

Abstract
In Arabidopsis, the regulation network of the seed maturation program controls the induction of seed dormancy. Wheat EST sequences showing homology with the master regulators of seed maturation, LEAFY COTYLEDON1 (LEC1), LEC2 and FUSCA3 (FUS3), were searched from databases and designated respectively as TaL1L (LEC1-LIKE), TaL2L (LEC2-LIKE), and TaFUS3. TaL1LA, TaL2LA and TaFUS3 mainly expressed in seeds or embryos, with the expression limited to the early stages of seed development. Results show that tissue-specific and developmental-stage-dependent expressions are similar to those of seed maturation regulators in Arabidopsis. In wheat cultivars, the expression level of TaL1LA is correlated significantly with the germination index (GI) of whole seeds at 40 days after pollination (DAP) (r = –0.83**). Expression levels of TaFUS3 and TaL2LA are significantly correlated respectively with GIs at 40 DAP and 50 DAP, except for dormant cultivars. No correlation was found between the expression level of TaVP1, orthologue of ABA INSENSITIVE3 (ABI3), and seed dormancy. DELAY OF GERMINATION1 (DOG1) was identified as a quantitative trait locus (QTL) for the regulation of seed dormancy in Arabidopsis. Its promoter has RY motif, which is a target sequence of LEC2. Significant correlation was found between the expression of TaDOG1 and seed dormancy except for dormant cultivars. These results indicate that TaL1LA, TaL2LA, and TaFUS3 are wheat orthologues of seed maturation regulators. The expressions of these genes affect the level of seed dormancy. Furthermore, the pathways, which involve seed maturation regulators and TaDOG1, are important for regulating seed dormancy in wheat.
Introduction
Pre-harvest sprouting, the premature germination of seeds in the spike before harvest, is influenced by rainfall and temperature during seed development. Pre-harvest sprouting reduces yields and grain quality by increasing the activity of hydrolytic enzymes such as α-amylase, and produces a large economic loss [1]. Seed dormancy, which is an important factor in the control of pre-harvest sprouting, prevents germination under favorable conditions (e.g. moisture and temperature) [2]. It is an inherent capacity for adaptation to several environments. However, a high level of dormancy is economically undesirable because uniform and rapid germination are necessary for crop cultivation [3]. Therefore, modern cultivars have lower seed dormancy, which triggers pre-harvest sprouting. Sprouting damage is becoming a severe problem to the production of cereals worldwide. For these reasons, resistance to pre-harvest sprouting is a critical breeding objective in the cultivation of many cereals including wheat and barley.
A plant hormone, abscisic acid (ABA), plays a fundamental role in the induction and maintenance of dormancy. In Arabidopsis, many genes involved in the biosynthesis (ABA1, ABA2, NCEDs), catabolism (CYP707A), and signaling (ABIs, ERA, ABH) have been identified [4]–[10]. Recent molecular genetic approaches suggest that the pathway independent of plant hormones as well as plant hormone dependent pathways are important for the control of seed dormancy [11]. Dormancy, which is induced early during seed maturation, increases until the seed is fully developed [12]. Seed dormancy is affected by the seed maturation program, which is controlled by at least four major regulators in Arabidopsis [13], [14], [15]. LEAFY COTYLEDON2 (LEC2), FUSCA3 (FUS3) and ABA INSENSITIVE3 (ABI3) are plant-specific transcription factors with B3 domain [16]–[18]. LEC1 encodes the HAP3 subunit of the CCAAT-binding transcription factor [19]. Mutations of seed maturation regulators have pleiotropic effects, and the mutants show several morphological and physiological defects. Actually, lec1, lec2 and fus3 mutants represent defects of cotyledon identity, loss of desiccation tolerance, and reduction of seed dormancy [18]–[27]. Furthermore, abi3 mutants show the reduction of desiccation tolerance and seed dormancy. In addition, ABA insensitivity and defects of chlorophyll degradation with normal cotyledon morphology were observed in abi3 mutants.
Transcription factors with B3 domain reveal distinct temporal expression patterns during seed development. Expression of ABI3 is maintained at a higher level through seed maturation, but expression of FUS3 is reduced before maturation [13], [17], [28]. LEC2 shows the peak of expression before the highest expression of FUS3 and ABI3. Furthermore, ABI3 and FUS3 preferentially express, respectively, in the whole embryo and protoderm of embryo [15], [28], [29]. LEC2 is expressed mainly during early embryo development [13], [18]. Therefore, B3 domain family genes have different traits for temporal expression.
LEC2 and FUS3 affect the accumulation of several hormones by regulating distinct pathways. FUS3 negatively regulates the accumulation of active form gibberellin (GA) by suppressing GA3ox2, and it positively regulates the accumulation of ABA [26], [30], [31]. LEC2 also suppresses the expression of GA3ox2 and acts as a positive regulator of auxin biosynthesis [30], [32]. LEC1 regulates the expressions of ABI3 and FUS3, and its peak expression is observed before up-regulation of B3 domain factors [13], [33]. These expression patterns suggest that LEC1 acts on the upstream of B3 regulation network. However, overexpression of LEC2 ectopically activates the embryonic pathways that are regulated by LEC1 [32]. Intricate networks are constructed by seed maturation regulators.
The B3 domain recognizes consensus RY/Sph sequence implicated in the seed specific expression. RY motif, which is involved in the promoter of maturation RNAs expressed during the maturation phase of seed development, is regulated directly by LEC2 [34]. DELAY OF GERMINATION1 (DOG1) has been identified as a quantitative trait locus (QTL) controlling the natural variation of seed dormancy in Arabidopsis [35]. DOG1 protein abundance in freshly harvested seeds acts as a timer for seed dormancy release [36]. In fact, DOG1 also has RY motif in its promoter similar to the seed maturation RNAs. The expression of DOG1 is controlled by B3 domain factors [35]. The regulation pathways involve B3 transcription factors; DOG1 plays an important role in connecting the control of seed maturation and dormancy.
