
Figure Five. Vinculin positive focal adhesions are colocalized to areas of altered membrane topography and composition.
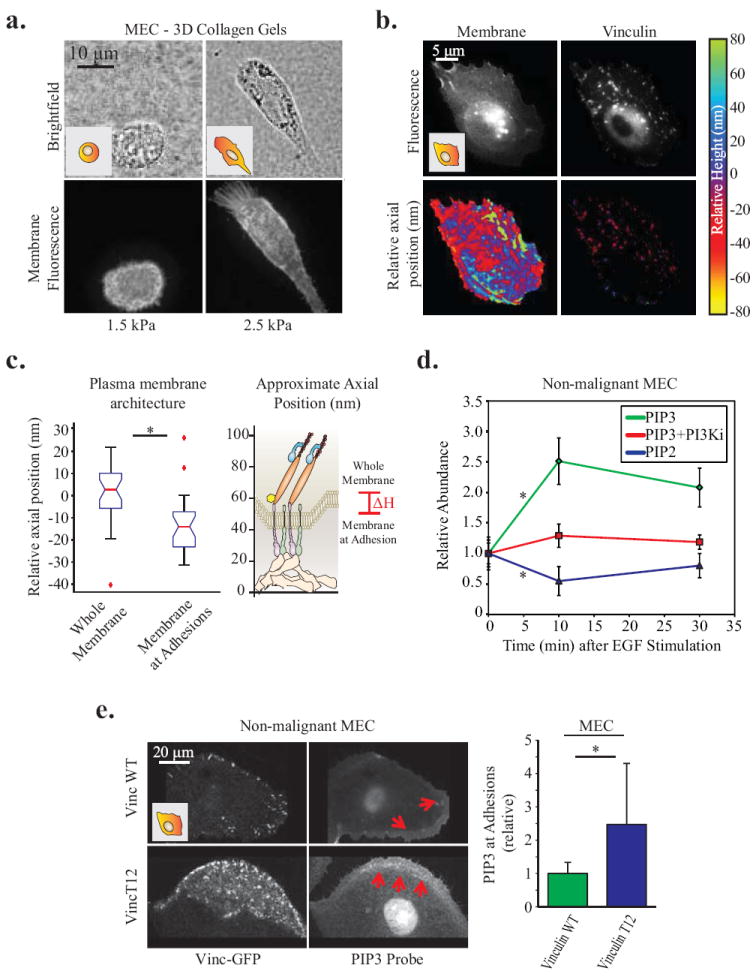
A. Non-malignant MEC in 3D collagen gels for 24hrs. Membrane protrusions are dependent on high (2.5 kPa) matrix stiffness; visualized via mEmerald-farnesyl membrane probe. B. Organization of vinculin and the ventral plasma membrane of MEC plated on a 2D glass surfaces, via SAIM measurements of vinculin-GFP and mCherry-farnesyl. Axial position is plotted relative to the mean height of the membrane and vinculin, respectively. C. Quantification of the significant decrease in membrane axial position at focal adhesions (p < .01; ±SD, n>18 cells per condition). D. Line graphs showing relative abundance of PIP2 and PIP3 in the ventral membrane of non-malignant MEC between 0-30 minutes following EGF stimulation. 10 minutes after stimulation PIP2 significantly decreases and PIP3 significantly increases. However, inhibiting PI3K activity (1μM GDC-0941) is able to abrogate this increase in PIP3 (p < .05; ±SE, n>25 cells per condition). E. Confocal images of pre-malignant MEC plated on 2D PA gels of increasing stiffness expressing vinculin-GFP activity mutants and a probe for PIP3 activity (mKO2-PH-Grp1), where red arrows indicate areas enriched in PIP3 activity. Measurements of all pixels in adhesions were averaged over whole cell; PIP3 was significantly enriched in focal adhesions of cells expressing activated vinculin T12 (p < .05; ±SD, n>7 cells per condition).