals. A recent human study using three groups of men (lean, obese nondiabetic, and obese Type 2 diabetic) showed that obese nondiabetic men had higher plasma melatonin levels than either lean or diabetic obese men (82). They also had significantly higher plasma leptin levels, but the physiological significance of these data in the context of the circadian clock remain to be seen. Further evidence for a role for melatonin in diabetes comes from two studies that independently identified a SNP near the melatonin receptor 1B gene, which encodes the melatonin 2 receptor, that is associated with increased fasting glucose and increased incidence of Type 2 diabetes in White Europeans and Asians (10, 77). More investigation is required to fully flesh out the connection between melatonin, diabetes, and the circadian clock. Regardless, these studies have highlighted the importance for the circadian clock and melatonin in glucose control and diabetes.
Immune System
Over the last few years, investigation into the circadian clock in the immune system has increased. Glucocorticoids (40) and melatonin (109) are critical regulators of both the circadian clock and immune system.
Circadian clock and immune system: TLR9, CLOCK, and NF-κβ.
Much of the early work in the circadian clock and the immune system involved identifying the circadian oscillation of multiple immune cell types such as T-cells, granulocytes, and lymphocyte levels in humans (1, 59, 88). It was not until recently, however, that the more molecular basis of the circadian regulation in the immune system was uncovered. One significant study looked at the circadian clock in innate immunity and showed that toll-like receptor 9 (TLR9) was a circadian target gene. The TLRs are critical for innate immunity and are responsible for the recognition of foreign pathogens. Once a TLR recognizes a foreign pathogen, it activates the innate immune response. TLR9 recognizes unmethylated CpG dinucleotides, which are found commonly in viruses and bacteria. In this study, Per2 (BRDm1) mutant animals were studied. This mouse model involves a deletion of the interaction domain of Per2, rendering Per2 unable to interact with other circadian clock proteins. In these animals, TLR9 was upregulated in whole spleen, but downregulated in splenic macrophages. TLR9 was also downregulated in peritoneal macrophages following CLOCK small interfering RNA (siRNA) treatment in vitro. In a mouse model of sepsis, Per2 mutant mice had increased sepsis severity that corresponded to the peak time of TLR9 expression (125).
T-cell response was shown to be regulated by the circadian clock. T-cell antigen response was decreased in CLOCK KO animals, most likely due to downregulation of the T-cell signaling kinase ZAP70 (38). They also showed that mice immunized during the inactive period had stronger T-cell response than mice immunized during the active period. Another study demonstrated that natural killer cell activity was decreased in a rat model of chronic jet lag (78). An interesting study demonstrated that Cry1/2 KO fibroblasts show increased pro-inflammatory cytokine production, with upregulation of nuclear factor (NF)-κB, a key transcription factor in the immune response (93). One recent study showed that CLOCK positively modulates NF-κB in a possible “circadian clock-independent fashion” that does not involve the transcriptional activity of CLOCK-BMAL1. This is suggested due to the effects still being seen in CLOCK (Δ19) mutant mice (129). This is one of the few studies that identify CLOCK having effects that do not involve the classical interaction with the E-box; if more of these interactions exist will be of great interest.
Reproductive System
Recent research has contributed to our understanding of the molecular circadian clock in the reproductive system. Much of the work on the circadian clock in the reproductive system has been done by investigating the role of the central clock in the female reproductive cycle. Less is known about the testicular clock and until lately, it was debated whether a testicular clock even existed (60, 91); however, new studies have shed light on the testicular clock.
Circadian clock and female reproductive system: gonadotropin-releasing peptide hormone and the central clock.
A focus of studies in the female reproductive clock has been the involvement of the gonadotropin-releasing peptide hormone (GnRH) neurons. GnRH is responsible for the release of follicle stimulating hormone and luteinizing hormone from the anterior pituitary gland. Release of GnRH and GnRH neuron activity is known to display a circadian pattern (81). For more detailed information on the role of the GnRH neurons and the circadian clock, the reader is directed to an excellent review (15). Transient expression of the dominant negative CLOCK (Δ19) protein in the GnRH-secreting GT1–7 cell line resulted in decreased secretion of GnRH (16). These cells were demonstrated to have a functional circadian clock, as measured by oscillating expression of the core circadian genes (46, 98). Interestingly, it was shown that GnRH induced the expression of Per1, but not Per2, through a protein kinase C-dependent pathway(99). Tissue-specific KO of the clock genes in the reproductive system may shed additional light on the physiological significance of these observations.
In the periphery, it is known that the clock genes are expressed and oscillate in the ovaries of mice, rats, and bovines (reviewed in Ref. 122). One study showed that the dorsomedial SCN is responsible for the circadian control of Kiss1 (involved in inducing ovulation) and the luteinizing hormone surge (126). SCN ablation studies showed that the SCN was necessary for the circadian control of ovulation and estrous cycles in rats (reviewed in Ref. 122). These findings demonstrate the importance of the central SCN clock for the circadian control of the peripheral ovarian clock. Female BMAL1 KO mice have decreased fertility with delayed estrous cycles and implantation defects (111). Decreased fertility and increased duration of estrous cycles were observed in CLOCK KO mice (16). Interestingly, a recent study demonstrated the existence of the circadian clock in the placenta (151), but the physiological significance of this observation remains to be seen.
Circadian clock and male reproductive system: beyond the 24-h clock.
Circadian clock gene expression has been shown to be expressed in a spermatogenesis stage-dependent manner (103). CLOCK and BMAL1 proteins were shown to be localized to the chromatoid body (CB), a cytoplasmic organelle found in the male germ cell (reviewed in Ref. 64). The exact physiological significance of this is unknown; however, BMAL1 KO spermatids displayed morphological deformities.
One of the more intriguing concepts of the circadian clock in the reproductive system is the presumably non-24 h reproductive cycle-dependent clock, specifically shown in the males (103) but perhaps occurs also in females. It brings another dimension to the understanding of the clock beyond 24-h cycles, but to 24-h cycles that are dependent on the reproductive cycle itself. The exact extent these non-24-h clocks play in the reproductive system remain to be seen.
In conclusion, the circadian clock plays an integral role in the regulation of a variety of physiological functions (Fig. 2). Future studies may reveal previously unrecognized roles for the circadian clock proteins in disease states beyond those discussed here. One of the more intriguing phenomena in the circadian clock field is the concept of integrated regulation. It is common knowledge that cross-talk occurs between physiological systems, such as that between the gut and the liver. The circadian clock provides an additional level of integration that is now more fully appreciated by researchers across multiple disciplines.
Fig. 2.