Rikiishi et al. [37] demonstrated that the amount of transcripts of a seed specific transcription factor, TaABF (ABRE Binding Factor) 1, is related to seed dormancy levels in wheat seeds. Several seed-specific transcription factors show fluctuating expressions depending on the developmental stage. Such expressions are determined by the maturation program, but seed maturation regulators, except for ABI3/VIVIPAROUS1 (VP1), have not been investigated in wheat. The present study investigated the expressions of master regulators of seed maturation, LEC1, LEC2, FUS3 and ABI3/VP1, in relation to wheat seed dormancy. EST sequences showing homology with Arabidopsis seed maturation regulators were searched from databases. TaL1L (LEC1-LIKE), TaL2L (LEC2-LIKE), and TaFUS3 were expressed mainly in seeds or embryos. Their expression was limited to the early stages of seed development, suggesting that these ESTs are orthologues of seed maturation regulators in wheat. For monocot species, a few studies have examined seed maturation regulators such as ZmLEC1 in maize [38], HvFUS3 in barley [39], and OsLFL1 in rice [40], [41]. LEC2 orthologue has not been identified in monocots. At present, TaL2LA is the first orthologue of LEC2 identified in a monocot species. Expression levels of wheat seed maturation regulators were examined in wheat cultivars showing different levels of seed dormancy. Then the potential effects of these expressions on dormancy were discussed.
Materials and Methods
Plant materials
Wheat cultivars, AUS1408 (AUS), Chihokukomugi (Chi), Chinese Spring (CS), Kitakei-1354 (Kit), Norin61 (N61), OW104 (OW), RL4137 (RL), Tamaizumi (Tam), and Zenkoujikomugi (Zen) were grown in a field under a plastic roof to protect them from rainfall. Seeds were sown in plastic trays in mid-November. Then 20 seedlings of each cultivar were transplanted to the field in mid-December. Spikes were tagged at anthesis. Seeds were harvested every 5 days from 35 days after pollination (DAP) to 50 DAP and then dried in an oven at 150°C for 3 hr to determine the water contents. To ensure the uniformity of developmental stage, seeds were collected only from primary and secondary florets of the center spikelets.
Germination test
Ten whole seeds and ten half seeds that had been cut transversely and contained the embryos were placed on filter paper in a Petri dish containing 6 ml of distilled water. The Petri dishes were incubated in the dark at 20°C. All germination tests had three replications. Germinated seeds were counted daily for 14 days. A weighted germination index (GI) was calculated to assign maximum weight to seeds that had germinated first and to assign less weight to those that germinated subsequently, as described by Rikiishi and Maekawa [42]. The GI values were converted into arcsine-transformed values and were used for statistical analyses.
RNA isolation and real-time quantitative RT-PCR analysis
Total RNA was isolated from 5 seeds at 10 DAP and 20 embryos at 20–50 DAP, and from 0.2 g of leaf and root tissues derived from Norin61 seedlings grown in the dark for 10 days using commercial kit (FastRNA Pro Green; Qbiogene Inc.). The mRNA was purified from 20 µg of total RNA (Poly (A) Purist MAG; Ambion Inc.). First-strand cDNA was prepared using a reagent kit (PrimeScript RT; Takara Bio Inc.). For quantitative RT-PCR, 5 µL of diluted cDNA (1∶25), 0.2 µM of each primer, and 10 µL of SYBER PremixExTaq (Takara Bio Inc.) with a total reaction volume of 20 µL was used in triplicate. Subsequently, PCR was performed (Light Cycler 2.0; Roche Diagnostics Corp.). The condition of the PCR reaction was 95°C for 10 s, followed by 40 cycles of 95°C for 5 s and 60°C for 20 s. Gene-specific primers designed with Primer Express software (Applied Biosystems) are listed in Table S1. The primers of TaVP1 were designed in the consensus regions among sequences derived from A, B and D genomes. The relative amount of each target transcript was determined by generating standard curves using a dilution series of amplified products of the target sequence. The amount of 1010 times dilution was defined as 1.0. Quantifications were performed using three biological replications. All quantifications were normalized to the amplification of TaCDCP (Cell Division Control Protein) [43]. The accession numbers of TaCDCP and TaDOG1 are, respectively, Ta54227 (Unigene Cluster) and RFL_Contig1498 (TriFLBD). All kits were used according to the respective manufacturer’s protocol.
Sequence analysis
Sequences of ESTs were obtained from searches of NCBI/GenBank/Blast/PlantGDB/TriFLDB databases. Accession numbers of LEC1, LEC2, FUS3, ABI3, VP1, and the respective EST sequences are listed in Table S2. Deduced amino acid sequences of the ESTs were compared with that of corresponding Arabidopsis gene using the bl2seq BLAST program (http://blast.ncbi.nlm.gov/) and similarity was evaluated by Query coverage (%) and E-value. Alignments of B and B3 domains in the deduced amino acid sequences were performed using GENETYX ver. 9 software (Genetyx Corp., Tokyo). Based on aligned sequences, an unrooted phylogenetic tree was constructed using Neighbor-Joining (NJ) method with 100 replications.
Results
1. Search for wheat orthologues of seed maturation regulators
EST sequences showing higher homology with seed maturation regulators of Arabidopsis were sought from several databases. In Arabidopsis, LEC1 and its homologous genes form B domain family. The B domain family genes were categorized into LEC1 type and non-LEC1 type. The LEC1-LIKE (L1L) had been identified as homologous gene of LEC1, thus L1L was categorized into LEC1 type; 83.3% of the amino acid sequences in B domain of L1L are identical with that of LEC1. Fourteen EST sequences representing higher homology with the B domain of LEC1 were found from 10 species (Fig. S1) and are designated as L1L. Deduced amino acid sequences showed less homology with LEC1 in the region outside of the B domain. In the sequence of B domain, ZmLEC1, a maize orthologue, revealed 83.3% amino acid sequences identity with LEC1. Amino acid sequences of wheat ESTs, TaL1LA/B/C, were 74.4%, 62.2% and 65.6%, respectively, identical with LEC1 in the B domain sequence. The percentages of homology are lower than that of ZmLEC1. The amino acid sequence of TaL1LA and L1L showed 74.4% identity in the B domain. TaL1LA showed similar level of homology with both LEC1 and L1L. The B domains of TaL1LB and TaL1LC showed higher homology with L1L (71.1% and 70.0%, respectively) than that with LEC1. A phylogenetic tree was constructed for B domain sequences of the ESTs together with LEC1, L1L, and 7 non-LEC1 type genes of Arabidopsis (Fig. 1A). TaL1LA, HvL1L, and ZmLEC1 were classified as LEC1 type, whereas TaL1LB and TaL1LC belonged to the non-LEC1 type.
