
Figure 1. Mechanisms of colorectal cancer initiation and dissemination.
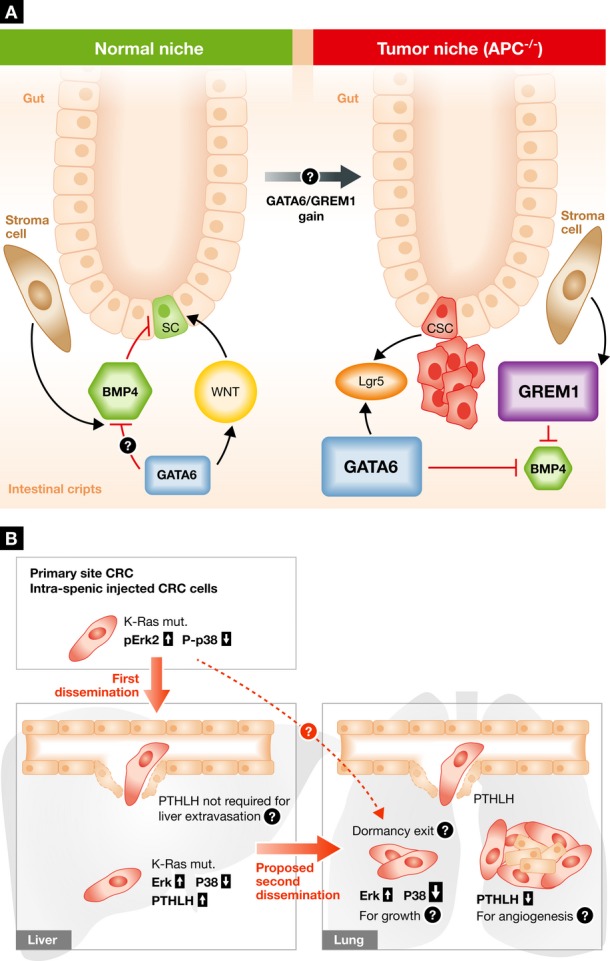
(A) The normal intestinal crypt niche is maintained by a continuous balance between growth-promoting (i.e., WNT) and inhibitory signals (i.e., BMPs). Batlle and co-workers show that in an APC, null background GATA6 signals promote tumorigenesis via inhibition of BMP4 expression and function in cancer stem cells (CSC). This effect is achieved by a competition of GATA6 for the same binding elements of β-catenin/TCF4 factors and by modifying the epigenetic landscape of the BMP4 promoter. Importantly, in patients, germ line gene duplication of GREM1, a BMP inhibitor, is linked to increased predisposition to CRC development. (B) High phospho-ERK2 is required for CRC metastatic seeding to the liver from the spleen or primary site, whereas metastatic seeding of the lung from the liver requires a strong downregulation of p38 activation. Loss of P-p38 also induces PTHLH secretion that facilitates cancer cell dissemination in the lung by killing the endothelial cells. It remains to be elucidated how PTHLH is not required for liver extravasation, how primary tumor endothelium and that associated with metastasis become insensitive to PTHLH, and whether P-ERK2 and P-p38 levels also affect dormancy and reactivation of disseminated tumor cells in this model.