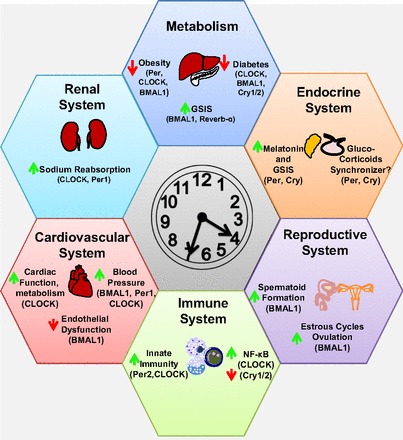
Integrative role for the circadian clock in the regulation of physiological function. Circadian proteins listed in parentheses dictate which proteins have been implicated in regulating the process in question. If a protein is not listed, it does not imply that it is not involved, but that it has not yet been tested. Green arrows represent induction by the circadian protein; red arrows represent repression. The time shown on the clock is for illustrative purposes only.
Another theme that has become apparent in recent years is the finding that the circadian clock proteins participate in “CLOCK-independent” activities, such as that between Cry1/2 and GR or CLOCK and NF-κβ. This does not mean the effects are time independent but that the action of one circadian protein is not dependent on the full participation of the other core circadian clock proteins. If any of the other circadian proteins participate in these types of interactions remains to be seen, and the outcome of such studies is likely to be of great interest to many biomedical disciplines. As mentioned above, genome-wide ChIP-seq analysis of the circadian proteins was recently performed in mouse liver samples over the 24-h cycle in mice (62). It will be extremely informative for studies such as these to be done in additional tissues to fully understand the role of circadian clock proteins in physiology.
Perspectives and Significance
A major purpose of the circadian clock is to increase the activity of a given physiological system when needed and decrease the system activity when it is not needed. This provides great power to the circadian clock but also can cause great detriment when things go wrong. The studies discussed in this review highlight the large impact of a dysregulated circadian clock. Though many of these findings stem from studies using global KO mice, cell-type and tissue-specific KO models are increasingly being used. The CCM mouse and the pancreas-specific BMAL1 KO mouse are examples that have increased our understanding of how the clock functions in these tissues. The generation of additional tissue-specific circadian gene KOs should greatly benefit the circadian physiology field. The use of constant light or circadian disruption with a 10 h-10 h light-dark cycle allows the generation of a dysregulated circadian clock in a WT animal. This model is presumably similar to circadian disruption observed in humans due to chronic jet lag or rotating shift work. It will be interesting to see if the_phenotypes that were discovered using genetic knockouts can be recapitulated in models of circadian disruption.
The findings reviewed here from multiple physiological systems provide support for consideration of the circadian clock as a potential new drug target in a variety of disorders. The circadian clock plays a key role in multiple physiological systems and thus might be an attractive target for diseases that involve multiple organ systems. As discussed above, researchers are increasingly experimenting with pharmacologically targeting the clock, such as the use of Cry1/2 agonists in diabetes or REV-ERB agonists in the treatment of obesity. Additional studies exploring pharmacological modulation of the clock are needed. It is quite apparent that the circadian clock plays an important role in physiology, and only time will tell to what extent manipulation of this powerful, yet delicate system, may aid in the remedy of pathophysiological conditions.
REFERENCES
- 1.Ackermann K, Revell VL, Lao O, Rombouts EJ, Skene DJ, Kayser M. Diurnal rhythms in blood cell populations and the effect of acute sleep deprivation in healthy young men. Sleep 35: 933–940, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Agarwal R. Regulation of circadian blood pressure: from mice to astronauts. Curr Opin Nephrol Hypertens 19: 51–58, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Albrecht U. The mammalian circadian clock: a network of gene expression. Front Biosci 9: 48–55, 2004 [DOI] [PubMed] [Google Scholar]
- 4.Albrecht U, Eichele G. The mammalian circadian clock. Curr Opin Genet Dev 13: 271–277, 2003 [DOI] [PubMed] [Google Scholar]
- 5.Anea CB, Cheng B, Sharma S, Kumar S, Caldwell RW, Yao L, Ali MI, Merloiu AM, Stepp DW, Black SM, Fulton DJ, Rudic RD. Increased superoxide and endothelial NO synthase uncoupling in blood vessels of Bmal1-knockout mice. Circ Res 111: 1157–1165, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ayala DE, Moya A, Crespo JJ, Castineira C, Dominguez-Sardina M, Gomara S, Sineiro E, Mojon A, Fontao MJ, Hermida RCHygia Project Investigators. Circadian pattern of ambulatory blood pressure in hypertensive patients with and without Type 2 diabetes. Chronobiol Int 30: 99–115, 2012 [DOI] [PubMed] [Google Scholar]
- 7.Balsalobre A, Brown SA, Marcacci L, Tronche F, Kellendonk C, Reichardt HM, Schutz G, Schibler U. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science 289: 2344–2347, 2000 [DOI] [PubMed] [Google Scholar]
- 8.Bandin C, Martinez-Nicolas A, Ordovas JM, Ros Lucas JA, Castell P, Silvente T, Madrid JA, Garaulet M. Differences in circadian rhythmicity in CLOCK 3111T/C genetic variants in moderate obese women as assessed by thermometry, actimetry and body position. Int J Obes (Lond) Nov 27. 10.1038/ijo.2012.180 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bass J, Takahashi JS. Circadian integration of metabolism and energetics. Science 330: 1349–1354, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bouatia-Naji N, Bonnefond A, Cavalcanti-Proenca C, Sparso T, Holmkvist J, Marchand M, Delplanque J, Lobbens S, Rocheleau G, Durand E, De Graeve F, Chevre JC, Borch-Johnsen K, Hartikainen AL, Ruokonen A, Tichet J, Marre M, Weill J, Heude B, Tauber M, Lemaire K, Schuit F, Elliott P, Jorgensen T, Charpentier G, Hadjadj S, Cauchi S, Vaxillaire M, Sladek R, Visvikis-Siest S, Balkau B, Levy-Marchal C, Pattou F, Meyre D, Blakemore AI, Jarvelin MR, Walley AJ, Hansen T, Dina C, Pedersen O, Froguel P. A variant near MTNR1B is associated with increased fasting plasma glucose levels and type 2 diabetes risk. Nat Genet 41: 89–94, 2009 [DOI] [PubMed] [Google Scholar]