Figure 1. Unrooted phylogenetic trees based on the sequences of conserved domains.

Trees were constructed from the deduced amino acid sequences of the B domains of LEC1 type, non-LEC1 type and their orthologues (A) and of the B3 domains of Arabidopsis AFL (ABI3/FUS3/LEC2) group genes and their orthologues (B). Phylogenetic trees were generated by Neighbor-Joining method. Scale bars represent amino acid substitutions per site. Abbreviations represent species names as follows: Af, Avena fatua; Bd, Brachypodium distachyon; Bn, Brassica napus; Cs, Cucumis sativus; Gm, Glycine max; Hv, Hordeum vulgare; Os, Oryza sativa; Ps, Pisum sativum; Pv, Panicum virgatum; Sb, Sorghum bicolor; Sl, Solanum lycopersicum; Ta, Triticum aestivum; Vv, Vitis vinifera; Zm, Zea mays.
HvFUS3 had been identified as a barley orthologue of FUS3. The EST with accession number JV941539 was found to have higher homology with HvFUS3. Deduced amino acid sequence of the EST showed 91.5% identity with HvFUS3. Therefore, the EST was designated as TaFUS3. Amino acid sequences of TaFUS3 and HvFUS3 in the B domain showed 61.2% and 63.1% identity with that of FUS3 of Arabidopsis, respectively.
Seven EST sequences that are found to have homology with LEC2 (Fig. S2) were designated as L2L (LEC2-LIKE). The EST with accession number of HM370539 of Brassica napus (BnL2L) showed highest homology with LEC2, and 66.5% and 77.7% of the deduced amino acid sequences were found to be identical with CDS and B3 domain sequences, respectively. In other L2Ls, the sequences outside of the B3 domain showed lower homology with LEC2. The higher homology was detected only in the B3 domain. The B3 domain sequences of L2Ls showed 47.6% (TaL2LA, TaL2LB) to 53.4% (BdL2L) identity with LEC2. In fact, B3 domains of L2Ls showed 44.7–52.4% and 52.4–58.3% identity with those of FUS3 and ABI3, respectively. The B3 domain sequences of L2Ls showed similar homology with those of LEC2, FUS3, and ABI3. Therefore, B3 domain sequences of LEC2, FUS3, ABI3/VP1 and their homologous EST sequences were classified into three groups using phylogenetic analysis (Fig. 1B). Our results showed that TaL2LA and TaL2LB belonged to the same group as LEC2. Sequence analyses results indicated that TaL1LA, TaL2LA/B and TaFUS3 are respective candidates of LEC1, LEC2, and FUS3 orthologues.
2. Expression analysis of seed maturation regulators
Expression levels of candidate genes were determined in several tissues. TaL1LA, TaL2LA and TaVP1 were expressed in seeds (10 DAP) or embryos (20–50 DAP) and showed no expression in leaf or root tissues (Fig. 2). Although TaFUS3 also showed higher expression in seeds and embryos, faint expression of this gene was detected in leaf and root tissues. However, TaL2LB was expressed in seed, leaf, and root tissues. For temporal expression patterns during seed development, TaL1LA and TaL2LA showed expression at 10 DAP. The expressions were decreased rapidly at 20 DAP (Fig. 3). Expression of TaFUS3 started at 10 DAP. The highest expression was observed at 20 DAP. TaFUS3 was expressed at low level at 30 DAP and thereafter. Although TaVP1 was expressed at a lower level at 10 DAP, a higher level in the remaining seed developmental stages (20–50 DAP) was observed. TaL2LB was expressed at all developmental stages (data not shown). The expression of TaL2LB showed different patterns from that of TaL2LA in terms of developmental stage and tissue specific expressions.
Figure 2. Relative expressions of wheat orthologues of seed maturation regulators in different tissues.

Relative amounts of transcripts were determined in leaves, roots, whole seeds at 10 DAP (TaL1LA, TaL2LA) and embryos at 20 DAP (TaL2LB, TaFUS3, TaVP1). Bars represent SE.
Figure 3. Relative expressions of wheat orthologues of seed maturation regulators at different seed developmental stages.

Relative amounts of transcripts were determined in whole seeds at 10 DAP and in embryos at 20–50 DAP. Bars represent SE.
3. Relation between the expression of seed maturation regulators and seed dormancy in wheat cultivars
Water contents of developing seeds were determined in nine wheat cultivars. All cultivars showed less than 20% at 40–45 DAP. Germination potential was evaluated by the germination index (GI) of half seeds, which were released from dormancy. Germination was observed at 20 DAP and the later stages in all cultivars. GIs of half seeds were low (20.2–37.0) at 20–30 DAP, but increased at 40 DAP (71.3–90.2). GIs of whole seeds (40–60 DAP) from the cultivars that exhibited different levels of seed dormancy varied (Fig. 4). Higher GIs were observed in Chi (44.5–93.8), RL (62.4–79.5), and CS (63.1–91.7), indicating that the levels of seed dormancy in these cultivars were low. Whereas, OW (0–2.4), Zen (0–6.9), and Tam (13.6–15.0) showed lower GIs and retained strong dormancy at 40–60 DAP. Although N61 also showed low GIs (0.7 and 12.9) at 40–50 DAP, the GI increased at 60 DAP (87.1). Dormancy was rapidly released at 60 DAP in N61. Seeds of AUS and Kit showed intermediate GIs and moderate dormancy levels. Relations between dormancy and expression levels of seed maturation regulators were examined in cultivars (Fig. 5). Expression of TaL1LA showed significant correlation with GIs at 40 DAP (r = –0.83**). Higher expressions tended to be associated with a higher level of seed dormancy. No significant correlation was observed in the cases of TaL2LA, TaFUS3, or TaVP1. However, the expressions of TaFUS3 were correlated significantly with GIs except for dormant cultivars, N61, OW, and Zen (r = –0.93**). Expression of TaL1LA also showed a significant correlation with GIs at 50 DAP (r = –0.70*). Although no significant correlation was observed between expressions of TaFUS3 and TaVP1 and GIs at 50 DAP, the expression of TaL2LA was correlated significantly with GIs (r = –0.86*), except for dormant cultivars as observed for TaFUS3 (Fig. 6).
