
Figure 4.
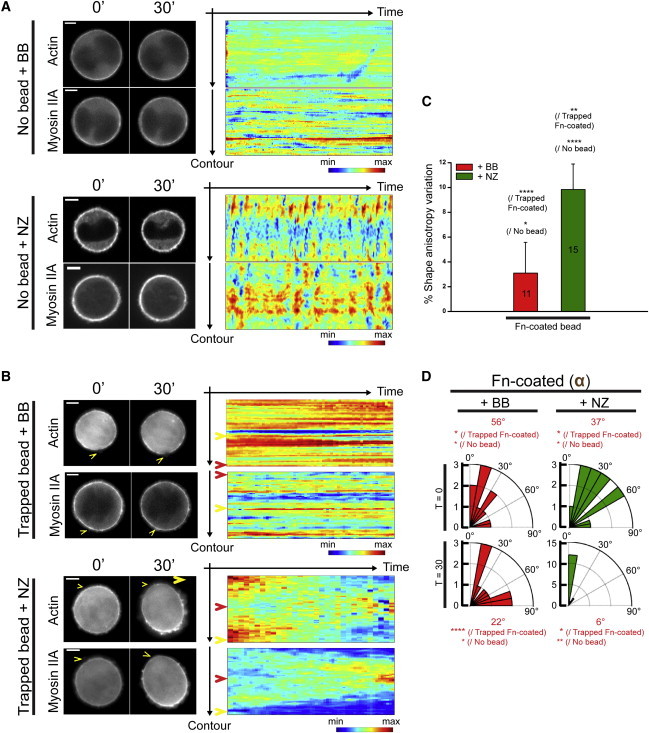
The dynamics of cortical actomyosin drive myosin-II-dependent cell-shape behaviors. Fluorescence images of single nonadherent cells expressing eGFP-actin and eGFP-myosin-IIA treated with chemical perturbants. (A) Cell-shape oscillation. Cell-contour kymographs exhibit fluctuation of cortical actomyosin when cells are treated with nocodazole (NZ), whereas the cortical distribution of actomyosin is frozen in the presence of blebbistatin (BB). (B–D) The application of a single Fn-coated bead does not induce cell shape polarization in the presence of BB. (B) The kymograph highlights a frozen cortical distribution of actomyosin. Depolymerizing the MTs does not impair the polarized redistribution of cortical actomyosin. (Yellow arrowheads) Cortical angular position of the trigger; (red arrowheads) opposite pole. Representative contour kymographs are displayed for each condition. All scale bars, 5 μm. (C) The shape anisotropy variation of nonadherent 3T3 cells is monitored while triggering cell shape polarization in the presence of BB or NZ. (D) Rose diagrams indicate the proportion of cells for each angular sector upon addition of BB or NZ. (Red) Median angle. Statistical differences are indicated (Dunnett’s test: P < 1; ∗∗, P < 0.5; ∗∗∗, P < 0.02; ∗∗∗∗, P < 0.0001).