
Figure 5. A model for regulation of HRR by Fbh1.
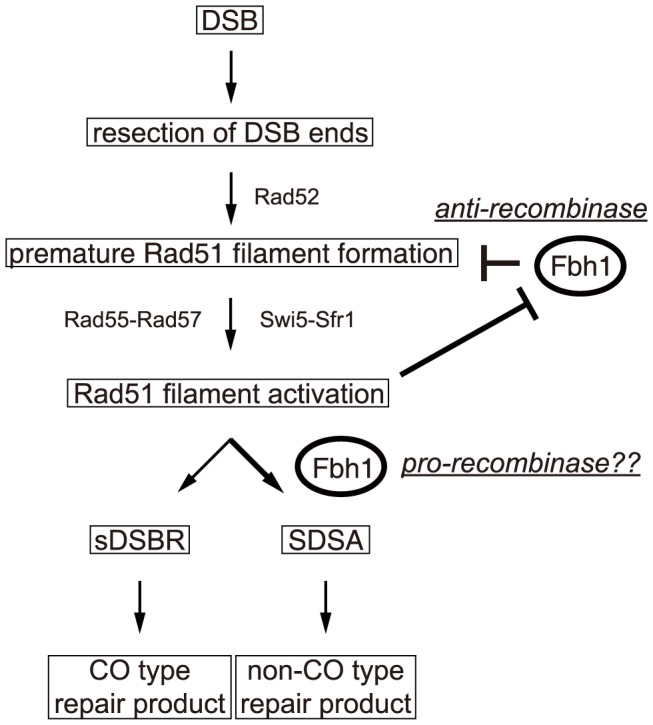
After DSB formation, the ends of a DSB are nucleolytically processed to produce recombinogenic DNA with 3′ ss overhangs. Rad51 binds the ssDNA region, an interaction that is mediated by Rad52. Because the resulting Rad51 filament is not yet activated (‘premature’), it is prone to be disrupted by the ‘anti-recombinase’ activity of Fbh1. Swi5-Sfr1 protects the Rad51 filament by activating it. It remains unclear whether Rad55-Rad57 also protects the Rad51 filament. Previously, we proposed that both Rad55-Rad57 and Swi5-Sfr1 sub-pathways are involved in DSBR and synthesis-dependent strand annealing (SDSA), and that Rad55-Rad57 is required for second-end capture (SEC) to form the double-Holliday junction in DSBR [8]. In this study, the results of the single-DSBR assay suggest that Fbh1 suppresses crossover formation, most probably by inhibiting Rad55-Rad57-mediated SEC during DSBR. On the other hand, the in vitro data showing Fbh1 stimulation of branch migration in the strand-exchange reaction suggests that Fbh1 might have a pro-recombination role in vivo, probably by stimulating the SDSA pathway to generate a non-crossover repair product. This possibility needs to be addressed in future studies. Subsequently, Rad51 would be degraded by SCFFbh1-dependent ubiquitination to prevent re-activation, either after the reaction or during stationary phase.