Figure 4. Germination index of whole seeds at 40–60 DAP in wheat cultivars.
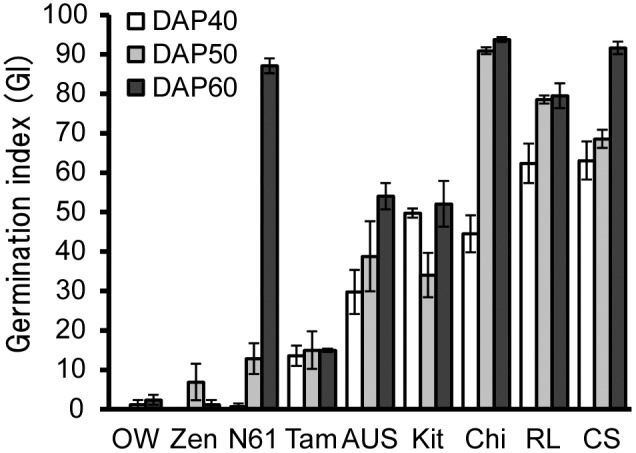
AUS, AUS1408; Chi, Chihokukomugi; CS, Chinese Spring; Kit, Kitakei-1354; N61, Norin61; OW, OW104; RL, RL4137; Tam, Tamaizumi; Zen, Zenkoujikomugi. Bars represent SE.
Figure 5. Relation between dormancy and relative expression of wheat orthologues of seed maturation regulators.

Dormancy represents germination index (GI) of whole seeds at 40 DAP. Numerals in parentheses represent correlation coefficients, except for dormant cultivars, N61, OW, and Zen. Asterisks (**) denote significance at the 1% level.
Figure 6. Relation between dormancy and relative expression of TaL2LA.
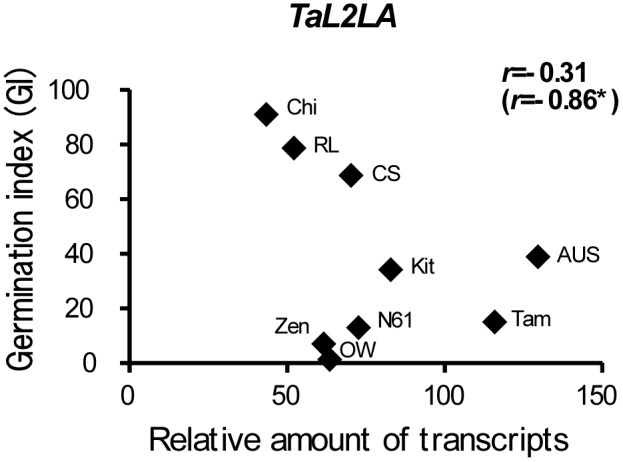
Dormancy represents germination index (GI) of whole seeds at 50 DAP. Numeral in parentheses represent correlation coefficients, except for dormant cultivars, N61, OW, and Zen. Asterisks (*) denote significance at the 5% level.
4. Expression analysis of DOG1 orthologue
TaDOG1 was expressed mainly in the embryos (Rikiishi and Maekawa, 2010). Our results showed peak expression of TaDOG1 at 20 DAP. The expression was decreased during the later developmental stages (Fig. 7A). The highest expression levels of TaDOG1 showed no significant correlation with GIs of whole seeds. However, significant correlation was detected between its expression levels and GIs of whole seeds at 50 DAP in six cultivars except for dormant cultivars, OW, Tam, and Zen (Fig. 7B).
Figure 7. Expression analysis of TaDOG1.
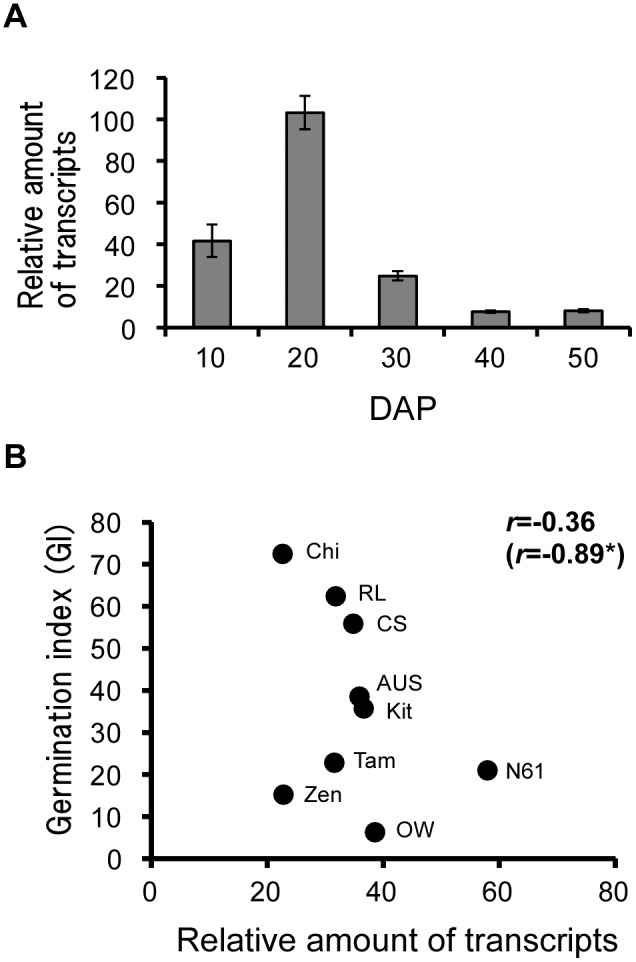
A: Relative expressions of TaDOG1 at different stages of seed development. Bars represent SE. B: Relation between relative expressions of transcripts and germination index (GI) of whole seeds at 50 DAP in wheat cultivars. Numeral in parentheses represent correlation coefficients, except for dormant cultivars, OW, Tam, and Zen. Asterisks (*) denote a significance at the 5% level.