- 11.Bugaj V, Pochynyuk O, Mironova E, Vandewalle A, Medina JL, Stockand JD. Regulation of the epithelial Na+ channel by endothelin-1 in rat collecting duct. Am J Physiol Renal Physiol 295: F1063–F1070, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bukowska A. [Anticarcinogenic role of melatonin–potential mechanisms]. Med Pr 62: 425–434, 2011 [PubMed] [Google Scholar]
- 13.Centonza L, Castoldi G, Chianca R, Busca G, Golin R, Zanchetti A, Stella A. Short-term analysis of the relationship between blood pressure and urinary sodium excretion in normotensive subjects. Clin Sci (Lond) 98: 495–500, 2000 [PubMed] [Google Scholar]
- 14.Challet E. Minireview: Entrainment of the suprachiasmatic clockwork in diurnal and nocturnal mammals. Endocrinology 148: 5648–5655, 2007 [DOI] [PubMed] [Google Scholar]
- 15.Chappell PE, Goodall CP, Tonsfeldt KJ, White RS, Bredeweg E, Latham KL. Modulation of gonadotrophin-releasing hormone secretion by an endogenous circadian clock. J Neuroendocrinol 21: 339–345, 2009 [DOI] [PubMed] [Google Scholar]
- 16.Chappell PE, White RS, Mellon PL. Circadian gene expression regulates pulsatile gonadotropin-releasing hormone (GnRH) secretory patterns in the hypothalamic GnRH-secreting GT1–7 cell line. J Neurosci 23: 11202–11213, 2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cheng B, Anea CB, Yao L, Chen F, Patel V, Merloiu A, Pati P, Caldwell RW, Fulton DJ, Rudic RD. Tissue-intrinsic dysfunction of circadian clock confers transplant arteriosclerosis. Proc Natl Acad Sci USA 108: 17147–17152, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cho H, Zhao X, Hatori M, Yu RT, Barish GD, Lam MT, Chong LW, Ditacchio L, Atkins AR, Glass CK, Liddle C, Auwerx J, Downes M, Panda S, Evans RM. Regulation of circadian behaviour and metabolism by REV-ERB-α and REV-ERB-β. Nature 485: 123–127, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Christ E, Pfeffer M, Korf HW, von Gall C. Pineal melatonin synthesis is altered in Period1 deficient mice. Neuroscience 171: 398–406, 2010 [DOI] [PubMed] [Google Scholar]
- 20.Chung S, Son GH, Kim K. Adrenal peripheral oscillator in generating the circadian glucocorticoid rhythm. Ann NY Acad Sci 1220: 71–81, 2011 [DOI] [PubMed] [Google Scholar]
- 21.Coomans CP, van den Berg SA, Houben T, van Klinken JB, van den Berg R, Pronk AC, Havekes LM, Romijn JA, Willems van Dijk K, Biermasz NR, Meijer JH. Detrimental effects of constant light exposure and high-fat diet on circadian energy metabolism and insulin sensitivity. FASEB J 27: 1721–1732, 2013 [DOI] [PubMed] [Google Scholar]
- 22.Coomans CP, van den Berg SA, Lucassen EA, Houben T, Pronk AC, van der Spek RD, Kalsbeek A, Biermasz NR, Willems van Dijk K, Romijn JA, Meijer JH. The suprachiasmatic nucleus controls circadian energy metabolism and hepatic insulin sensitivity. Diabetes 62: 1102–1108, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Curtis AM, Cheng Y, Kapoor S, Reilly D, Price TS, Fitzgerald GA. Circadian variation of blood pressure and the vascular response to asynchronous stress. Proc Natl Acad Sci USA 104: 3450–3455, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dallmann R, Viola AU, Tarokh L, Cajochen C, Brown SA. The human circadian metabolome. Proc Natl Acad Sci USA 109: 2625–2629, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dallmann R, Weaver DR. Altered body mass regulation in male mPeriod mutant mice on high-fat diet. Chronobiol Int 27: 1317–1328, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Damiola F, Le Minh N, Preitner N, Kornmann B, Fleury-Olela F, Schibler U. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev 14: 2950–2961, 2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Debonneville C, Staub O. Participation of the ubiquitin-conjugating enzyme UBE2E3 in Nedd4–2-dependent regulation of the epithelial Na+ channel. Mol Cell Biol 24: 2397–2409, 2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Delezie J, Dumont S, Dardente H, Oudart H, Grechez-Cassiau A, Klosen P, Teboul M, Delaunay F, Pevet P, Challet E. The nuclear receptor REV-ERBalpha is required for the daily balance of carbohydrate and lipid metabolism. FASEB J 26: 3321–3335, 2012 [DOI] [PubMed] [Google Scholar]
- 29.Dibner C, Schibler U, Albrecht U. The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annu Rev Physiol 72: 517–549, 2010 [DOI] [PubMed] [Google Scholar]
- 30.Doi M, Takahashi Y, Komatsu R, Yamazaki F, Yamada H, Haraguchi S, Emoto N, Okuno Y, Tsujimoto G, Kanematsu A, Ogawa O, Todo T, Tsutsui K, van der Horst GT, Okamura H. Salt-sensitive hypertension in circadian clock-deficient Cry-null mice involves dysregulated adrenal Hsd3b6. Nat Med 16: 67–74, 2010 [DOI] [PubMed] [Google Scholar]
- 31.Durgan DJ, Pulinilkunnil T, Villegas-Montoya C, Garvey ME, Frangogiannis NG, Michael LH, Chow CW, Dyck JR, Young ME. Short communication: ischemia/reperfusion tolerance is time-of-day-dependent: mediation by the cardiomyocyte circadian clock. Circ Res 106: 546–550, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Durgan DJ, Trexler NA, Egbejimi O, McElfresh TA, Suk HY, Petterson LE, Shaw CA, Hardin PE, Bray MS, Chandler MP, Chow CW, Young ME. The circadian clock within the cardiomyocyte is essential for responsiveness of the heart to fatty acids. J Biol Chem 281: 24254–24269, 2006 [DOI] [PubMed] [Google Scholar]
- 33.Durgan DJ, Young ME. The cardiomyocyte circadian clock: emerging roles in health and disease. Circ Res 106: 647–658, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Dyer AR, Martin GJ, Burton WN, Levin M, Stamler J. Blood pressure and diurnal variation in sodium, potassium, and water excretion. J Hum Hypertens 12: 363–371, 1998 [DOI] [PubMed] [Google Scholar]
