
Extended Data Figure 6. miR-34/449 deficiency in frog MCCs causes defective ciliogenesis without affecting cell fate specification.
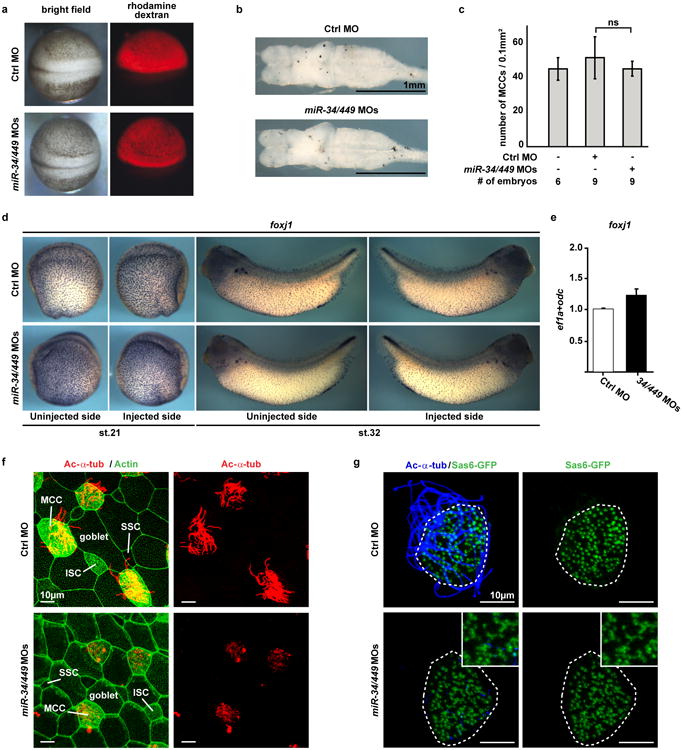
a. Injection of Ctrl or miR-34/449 MOs does not affect general embryonic development or neural tube closure. Xenopus laevis embryos were injected unilaterally with MOs at the 2-4 cell stage and analyzed at neurula stages (18-20). Targeting of the skin ectoderm was confirmed by coinjection of fluorescent rhodamine dextran. b. Frog miR-34/449 morphants do not exhibit hydrocephalus. Embryos were injected animally with control or miR-34/449 MOs into both dorsal blastomeres at the 4 cell stage to target the neural tube and brain regions. Subsequently, the whole brains were dissected and analyzed at stage 45/46. The lack of hydrocephalus in miR-34/449 morphants argues against a role of miR-34/449 in ependymal ciliation. c. Quantification of fully ciliated, partially ciliated or non-ciliated MCCs reveals no significant change in total number of MCC-fated cells in miR-34/449 morphants. Error bars represent s.d. Wilcoxon Two Sample test, n.s., P > 0.05. d, e. foxj1 expression and specification of MCC fate is unaltered in miR-34/449 deficient embryos. d. Embryos were unilaterally injected with Ctrl or miR-34/449 MOs to the right side at 2-4 cell stage, cultured until stage 21 or 32 and processed for in situ hybridization to monitor foxj1 expression in the mucociliary epithelium of the skin. No change in foxj1 expression can be detected. e. Real time PCR analysis in Ctrl or miR34/449 MOs injected skin explants at stage 26 (onset of ciliation) does not indicate reduced expression levels of foxj1. f. miR-34/449 deficient frog embryos exhibit normal development of the mucociliary ectodermal epithelium. Detailed analysis of the embryonic skin at stage 30-32 reveals the presence (specification and intercalation) of all cell types in miR-34/449 morphants, including large goblet cells, small secretory cells (SSC), Ac-α-tub positive ciliated cells (MCC) and non-tubulin enriched ion secreting cells (ISC). g. miR-34/449 morphant MCCs exhibit an uneven distribution of basal bodies. Sas6-gfp mRNA was injected at the 2-4 cell stage to visualize basal bodies at stage 30-32. In control embryos Sas6-GFP foci are evenly distributed in fully ciliated MCCs, while miR-34/449 morphant MCCs are characterized by an uneven distribution and aggregation of basal bodies, which frequently fail to grow cilia (Ac-α-tub staining). Such phenotype is characteristic for basal body docking defects. Embryos/cells analyzed: Ctrl MO (4/7), miR-34/440MOs (6/10).