Discussion
Seed maturation is regulated by at least four master regulators in Arabidopsis. These master regulators are classified into two groups based on the conserved domain. LEC1 encodes HAP3 subunit of CCAAT-binding factor with B domain. Other regulators, LEC2, FUS3, and ABI3 belong in the group of transcription factor with B3 domain. These regulators control the seed maturation, dormancy, and desiccation tolerance [12], [13], [15], [22]–[24]. In the present study, wheat orthologues of seed maturation regulators were identified. Relations between their expressions and seed dormancy were examined.
The B domain of LEC1 is conserved in eukaryotes [19]. Genes encoding HAP3 subunit were classified into LEC1 type (LEC1 and L1L) and non-LEC1 type in Arabidopsis [33]. L1L was able to rescue the mutant phenotypes of lec1. However, non-LEC1 type genes did not complement LEC1 functions. L1L has similar functions as LEC1. The functions of non-LEC1 type are distinct from that of LEC1. TaL1LA, TaL1LB, and TaL1LC are homologous EST sequences with LEC1. TaL1LA and TaL1LB/C are classified, respectively, as LEC1 type and non-LEC1 type. Furthermore, TaL1LA is expressed specifically in seeds, in which the expression is limited to 10 DAP during seed development. In Arabidopsis, LEC1 also showed seed-specific expression. Transcripts were detected early during embryogenesis [19], [44]. However, L1L is expressed mainly in the seeds, but weak expression was also observed in the other tissues [33]. Tissue and temporal specific expressions indicated that TaL1LA is a wheat orthologue of LEC1.
HvFUS3 had been identified as barley orthologue of FUS3, and the functions of HvFUS3 are similar to that of FUS3 [39]. TaFUS3 showed higher homology with HvFUS3 and showed seed specific expression. The expression of TaFUS3 was detected at the early stage of seed development. TaVP1 had been identified by McKibbin et al. [45]. Expression of TaVP1 was maintained in higher levels throughout the maturation phase of seed development, similar to that of ABI3 [28]. No orthologue of LEC2 has been identified in monocot species. In the present study, TaL2LA and TaL2LB are found to be wheat ESTs showing homology with the B3 domain of LEC2. Although the B3 domain sequences of TaL2LA and TaL2LB showed similar levels of homology with LEC2, FUS3, and ABI3; TaL2Ls are classified by our phylogenetic analysis into the same group of LEC2. LEC2 showed the highest expression at 8–9 DAP, which showed a decline at the later developmental stages. No transcript of LEC2 was detected in the dry seeds of Arabidopsis [13], [18]. TaL2LA was specifically expressed in the seeds. However, the expression was limited to the early stage of seed development. These results indicate that TaL2LA is a wheat orthologue of LEC2. Kroj et al. [13] reported that the expression of LEC2 precedes that of FUS3 and ABI3 during embryo development. Among our results, TaFUS3 showed the highest expression at 20 DAP, although the peak expression of TaL2LA was observed at 10 DAP. Sequential expressions of TaL2LA and TaFUS3 were similar to those of LEC2 and FUS3, supporting that these genes are orthologues of seed maturation regulators. TaL2LB showed different expression patterns from that of TaL2LA and LEC2. Plant specific transcription factors with B3 domain include several gene families such as AFL (ABI3/FUS3/LEC2), VAL (VP1/ABI3-Like), RAV (Related to ABI3/VP1), ARF (Auxin Response Factor), and REM (Reproductive Meristem) [46]. These transcription factors control several growth and development aspects in Arabidopsis. Diverse functions of B3 domain genes suggest that TaL2LB has other functions than regulating seed dormancy.
Expression levels of seed maturation regulators are compared with seed dormancy levels in wheat cultivars. Expression levels of TaL1LA showed significant correlation with GI of whole seeds. Expression levels of TaL2LA and TaFUS3 were also correlated significantly with seed dormancy levels, except for dormant cultivars. Taking into account these facts and our observations together, the expression levels of TaL1LA, TaL2LA, and TaFUS3 affect the levels of seed dormancy in wheat cultivars. N61, Zen, and OW showed strong dormancy; nevertheless, the expression levels of TaL2LA and TaFUS3 in these cultivars were similar to those of moderate cultivars. Baumbusch et al. [47] reported that the expressions of LEC1, FUS3, and ABI3 do not differ among ecotypes (Ws, C24, Cvi), that show large variation in dormancy levels. The after-ripening requirement of Cvi was distinct from those of other accessions in Arabidopsis [48], suggesting that the level of dormancy might be determined by different regulatory pathways among ecotypes. Some of those pathways might be divergent from those for seed maturation regulators. Dormancy levels of OW and Zen cultivars were found to be distinct from other cultivars because poor germination was observed at 60 DAP. In these cultivars, dormancy levels might be regulated mainly by a pathway that is independent from that of seed maturation regulators.
Jones et al. [49] demonstrated that the mRNA level of AfVP1 has strong correlation with seed dormancy level in wild oat, Avena fatua. However, relative amounts of TaVP1 transcripts were unrelated to seed dormancy in our experiments. Liu et al. [50] also reported that the expression of TaVP1 is not related to dormancy release by after-ripening. McKibbin et al. [45] reported that alternative splicing of TaVP1 may contribute to increased susceptibility to pre-harvest sprouting. In wheat, the abundance of misspliced transcripts might affect the function of TaVP1.