- 35.Eckel-Mahan KL, Patel VR, Mohney RP, Vignola KS, Baldi P, Sassone-Corsi P. Coordination of the transcriptome and metabolome by the circadian clock. Proc Natl Acad Sci USA 109: 5541–5546, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Eckle T, Hartmann K, Bonney S, Reithel S, Mittelbronn M, Walker LA, Lowes BD, Han J, Borchers CH, Buttrick PM, Kominsky DJ, Colgan SP, Eltzschig HK. Adora2b-elicited Per2 stabilization promotes a HIF-dependent metabolic switch crucial for myocardial adaptation to ischemia. Nat Med 18: 774–782, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fabbian F, Smolensky MH, Tiseo R, Pala M, Manfredini R, Portaluppi F. Dipper and non-dipper blood pressure 24-hour patterns: circadian rhythm-dependent physiologic and pathophysiologic mechanisms. Chronobiol Int 30: 17–30, 2013 [DOI] [PubMed] [Google Scholar]
- 38.Fortier EE, Rooney J, Dardente H, Hardy MP, Labrecque N, Cermakian N. Circadian variation of the response of T cells to antigen. J Immunol 187: 6291–6300, 2011 [DOI] [PubMed] [Google Scholar]
- 39.Fournier S, Eeckhout E, Mangiacapra F, Trana C, Lauriers N, Beggah AT, Monney P, Cook S, Bardy D, Vogt P, Muller O. Circadian variations of ischemic burden among patients with myocardial infarction undergoing primary percutaneous coronary intervention. Am Heart J 163: 208–213, 2012 [DOI] [PubMed] [Google Scholar]
- 40.Franchimont D. Overview of the actions of glucocorticoids on the immune response: a good model to characterize new pathways of immunosuppression for new treatment strategies. Ann NY Acad Sci 1024: 124–137, 2004 [DOI] [PubMed] [Google Scholar]
- 41.Fujino Y, Iso H, Tamakoshi A, Inaba Y, Koizumi A, Kubo T, Yoshimura T. A prospective cohort study of shift work and risk of ischemic heart disease in Japanese male workers. Am J Epidemiol 164: 128–135, 2006 [DOI] [PubMed] [Google Scholar]
- 42.Fukuda M, Wakamatsu-Yamanaka T, Mizuno M, Miura T, Tomonari T, Kato Y, Ichikawa T, Miyagi S, Shirasawa Y, Ito A, Yoshida A, Kimura G. Angiotensin receptor blockers shift the circadian rhythm of blood pressure by suppressing tubular sodium reabsorption. Am J Physiol Renal Physiol 301: F953–F957, 2011 [DOI] [PubMed] [Google Scholar]
- 43.Gale JE, Cox HI, Qian J, Block GD, Colwell CS, Matveyenko AV. Disruption of circadian rhythms accelerates development of diabetes through pancreatic beta-cell loss and dysfunction. J Biol Rhythms 26: 423–433, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gallant AR, Lundgren J, Drapeau V. The night-eating syndrome and obesity. Obes Rev 13: 528–536, 2012 [DOI] [PubMed] [Google Scholar]
- 45.Gekakis N, Staknis D, Nguyen HB, Davis FC, Wilsbacher LD, King DP, Takahashi JS, Weitz CJ. Role of the CLOCK protein in the mammalian circadian mechanism. Science 280: 1564–1569, 1998 [DOI] [PubMed] [Google Scholar]
- 46.Gillespie JM, Chan BP, Roy D, Cai F, Belsham DD. Expression of circadian rhythm genes in gonadotropin-releasing hormone-secreting GT1–7 neurons. Endocrinology 144: 5285–5292, 2003 [DOI] [PubMed] [Google Scholar]
- 47.Gumz ML, Cheng KY, Lynch IJ, Stow LR, Greenlee MM, Cain BD, Wingo CS. Regulation of alphaENaC expression by the circadian clock protein Period 1 in mpkCCD(c14) cells. Biochim Biophys Acta 1799: 622–629, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Gumz ML, Stow LR, Lynch IJ, Greenlee MM, Rudin A, Cain BD, Weaver DR, Wingo CS. The circadian clock protein Period 1 regulates expression of the renal epithelial sodium channel in mice. J Clin Invest 119: 2423–2434, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Guo B, Chatterjee S, Li L, Kim JM, Lee J, Yechoor VK, Minze LJ, Hsueh W, Ma K. The clock gene, brain and muscle Arnt-like 1, regulates adipogenesis via Wnt signaling pathway. FASEB J 26: 3453–3463, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hatori M, Vollmers C, Zarrinpar A, DiTacchio L, Bushong EA, Gill S, Leblanc M, Chaix A, Joens M, Fitzpatrick JA, Ellisman MH, Panda S. Time-restricted feeding without reducing caloric intake prevents metabolic diseases in mice fed a high-fat diet. Cell Metab 15: 848–860, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Hemmeryckx B, Himmelreich U, Hoylaerts MF, Lijnen HR. Impact of clock gene Bmal1 deficiency on nutritionally induced obesity in mice. Obesity (Silver Spring) 19: 659–661, 2011 [DOI] [PubMed] [Google Scholar]
- 52.Hirota T, Lee JW, St John PC, Sawa M, Iwaisako K, Noguchi T, Pongsawakul PY, Sonntag T, Welsh DK, Brenner DA, Doyle FJ, 3rd, Schultz PG, Kay SA. Identification of small molecule activators of cryptochrome. Science 337: 1094–1097, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Hossmann V, Fitzgerald GA, Dollery CT. Circadian rhythm of baroreflex reactivity and adrenergic vascular response. Cardiovasc Res 14: 125–129, 1980 [DOI] [PubMed] [Google Scholar]
- 54.Imai S. “Clocks” in the NAD World: NAD as a metabolic oscillator for the regulation of metabolism and aging. Biochim Biophys Acta 1804: 1584–1590, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Jeyaraj D, Haldar SM, Wan X, McCauley MD, Ripperger JA, Hu K, Lu Y, Eapen BL, Sharma N, Ficker E, Cutler MJ, Gulick J, Sanbe A, Robbins J, Demolombe S, Kondratov RV, Shea SA, Albrecht U, Wehrens XH, Rosenbaum DS, Jain MK. Circadian rhythms govern cardiac repolarization and arrhythmogenesis. Nature 483: 96–99, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Jung-Hynes B, Huang W, Reiter RJ, Ahmad N. Melatonin resynchronizes dysregulated circadian rhythm circuitry in human prostate cancer cells. J Pineal Res 49: 60–68, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kalsbeek A, Scheer FA, Perreau-Lenz S, La Fleur SE, Yi CX, Fliers E, Buijs RM. Circadian disruption and SCN control of energy metabolism. FEBS Lett 585: 1412–1426, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kaneko M, Zechman FW, Smith RE. Circadian variation in human peripheral blood flow levels and exercise responses. J Appl Physiol 25: 109–114, 1968 [DOI] [PubMed] [Google Scholar]