Studies of the regulation networks of seed maturation have indicated that LEC1, which acts as a primary regulator of FUS3 and ABI3, regulates early embryogenesis, desiccation tolerance and accumulation of seed storage proteins. However, LEC2 is not involved in these pathways [51]. LEC2 regulates triacylglycerol and fatty acid biosynthesis, as well as the synthesis of seed storage proteins independent of the regulation pathway of LEC1. Our results show that, although the expression levels of TaL1LA and TaFUS3 are significantly correlated with GIs at 40 DAP, TaL2LA showed correlation with GIs at 50 DAP. The expressions of TaL1LA/TaFUS3 and TaL2LA affect the levels of seed dormancy at different developmental stages. Thus, TaL1LA and TaL2LA might act on the different regulatory pathways similar to those of Arabidopsis. However, the regulatory networks of seed maturation remain unknown in monocot species. In fact, only few studies have examined seed maturation regulators in monocots, including ZmLEC1 [38], HvFUS3 [39], and OsLFL1 [40], [41], apart from the orthologues of ABI3/VP1. Furthermore, no orthologue of LEC2 has been identified in monocot species [51]. TaL2LA is the first orthologue of LEC2 in monocot. No report has described the association of the expression of four seed maturation regulators with seed dormancy in monocots. Regulatory networks of seed maturation can be compared between monocots and dicots in further investigations.
DOG1 has been identified as a QTL for the regulation of seed dormancy. In fact, DOG1 expression is associated with the geographical variation of seed dormancy in Arabidopsis accessions [35], [52]. Rikiishi and Maekawa [42] reported that TaDOG1 is related to the regulation of seed dormancy in wheat. DOG1 has a RY motif in the promoter. RY motif is a target sequence of B3 domain type transcription factors, suggesting that DOG1 is under the control of LEC2 in Arabidopsis [13], [34], [35], [53]. In the present study, the expression of TaDOG1 showed a significant correlation with dormancy level except for dormant cultivars. Such correlation is similar to that observed with seed maturation regulators. Furthermore, Rikiishi et al. [37] demonstrated that the expression of bZIP type transcription factor, TaABF1 is related to the level of seed dormancy in wheat. LEC1 conferred ABA inducibility to target genes by recruiting ABRE binding factor such as bZIP [54], [55]. TaDOG1 and TaABF1 might act on the regulation of seed dormancy under the control of seed maturation regulators. The regulatory networks of seed maturation might be conserved for the control of seed dormancy in dicot and monocot species. To further elucidate the regulation of seed dormancy in monocots, seed maturation regulators should be investigated in details.
Supporting Information
Alignment of the deduced amino acid sequences of the B domains of LEC1, L1L and their orthologues. Characters filled with black and gray represent perfect matches and more than 50% matches, respectively, among genes.
(TIF)
Alignment of the deduced amino acid sequences of the B3 domains of LEC2, FUS3/TaFUS3, ABI3/TaVP1, and seven LEC2 orthologues. Characters filled with black and gray represent perfect matches and more than 50% matches, respectively, among genes.
(TIF)
Gene-specific primers used for quantitative RT-PCR.
(DOCX)
Accession numbers of LEC1, LEC2, FUS3, ABI3/VP1 and their orthologous genes.
(DOCX)
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by a grant from the Ohara Foundation for Agricultural Research. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Gubler F, Millar AA, Jacobsen JV (2005) Dormancy release, ABA and pre-harvest sproutiong. Curr Opin Plant Biol 8: 183–187. [DOI] [PubMed] [Google Scholar]
- 2. Bewley JD (1997) Seed germination and dormancy. Plant Cell 9: 1055–1066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Pourkheirandish M, Komatsuda T (2007) The importance of barley genetics and domestication in a global perspective. Ann Bot 100: 999–1008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Finkelstein RR, Gampala SSL, Rock CD (2002) Abscisic acid signaling in seeds and seedling. Plant Cell 14: S15–S45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Himmelbach A, Yang Y, Grill E (2003) Relay and control of abscisic acid signaling. Curr Opin Plant Biol 6: 470–479. [DOI] [PubMed] [Google Scholar]
- 6. Nambara E, Marion-Poll A (2003) ABA action and interactions in seeds. Trends Plant Sci 8: 213–217. [DOI] [PubMed] [Google Scholar]
- 7. Kushiro T, Okamoto M, Nakabayashi K, Yamagishi K, Kitamura S, et al. (2004) The Arabidopsis cytochrome P450CYP707A encodes ABA 8’-hydroxylases: key enzymes in ABA catabolism. EMBO J 23: 1647–1656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Saito S, Hirai N, Matsumoto C, Ohigarashi H, Ohta D, et al. (2004) Arabidopsis CYP707As encode (+)-abscisic acid 8’-hydoroxylase, a key enzyme in the oxidative catabolism of abscisic acid. Plant Physiol 134: 1439–1449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Okamoto M, Kuwahara A, Seo M, Kushiro T, Asami T, et al. (2006) CYP707A1 and CYP707A2, which encode abscisic acid 8’-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis. Plant Physiol 141: 97–107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Seo M, Hanada A, Kuwahara A, Endo A, Okamoto M, et al. (2006) Regulation of hormone metabolism in Arabidopsis seeds: phytochrome regulation of abscisic acid metabolism and abscisic acid regulation of gibbellin metabolism. Plant J 48: 354–366. [DOI] [PubMed] [Google Scholar]
- 11. Holdsworth MJ, Bentsink L, Soppe WJJ (2008) Molecular networks regulating Arabidopsis seed maturation, after ripening, dormancy and germination. New Phytol 179: 33–54. [DOI] [PubMed] [Google Scholar]
- 12. Raz V, Bergervoet JHW, Koornneef M (2001) Sequential steps for developmental arrest in Arabidopsis seeds. Development 128: 243–252. [DOI] [PubMed] [Google Scholar]
- 13. Kroj T, Savino G, Valon C, Giraudat J, Parcy F (2003) Regulation of storage protein gene expression in Arabidopsis . Development 130: 6065–6073. [DOI] [PubMed] [Google Scholar]
- 14. Kagaya Y, Toyoshima R, Okuda R, Usui H, Yamamoto A, et al. (2005) LEAFY COTYLEDONE1 controls seed storage protein genes through its regulation of FUSCA3 and ABSCISIC ACID INSENSIVE3. Plant Cell Physiol 46: 399–406. [DOI] [PubMed] [Google Scholar]
- 15. To A, Valon C, Savino G, Guileminot J, Devic M, et al. (2006) A network of local and redundant gene regulation governs Arabidopsis seed maturation. Plant Cell 18: 1642–1651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Giraudat J, Hauge BM, Valon C, Smalle J, Parcy F, et al. (1992) Isolation of the Arabidopsis Abi3 gene by positional cloning. Plant Cell 4: 1251–1261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Luerßen H, Kirik V, Herrmann P, Miséra S (1998) FUSCA3 encodes a protein with a conserved VP1/ABI3-loke B3 domain which is of functional importance for the regulation of seed maturation in Arabidopsis thaliana . Plant J 15: 755–764. [DOI] [PubMed] [Google Scholar]
- 18. Stone SL, Kwong LW, Yee KM, Pelletier J, Lepiniec L, et al. (2001) LEAFY COTYLEDONE2 encodes a B3 domain transcription factor that induces embryo development. Proc Natl Acad Sci USA 98: 11806–11811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Lotan T, Ohto M, Yee KM, West MAL, Lo R, et al. (1998) Arabidopsis LEAFY COTYLEDON1 is sufficient to induce embryo development in vegetative cells. Cell 93: 1195–1205. [DOI] [PubMed] [Google Scholar]
- 20. Meinke DW (1992) A homeotic mutant of Arabidopsis thaliana with leaf cotyledons. Science 258: 1647–1650. [DOI] [PubMed] [Google Scholar]
- 21. Bäumlein H, Miséra S, Luerßen H, Kölle K, Horstmann C, et al. (1994) The FUS3 gene of Arabidopsis thaliana is a regulator of gene expression during late embryogenesis. Plant J 6: 379–387. [Google Scholar]
- 22. Keith K, Kraml M, Dengler NG, McCourt P (1994) fusca3: A heterochronic mutation affecting late embryo development in Arabidopsis. Plant Cell 6: 589–600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Meinke DW, Franzmann LH, Nickle TC, Yeung EC (1994) Leafy cotyledon mutants of Arabidopsis. Plant Cell 6: 1049–1064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. West MAL, Yee KM, Danao J, Zimmerman JL, Fischer RL, et al. (1994) LEAFY COTYLEDON1 is an essential regulator of late embryogenesis and cotyledon identity in Arabidopsis. Plant Cell 6: 1731–1745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Parcy F, Valon C, Kohara A, Miséra S, Giraudat J (1997) The ABSCISIC ACID-INSENSITIVE3, FUSCA3, and LEAFY COTYLEDON1 loci act in concert to control multiple aspects of Arabidopsis seed development. Plant Cell 9: 1265–1277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Nambara E, Hayama R, Tsuchiya Y, Nishimura M, Kawaide H, et al. (2000) The role of ABI3 and FUS3 loci in Arabidopsis thaliana on phase transition from late embryo development to germination. Dev Biol 220: 412–423. [DOI] [PubMed] [Google Scholar]
- 27. Vicient CM, Bies-Etheve N, Delseny M (2000) Changes in gene expression in the leafy cotyledon1 (lec1) and fusca3 (fus3) mutants of Arabidopsis thaliana L. J Exp Bot. 51: 995–1003. [DOI] [PubMed] [Google Scholar]
- 28. Parcy F, Valon C, Raynal M, Gaubier-Comella P, Delseny M, et al. (1994) Regulation of gene expression programs during Arabidopsis seed development: roles of the ABI3 locus and of endogenous abscisic acid. Plant Cell 6: 1567–1582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Tsuchiya Y, Nambara E, Naito S, McCourt P (2004) The FUS3 transcription factor functions through the epidermal regulator TTG1 during embryogenesis in Arabidopsis . Plant J 37: 73–81. [DOI] [PubMed] [Google Scholar]
- 30. Curaba J, Moritz T, Blervaque R, Parcy F, Raz V, et al. (2004) AtGA3ox2, a key gene responsible for bioactive gibberellin biosynthesis, is regulated during embryogenesis by LEAFY COTYLEDON2 and FUSCA3 in Arabidopsis . Plant Physiol 136: 3660–3669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Gazzarrini S, Tsuchiya Y, Lumba S, Okamoto M, McCourt P (2004) The transcription factor FUSCA3 controls developmental timing in Arabidopsis through the hormones gibberellin and abscisic acid. Dev Cell 7: 373–385. [DOI] [PubMed] [Google Scholar]
- 32. Stone SL, Braybrook SA, Paula SL, Kwong LW, Meuser J, et al. (2008) Arabidopsis LEAFY COTYLEDON2 induces maturation traits and auxin activity: implications for somatic embryogenesis. Proc Natl Acad Sci USA 105: 3151–3156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Kwong RW, Bui AQ, Lee H, Kwong LW, Fischer RL, et al. (2003) LEAFY COTYLEDON1-LIKE defines a class of regulators essential for embryo development. Plant Cell 15: 5–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Braybrook SA, Stone SL, Park S, Bui AQ, Le BH, et al. (2006) Genes directly regulated by LEAFY COTYLEDONE2 provide insight into the control of embryo maturation and somatic embryogenesis. Proc Natl Acad Sci USA 103: 3468–3473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Bentsink L, Jowett J, Hanhart CJ, Koornneef M (2006) Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis. Proc Natl Acad Sci USA 103: 17042–17047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Nakabayashi K, Bartsch M, Xiang Y, Hiatton E, Pellengahr S, et al. (2012) The time required for dormancy release in Arabidopsis is determined by DELAY OF GERMINATION1 protein levels in freshly harvested seeds. Plant Cell 24: 2826–2838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Rikiishi K, Matsuura T, Maekawa M (2010) TaABF1, ABA response element binding factor 1, is related to seed dormancy and ABA sensitivity in wheat (Triticum aestivum L.) seeds. J Cereal Sci 52: 236–238. [Google Scholar]