- 59.Kirsch S, Thijssen S, Alarcon Salvador S, Heine GH, van Bentum K, Fliser D, Sester M, Sester U. T-cell numbers and antigen-specific T-cell function follow different circadian rhythms. J Clin Immunol 32: 1381–1389, 2012 [DOI] [PubMed] [Google Scholar]
- 60.Klose M, Grote K, Lerchl A. Temporal control of spermatogenesis is independent of the central circadian pacemaker in Djungarian hamsters (Phodopus sungorus). Biol Reprod 84: 124–129, 2011 [DOI] [PubMed] [Google Scholar]
- 61.Ko GY, Shi L, Ko ML. Circadian regulation of ion channels and their functions. J Neurochem 110: 1150–1169, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Koike N, Yoo SH, Huang HC, Kumar V, Lee C, Kim TK, Takahashi JS. Transcriptional architecture and chromatin landscape of the core circadian clock in mammals. Science 338: 349–354, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Komoto S, Kondo H, Fukuta O, Togari A. Comparison of beta-adrenergic and glucocorticoid signaling on clock gene and osteoblast-related gene expressions in human osteoblast. Chronobiol Int 29: 66–74, 2012 [DOI] [PubMed] [Google Scholar]
- 64.Kotaja N, Bhattacharyya SN, Jaskiewicz L, Kimmins S, Parvinen M, Filipowicz W, Sassone-Corsi P. The chromatoid body of male germ cells: similarity with processing bodies and presence of Dicer and microRNA pathway components. Proc Natl Acad Sci USA 103: 2647–2652, 2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kraft M, Martin RJ. Chronobiology and chronotherapy in medicine. Dis Mon 41: 506–575, 1995 [DOI] [PubMed] [Google Scholar]
- 66.Kunieda T, Minamino T, Miura K, Katsuno T, Tateno K, Miyauchi H, Kaneko S, Bradfield CA, FitzGerald GA, Komuro I. Reduced nitric oxide causes age-associated impairment of circadian rhythmicity. Circ Res 102: 607–614, 2008 [DOI] [PubMed] [Google Scholar]
- 67.la Fleur SE, Kalsbeek A, Wortel J, Fekkes ML, Buijs RM. A daily rhythm in glucose tolerance: a role for the suprachiasmatic nucleus. Diabetes 50: 1237–1243, 2001 [DOI] [PubMed] [Google Scholar]
- 68.Lamia KA, Papp SJ, Yu RT, Barish GD, Uhlenhaut NH, Jonker JW, Downes M, Evans RM. Cryptochromes mediate rhythmic repression of the glucocorticoid receptor. Nature 480: 552–556, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Larochelle P. Circadian variation in blood pressure: dipper or nondipper. J Clin Hypertens (Greenwich) 4: 3–8, 2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Lee HM, Chen R, Kim H, Etchegaray JP, Weaver DR, Lee C. The period of the circadian oscillator is primarily determined by the balance between casein kinase 1 and protein phosphatase 1. Proc Natl Acad Sci USA 108: 16451–16456, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Lee IH, Campbell CR, Song SH, Day ML, Kumar S, Cook DI, Dinudom A. The activity of the epithelial sodium channels is regulated by caveolin-1 via a Nedd4–2-dependent mechanism. J Biol Chem 284: 12663–12669, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Lee J, Kim MS, Li R, Liu VY, Fu L, Moore DD, Ma K, Yechoor VK. Loss of Bmal1 leads to uncoupling and impaired glucose-stimulated insulin secretion in beta-cells. Islets 3: 381–388, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Lefta M, Campbell KS, Feng HZ, Jin JP, Esser KA. Development of dilated cardiomyopathy in Bmal1 deficient mice. Am J Physiol Heart Circ Physiol 303: H475–H485, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Li AJ, Wiater MF, Oostrom MT, Smith BR, Wang Q, Dinh TT, Roberts BL, Jansen HT, Ritter S. Leptin-sensitive neurons in the arcuate nuclei contribute to endogenous feeding rhythms. Am J Physiol Regul Integr Comp Physiol 302: R1313–R1326, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Lieu SJ, Curhan GC, Schernhammer ES, Forman JP. Rotating night shift work and disparate hypertension risk in African-Americans. J Hypertens 30: 61–66, 2012 [DOI] [PubMed] [Google Scholar]
- 76.Liu C, Weaver DR, Jin X, Shearman LP, Pieschl RL, Gribkoff VK, Reppert SM. Molecular dissection of two distinct actions of melatonin on the suprachiasmatic circadian clock. Neuron 19: 91–102, 1997 [DOI] [PubMed] [Google Scholar]
- 77.Liu C, Wu Y, Li H, Qi Q, Langenberg C, Loos RJ, Lin X. MTNR1B rs10830963 is associated with fasting plasma glucose, HbA1C and impaired beta-cell function in Chinese Hans from Shanghai. BMC Med Genet 11: 59, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Logan RW, Zhang C, Murugan S, O'Connell S, Levitt D, Rosenwasser AM, Sarkar DK. Chronic shift-lag alters the circadian clock of NK cells and promotes lung cancer growth in rats. J Immunol 188: 2583–2591, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Loudon AS. Circadian biology: a 2.5 billion year old clock. Curr Biol 22: R570–R571, 2012 [DOI] [PubMed] [Google Scholar]
- 80.Lubarski I, Pihakaski-Maunsbach K, Karlish SJ, Maunsbach AB, Garty H. Interaction with the Na,K-ATPase and tissue distribution of FXYD5 (related to ion channel). J Biol Chem 280: 37717–37724, 2005 [DOI] [PubMed] [Google Scholar]
- 81.Mahoney MM, Sisk C, Ross HE, Smale L. Circadian regulation of gonadotropin-releasing hormone neurons and the preovulatory surge in luteinizing hormone in the diurnal rodent, Arvicanthis niloticus, and in a nocturnal rodent, Rattus norvegicus. Biol Reprod 70: 1049–1054, 2004 [DOI] [PubMed] [Google Scholar]
- 82.Mantele S, Otway DT, Middleton B, Bretschneider S, Wright J, Robertson MD, Skene DJ, Johnston JD. Daily rhythms of plasma melatonin, but not plasma leptin or leptin mRNA, vary between lean, obese and type 2 diabetic men. PLos One 7: e37123, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Marcheva B, Ramsey KM, Buhr ED, Kobayashi Y, Su H, Ko CH, Ivanova G, Omura C, Mo S, Vitaterna MH, Lopez JP, Philipson LH, Bradfield CA, Crosby SD, JeBailey L, Wang X, Takahashi JS, Bass J. Disruption of the clock components CLOCK and BMAL1 leads to hypoinsulinaemia and diabetes. Nature 466: 627–631, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Marques FZ, Campain AE, Tomaszewski M, Zukowska-Szczechowska E, Yang YH, Charchar FJ, Morris BJ. Gene expression profiling reveals renin mRNA overexpression in human hypertensive kidneys and a role for microRNAs. Hypertension 58: 1093–1098, 2011 [DOI] [PubMed] [Google Scholar]
- 85.Marrone O, Salvaggio A, Bue AL, Bonanno A, Riccobono L, Insalaco G, Bonsignore MR. Blood pressure changes after automatic and fixed CPAP in obstructive sleep apnea: relationship with nocturnal sympathetic activity. Clin Exp Hypertens 33: 373–380, 2011 [DOI] [PubMed] [Google Scholar]
- 86.Martino TA, Tata N, Belsham DD, Chalmers J, Straume M, Lee P, Pribiag H, Khaper N, Liu PP, Dawood F, Backx PH, Ralph MR, Sole MJ. Disturbed diurnal rhythm alters gene expression and exacerbates cardiovascular disease with rescue by resynchronization. Hypertension 49: 1104–1113, 2007 [DOI] [PubMed] [Google Scholar]
- 87.Martino TA, Tata N, Simpson JA, Vanderlaan R, Dawood F, Kabir MG, Khaper N, Cifelli C, Podobed P, Liu PP, Husain M, Heximer S, Backx PH, Sole MJ. The primary benefits of angiotensin-converting enzyme inhibition on cardiac remodeling occur during sleep time in murine pressure overload hypertrophy. J Am Coll Cardiol 57: 2020–2028, 2011 [DOI] [PubMed] [Google Scholar]
- 88.Mazzoccoli G, Southern RB, De Cata A, Giuliani F, Fontana A, Copetti M, Pellegrini F, Tarquini R. A timetable of 24-hour patterns for human lymphocyte subpopulations. J Biol Regul Homeost Agents 25: 387–395, 2011 [PubMed] [Google Scholar]
- 89.Mojon A, Ayala DE, Pineiro L, Otero A, Crespo JJ, Moya A, Boveda J, de Lis JP, Fernandez JR, Hermida RC. Comparison of ambulatory blood pressure parameters of hypertensive patients with and without chronic kidney disease. Chronobiol Int 30: 145–158, 2012 [DOI] [PubMed] [Google Scholar]
- 90.Moniwa N, Varagic J, Ahmad S, VonCannon JL, Ferrario CM. Restoration of the blood pressure circadian rhythm by direct renin inhibition and blockade of angiotensin II receptors in mRen2.Lewis hypertensive rats. Ther Adv Cardiovasc Dis 6: 15–29, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Morse D, Cermakian N, Brancorsini S, Parvinen M, Sassone-Corsi P. No circadian rhythms in testis: Period1 expression is clock independent and developmentally regulated in the mouse. Mol Endocrinol 17: 141–151, 2003 [DOI] [PubMed] [Google Scholar]
- 92.Naito Y, Tsujino T, Kawasaki D, Okumura T, Morimoto S, Masai M, Sakoda T, Fujioka Y, Ohyanagi M, Iwasaki T. Circadian gene expression of clock genes and plasminogen activator inhibitor-1 in heart and aorta of spontaneously hypertensive and Wistar-Kyoto rats. J Hypertens 21: 1107–1115, 2003 [DOI] [PubMed] [Google Scholar]
- 93.Narasimamurthy R, Hatori M, Nayak SK, Liu F, Panda S, Verma IM. Circadian clock protein cryptochrome regulates the expression of proinflammatory cytokines. Proc Natl Acad Sci USA 109: 12662–12667, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Nebzydoski SJ, Pozzo S, Nemec L, Rankin MK, Gressley TF. The effect of dexamethasone on clock gene mRNA levels in bovine neutrophils and lymphocytes. Vet Immunol Immunopathol 138: 183–192, 2010 [DOI] [PubMed] [Google Scholar]
- 95.Nikolaeva S, Pradervand S, Centeno G, Zavadova V, Tokonami N, Maillard M, Bonny O, Firsov D. The circadian clock modulates renal sodium handling. J Am Soc Nephrol 23: 1019–1026, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Nishinaga H, Komatsu R, Doi M, Fustin JM, Yamada H, Okura R, Yamaguchi Y, Matsuo M, Emoto N, Okamura H. Circadian expression of the Na+/H+ exchanger NHE3 in the mouse renal medulla. Biomed Res (Tokyo) 30: 87–93, 2009 [DOI] [PubMed] [Google Scholar]
- 97.Ohta H, Mitchell AC, McMahon DG. Constant light disrupts the developing mouse biological clock. Pediatr Res 60: 304–308, 2006 [DOI] [PubMed] [Google Scholar]
- 98.Olcese J, Domagalski R, Bednorz A, Weaver DR, Urbanski HF, Reuss S, Middendorff R. Expression and regulation of mPer1 in immortalized GnRH neurons. Neuroreport 14: 613–618, 2003 [DOI] [PubMed] [Google Scholar]
- 99.Olcese J, Sikes HE, Resuehr D. Induction of PER1 mRNA expression in immortalized gonadotropes by gonadotropin-releasing hormone (GnRH): involvement of protein kinase C and MAP kinase signaling. Chronobiol Int 23: 143–150, 2006 [DOI] [PubMed] [Google Scholar]
- 100.Padmanabhan K, Robles MS, Westerling T, Weitz CJ. Feedback regulation of transcriptional termination by the mammalian circadian clock PERIOD complex. Science 337: 599–602, 2012 [DOI] [PubMed] [Google Scholar]
- 101.Panza JA, Epstein SE, Quyyumi AA. Circadian variation in vascular tone and its relation to alpha-sympathetic vasoconstrictor activity. N Engl J Med 325: 986–990, 1991 [DOI] [PubMed] [Google Scholar]
- 102.Pappa KI, Gazouli M, Anastasiou E, Iliodromiti Z, Antsaklis A, Anagnou NP. The major circadian pacemaker ARNT-like protein-1 (BMAL1) is associated with susceptibility to gestational diabetes mellitus. Diabetes Res Clin Pract 99: 151–157, 2012 [DOI] [PubMed] [Google Scholar]
- 103.Peruquetti RL, de Mateo S, Sassone-Corsi P. Circadian proteins CLOCK and BMAL1 in the chromatoid body, a RNA processing granule of male germ cells. PLos One 7: e42695, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Peschke E, Muhlbauer E. New evidence for a role of melatonin in glucose regulation. Best Pract Res Clin Endocrinol Metab 24: 829–841, 2010 [DOI] [PubMed] [Google Scholar]
- 105.Pezuk P, Mohawk JA, Wang LA, Menaker M. Glucocorticoids as entraining signals for peripheral circadian oscillators. Endocrinology 153: 4775–4783, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Pfeffer M, Rauch A, Korf HW, von Gall C. The endogenous melatonin (MT) signal facilitates reentrainment of the circadian system to light-induced phase advances by acting upon MT2 receptors. Chronobiol Int 29: 415–429, 2012 [DOI] [PubMed] [Google Scholar]