- 38. Shen B, Allen WB, Zheng P, Li C, Glassman K, et al. (2010) Expression of ZmLEC1 and ZmWRI1 increases seed oil production in maize. Plant Physiol 153: 980–987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Moreno-Risueno MA, González N, Díaz I, Parcy F, Carbonero P, et al. (2008) FUSCA3 from barley unveils a common transcriptional regulation of seed-specific genes between cereals and Arabidopsis. Plant J 53: 882–894. [DOI] [PubMed] [Google Scholar]
- 40. Peng L-T, Shi Z-Y, Li L, Shen G-Z, Zhang J-L (2008) Overexpression of transcription factor OsLFL1 delays flowering time in Oryza sativa. J Plant Physiol 165: 876–885. [DOI] [PubMed] [Google Scholar]
- 41. Guo X, Hou X, Fang J, Wei P, Xu B, et al. (2013) The rice GERMINATION DEFECTIVE 1, encoding a B3 domain transcriptional repressor, regulates seed germination and seedling development by integrating GA and carbohydrate metabolism. Plant J 75: 403–416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Rikiishi K, Maekawa M (2010) Characterization of a novel wheat (Triticum aestivum L.) mutant with reduced seed dormancy. J Cereal Sci 51: 292–298. [Google Scholar]
- 43. Paolacci AR, Tanzarella OA, Porceddu E, Ciaffi M (2009) Identification and validation of reference genes for quantitative RT-PCR normalization in wheat. BMC Mol Biol 10: 11 dot:10.1186/1471-2199-10-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Lee H, Fischer RL, Goldberg RB, Harada JJ (2003) Arabidopsis LEAFY COTYLEDON1 represents a functionally specialized subunit of the CCAAT binding transcription factor. Proc Natl Acad Sci USA 100: 2152–2156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. McKibbin RS, Wilkinson MD, Bailey PC, Flintham JE, Andrew LM, et al. (2002) Transcripts Vp-1 homeologues are misspliced in modern wheat and ancestral species. Proc Natl Acad Sci USA 99: 10203–10208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Swaminathan K, Peterson K, Jack T (2008) The plant B3 superfamily. Trends Plant Sci 13: 647–655. [DOI] [PubMed] [Google Scholar]
- 47. Baumbusch LO, Hughes DW, Galau GA, Jakobsen KS (2004) LEC1, FUS3, ABI3 and Em expression reveals no correlation with dormancy in Arabidopsis . J Exp Bot 55: 77–87. [DOI] [PubMed] [Google Scholar]
- 48. Alonsi-Blanco C, Bentsink L, Hanhart CJ, Vries HB, Koornneef M (2003) Analysis of natural allelic variation at seed dormancy loci of Arabidopsis thaliana . Genetics 164: 711–729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Jones HD, Peters NCB, Holdsworth MJ (1997) Genotype and environment interact to control dormancy and differential expression of the VIVIPAROUS 1 homologue in embryos of Avena fatua . Plant J 12: 911–920. [DOI] [PubMed] [Google Scholar]
- 50. Liu A, Gao F, Kanno Y, Jordan MC, Kamiya Y, et al. (2013) Regulation of wheat seed dormancy by after-ripening is mediated by specific transcriptional switches that induce changes in seed hormone metabolism and signaling. PLoS ONE 8(2): e56570 doi:10.1371/journal.pone.0056570 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Sreenivasulu N, Wobus U (2013) Seed-development programs: A systems biology-based comparison between dicots and monocots. Annu Rev Plant Biol 64: 189–217. [DOI] [PubMed] [Google Scholar]
- 52. Chiang GCK, Bartsch M, Barua D, Nakabayashi K, Debieu M, et al. (2011) DOG1 expression is predicted by the seed-maturation environment and contributes to geographical variation in germination in Arabidopsis thaliana . Mol Ecol 20: 3336–3349. [DOI] [PubMed] [Google Scholar]
- 53. Reidt W, Wohlfarth T, Ellerstrom M, Czihal A, Tewes A, et al. (2000) Gene regulation during late embryogenesis: the RY motif of maturation-specific gene promoters is a direct target of the FUS3 gene product. Plant J 21: 401–408. [DOI] [PubMed] [Google Scholar]
- 54. Yamamoto A, Kagaya Y, Toyoshima R, Kagaya M, Takeda S, et al. (2009) Arabidopsis NF-YB subunits LEC1 and LEC1-LIKE activate transcription by interacting with seed-specific ABRE-binding factors. Plant J 58: 843–856. [DOI] [PubMed] [Google Scholar]
- 55. Junker A, Mönke G, Rutten T, Keilwagen J, Seifert M, et al. (2012) Elongation-related functions of LEAFY COTYLEDON1 during the development of Arabidopsis thaliana. Plant J 71: 427–442. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Alignment of the deduced amino acid sequences of the B domains of LEC1, L1L and their orthologues. Characters filled with black and gray represent perfect matches and more than 50% matches, respectively, among genes.
(TIF)
Alignment of the deduced amino acid sequences of the B3 domains of LEC2, FUS3/TaFUS3, ABI3/TaVP1, and seven LEC2 orthologues. Characters filled with black and gray represent perfect matches and more than 50% matches, respectively, among genes.
(TIF)
Gene-specific primers used for quantitative RT-PCR.
(DOCX)
Accession numbers of LEC1, LEC2, FUS3, ABI3/VP1 and their orthologous genes.
(DOCX)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.