- 107.Polonia J, Diogo D, Caupers P, Damasceno A. Influence of two doses of irbesartan on non-dipper circadian blood pressure rhythm in salt-sensitive black hypertensives under high salt diet. J Cardiovasc Pharmacol 42: 98–104, 2003 [DOI] [PubMed] [Google Scholar]
- 108.Prasai MJ, Pernicova I, Grant PJ, Scott EM. An endocrinologist's guide to the clock. J Clin Endocrinol Metab 96: 913–922, 2011 [DOI] [PubMed] [Google Scholar]
- 109.Radogna F, Diederich M, Ghibelli L. Melatonin: a pleiotropic molecule regulating inflammation. Biochem Pharmacol 80: 1844–1852, 2010 [DOI] [PubMed] [Google Scholar]
- 110.Ramsey KM, Yoshino J, Brace CS, Abrassart D, Kobayashi Y, Marcheva B, Hong HK, Chong JL, Buhr ED, Lee C, Takahashi JS, Imai S, Bass J. Circadian clock feedback cycle through NAMPT-mediated NAD+ biosynthesis. Science 324: 651–654, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Ratajczak CK, Boehle KL, Muglia LJ. Impaired steroidogenesis and implantation failure in Bmal1-/- mice. Endocrinology 150: 1879–1885, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Reddy AB, Maywood ES, Karp NA, King VM, Inoue Y, Gonzalez FJ, Lilley KS, Kyriacou CP, Hastings MH. Glucocorticoid signaling synchronizes the liver circadian transcriptome. Hepatology 45: 1478–1488, 2007 [DOI] [PubMed] [Google Scholar]
- 113.Reddy TE, Gertz J, Crawford GE, Garabedian MJ, Myers RM. The hypersensitive glucocorticoid response specifically regulates period 1 and expression of circadian genes. Mol Cell Biol 32: 3756–3767, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Redman J, Armstrong S, Ng KT. Free-running activity rhythms in the rat: entrainment by melatonin. Science 219: 1089–1091, 1983 [DOI] [PubMed] [Google Scholar]
- 115.Reiter R, Swingen C, Moore L, Henry TD, Traverse JH. Circadian Dependence of infarct size and left ventricular function after ST elevation myocardial infarction. Circ Res 110: 105–110, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Richards J, Greenlee MM, Jeffers LA, Cheng KY, Guo L, Eaton DC, Gumz ML. Inhibition of alphaENaC expression and ENaC activity following blockade of the circadian clock-regulatory kinases CKIδ/ε. Am J Physiol Renal Physiol 303: F918–F927, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Richards J, Gumz ML. Advances in understanding the peripheral circadian clocks. FASEB J 26: 3602–3613, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Rutter J, Reick M, Wu LC, McKnight SL. Regulation of clock and NPAS2 DNA binding by the redox state of NAD cofactors. Science 293: 510–514, 2001 [DOI] [PubMed] [Google Scholar]
- 119.Sachdeva A, Weder AB. Nocturnal sodium excretion, blood pressure dipping, and sodium sensitivity. Hypertension 48: 527–533, 2006 [DOI] [PubMed] [Google Scholar]
- 120.Saifur Rohman M, Emoto N, Nonaka H, Okura R, Nishimura M, Yagita K, van der Horst GT, Matsuo M, Okamura H, Yokoyama M. Circadian clock genes directly regulate expression of the Na(+)/H(+) exchanger NHE3 in the kidney. Kidney Int 67: 1410–1419, 2005 [DOI] [PubMed] [Google Scholar]
- 121.Schoenhard JA, Smith LH, Painter CA, Eren M, Johnson CH, Vaughan DE. Regulation of the PAI-1 promoter by circadian clock components: differential activation by BMAL1 and BMAL2. J Mol Cell Cardiol 35: 473–481, 2003 [DOI] [PubMed] [Google Scholar]
- 122.Sellix MT, Menaker M. Circadian clocks in the ovary. Trends Endocrinol Metab 21: 628–636, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Shea SA, Hilton MF, Hu K, Scheer FA. Existence of an endogenous circadian blood pressure rhythm in humans that peaks in the evening. Circ Res 108: 980–984, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Sherman H, Genzer Y, Cohen R, Chapnik N, Madar Z, Froy O. Timed high-fat diet resets circadian metabolism and prevents obesity. FASEB J 26: 3493–3502, 2012 [DOI] [PubMed] [Google Scholar]
- 125.Silver AC, Arjona A, Walker WE, Fikrig E. The circadian clock controls toll-like receptor 9-mediated innate and adaptive immunity. Immunity 36: 251–261, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Smarr BL, Morris E, de la Iglesia HO. The dorsomedial suprachiasmatic nucleus times circadian expression of Kiss1 and the luteinizing hormone surge. Endocrinology 153: 2839–2850, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Solt LA, Wang Y, Banerjee S, Hughes T, Kojetin DJ, Lundasen T, Shin Y, Liu J, Cameron MD, Noel R, Yoo SH, Takahashi JS, Butler AA, Kamenecka TM, Burris TP. Regulation of circadian behaviour and metabolism by synthetic REV-ERB agonists. Nature 485: 62–68, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Son GH, Chung S, Kim K. The adrenal peripheral clock: glucocorticoid and the circadian timing system. Front Neuroendocrinol 32: 451–465, 2011 [DOI] [PubMed] [Google Scholar]
- 129.Spengler ML, Kuropatwinski KK, Comas M, Gasparian AV, Fedtsova N, Gleiberman AS, Gitlin II, Artemicheva NM, Deluca KA, Gudkov AV, Antoch MP. Core circadian protein CLOCK is a positive regulator of NF-kappaB-mediated transcription. Proc Natl Acad Sci USA 109: E2457–E2465, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Staessen J, Broughton PM, Fletcher AE, Markowe HL, Marmot MG, Rose G, Semmence A, Shipley MJ, Bulpitt CJ. The assessment of the relationship between blood pressure and sodium intake using whole-day, daytime and overnight urine collections. J Hypertens 9: 1035–1040, 1991 [DOI] [PubMed] [Google Scholar]
- 131.Staessen JA, Birkenhager W, Bulpitt CJ, Fagard R, Fletcher AE, Lijnen P, Thijs L, Amery A. The relationship between blood pressure and sodium and potassium excretion during the day and at night. J Hypertens 11: 443–447, 1993 [DOI] [PubMed] [Google Scholar]
- 132.Stamenkovic JA, Olsson AH, Nagorny CL, Malmgren S, Dekker-Nitert M, Ling C, Mulder H. Regulation of core clock genes in human islets. Metabolism 61: 978–985, 2012 [DOI] [PubMed] [Google Scholar]
- 133.Storch KF, Lipan O, Leykin I, Viswanathan N, Davis FC, Wong WH, Weitz CJ. Extensive and divergent circadian gene expression in liver and heart. Nature 417: 78–83, 2002 [DOI] [PubMed] [Google Scholar]
- 134.Stow LR, Gumz ML. The circadian clock in the kidney. J Am Soc Nephrol 22: 598–604, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Stow LR, Richards J, Cheng KY, Lynch IJ, Jeffers LA, Greenlee MM, Cain BD, Wingo CS, Gumz ML. The circadian protein period 1 contributes to blood pressure control and coordinately regulates renal sodium transport genes. Hypertension 59: 1151–1156, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Stow LR, Voren GE, Gumz ML, Wingo CS, Cain BD. Dexamethasone stimulates endothelin-1 gene expression in renal collecting duct cells. Steroids 77: 360–366, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Sujino M, Furukawa K, Koinuma S, Fujioka A, Nagano M, Iigo M, Shigeyoshi Y. Differential entrainment of peripheral clocks in the rat by glucocorticoid and feeding. Endocrinology 153: 2277–2286, 2012 [DOI] [PubMed] [Google Scholar]
- 138.Suzuki J, Ogawa M, Tamura N, Maejima Y, Takayama K, Maemura K, Honda K, Hirata Y, Nagai R, Isobe M. A critical role of sympathetic nerve regulation for the treatment of impaired daily rhythm in hypertensive Dahl rats. Hypertens Res 33: 1060–1065, 2010 [DOI] [PubMed] [Google Scholar]
- 139.Takeda N, Maemura K. Circadian clock and cardiovascular disease. J Cardiol 57: 249–256, 2011 [DOI] [PubMed] [Google Scholar]
- 140.Tanida M, Yamatodani A, Niijima A, Shen J, Todo T, Nagai K. Autonomic and cardiovascular responses to scent stimulation are altered in cry KO mice. Neurosci Lett 413: 177–182, 2007 [DOI] [PubMed] [Google Scholar]
- 141.Tian XY, Wong WT, Xu A, Lu Y, Zhang Y, Wang L, Cheang WS, Wang Y, Yao X, Huang Y. Uncoupling protein-2 protects endothelial function in diet-induced obese mice. Circ Res 110: 1211–1216, 2012 [DOI] [PubMed] [Google Scholar]
- 142.Tsimakouridze EV, Straume M, Podobed PS, Chin H, LaMarre J, Johnson R, Antenos M, Kirby GM, Mackay A, Huether P, Simpson JA, Sole M, Gadal G, Martino TA. Chronomics of pressure overload-induced cardiac hypertrophy in mice reveals altered day/night gene expression and biomarkers of heart disease. Chronobiol Int 29: 810–821, 2012 [DOI] [PubMed] [Google Scholar]
- 143.Turek FW, Joshu C, Kohsaka A, Lin E, Ivanova G, McDearmon E, Laposky A, Losee-Olson S, Easton A, Jensen DR, Eckel RH, Takahashi JS, Bass J. Obesity and metabolic syndrome in circadian Clock mutant mice. Science 308: 1043–1045, 2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Uzu T, Kimura G. Diuretics shift circadian rhythm of blood pressure from nondipper to dipper in essential hypertension. Circulation 100: 1635–1638, 1999 [DOI] [PubMed] [Google Scholar]
- 145.Uzu T, Kimura G, Yamauchi A, Kanasaki M, Isshiki K, Araki S, Sugiomoto T, Nishio Y, Maegawa H, Koya D, Haneda M, Kashiwagi A. Enhanced sodium sensitivity and disturbed circadian rhythm of blood pressure in essential hypertension. J Hypertens 24: 1627–1632, 2006 [DOI] [PubMed] [Google Scholar]
- 146.van der Veen DR, Shao J, Xi Y, Li L, Duffield GE. Cardiac atrial circadian rhythms in PERIOD2::LUCIFERASE and per1:luc Mice: amplitude and phase responses to glucocorticoid signaling and medium treatment. PLos One 7: e47692, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Venegas C, Garcia JA, Doerrier C, Volt H, Escames G, Lopez LC, Reiter RJ, Acuna-Castroviejo D. Analysis of the daily changes of melatonin receptors in the rat liver. J Pineal Res 54: 313–321, 2012 [DOI] [PubMed] [Google Scholar]
- 148.Vieira E, Marroqui L, Batista TM, Caballero-Garrido E, Carneiro EM, Boschero AC, Nadal A, Quesada I. The clock gene Rev-erbalpha regulates pancreatic beta-cell function: modulation by leptin and high-fat diet. Endocrinology 153: 592–601, 2012 [DOI] [PubMed] [Google Scholar]
- 149.Virshup DM, Eide EJ, Forger DB, Gallego M, Harnish EV. Reversible protein phosphorylation regulates circadian rhythms. Cold Spring Harb Symp Quant Biol 72: 413–420, 2007 [DOI] [PubMed] [Google Scholar]
- 150.Viswambharan H, Carvas JM, Antic V, Marecic A, Jud C, Zaugg CE, Ming XF, Montani JP, Albrecht U, Yang Z. Mutation of the circadian clock gene Per2 alters vascular endothelial function. Circulation 115: 2188–2195, 2007 [DOI] [PubMed] [Google Scholar]
- 151.Waddell BJ, Wharfe MD, Crew RC, Mark PJ. A rhythmic placenta? Circadian variation, clock genes and placental function. Placenta 33: 533–539, 2012 [DOI] [PubMed] [Google Scholar]
- 152.Wang N, Yang G, Jia Z, Zhang H, Aoyagi T, Soodvilai S, Symons JD, Schnermann JB, Gonzalez FJ, Litwin SE, Yang T. Vascular PPARgamma controls circadian variation in blood pressure and heart rate through Bmal1. Cell Metab 8: 482–491, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Westgate EJ, Cheng Y, Reilly DF, Price TS, Walisser JA, Bradfield CA, FitzGerald GA. Genetic components of the circadian clock regulate thrombogenesis in vivo. Circulation 117: 2087–2095, 2008 [DOI] [PubMed] [Google Scholar]
- 154.Wu T, Ni Y, Dong Y, Xu J, Song X, Kato H, Fu Z. Regulation of circadian gene expression in the kidney by light and food cues in rats. Am J Physiol Regul Integr Comp Physiol 298: R635–R641, 2010 [DOI] [PubMed] [Google Scholar]
- 155.Zhang EE, Kay SA. Clocks not winding down: unravelling circadian networks. Nat Rev Mol Cell Biol 11: 764–776, 2010 [DOI] [PubMed] [Google Scholar]
- 156.Zhang EE, Liu Y, Dentin R, Pongsawakul PY, Liu AC, Hirota T, Nusinow DA, Sun X, Landais S, Kodama Y, Brenner DA, Montminy M, Kay SA. Cryptochrome mediates circadian regulation of cAMP signaling and hepatic gluconeogenesis. Nat Med 16: 1152–1156, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Zuber AM, Centeno G, Pradervand S, Nikolaeva S, Maquelin L, Cardinaux L, Bonny O, Firsov D. Molecular clock is involved in predictive circadian adjustment of renal function. Proc Natl Acad Sci USA 106: 16523–16528, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]