

Abstract
China is the world's largest producer and consumer of fertilizer N, and decades of overuse has caused nitrate leaching and possibly soil acidification. We hypothesized that this would enhance the soils' propensity to emit N2O from denitrification by reducing the expression of the enzyme N2O reductase. We investigated this by standardized oxic/anoxic incubations of soils from five long-term fertilization experiments in different regions of China. After adjusting the nitrate concentration to 2 mM, we measured oxic respiration (R), potential denitrification (D), substrate-induced denitrification, and the denitrification product stoichiometry (NO, N2O, N2). Soils with a history of high fertilizer N levels had high N2O/(N2O+N2) ratios, but only in those field experiments where soil pH had been lowered by N fertilization. By comparing all soils, we found a strong negative correlation between pH and the N2O/(N2O+N2) product ratio (r2 = 0.759, P < 0.001). In contrast, the potential denitrification (D) was found to be a linear function of oxic respiration (R), and the ratio D/R was largely unaffected by soil pH. The immediate effect of liming acidified soils was lowered N2O/(N2O+N2) ratios. The results provide evidence that soil pH has a marginal direct effect on potential denitrification, but that it is the master variable controlling the percentage of denitrified N emitted as N2O. It has been known for long that low pH may result in high N2O/(N2O+N2) product ratios of denitrification, but our documentation of a pervasive pH-control of this ratio across soil types and management practices is new. The results are in good agreement with new understanding of how pH may interfere with the expression of N2O reductase. We argue that the management of soil pH should be high on the agenda for mitigating N2O emissions in the future, particularly for countries where ongoing intensification of plant production is likely to acidify the soils.
Keywords: agriculture, denitrification, fertilizer, Nitrous oxide, soil acidification
Introduction
China has become the world's largest producer and consumer of fertilizer nitrogen (Cui et al., 2013), and the emission of greenhouse gases associated with the N fertilizer chain in China was recently assessed by Zhang et al. (2013). They estimate total emissions in 2010 to be 452 Tg CO2-equivalents, of which 61% is due to emissions from the production of fertilizers (including CO2 and CH4 from coal mining), and around 32% is ascribed to emissions of N2O from soils. Their scenario analyses for emissions during the next two decades suggest that substantial reductions can be achieved by improved technology in the fertilizer industry, and by reductions in N fertilizer rates. The latter is possible without reducing yields, since Chinese farmers use much more mineral N than that needed by crops (Ju et al., 2004; Fan et al., 2005; Jiang et al., 2006; Zhao et al., 2006; Qiu et al., 2012). Although substantial reductions in N fertilizer levels are technically feasible without jeopardizing food security for the Chinese population, it appears difficult to realize for a number of societal/political reasons (Zhang et al., 2013). The most likely scenario for decades to come is thus that China will continue to use excessive amounts of fertilizers.
Nationwide estimations of fertilizer induced N2O emissions are most commonly based on emission factors (a constant fraction of the fertilizer N is emitted as N2O-N). Zhang et al. (2013) are no exception in this respect, although they base their emission factors (1.05% for upland soils and 0.41% for paddy fields) on measured emissions from Chinese field experiments (instead of IPCC's default values, 1% for upland soils and 0.3% for paddy fields; IPCC, 2006). This is indisputably the best option for assessing current emissions, but possibly not for predicting future emissions (scenarios). The reason for this is that the fraction of fertilizer N emitted as N2O may change gradually depending on agronomic practice. We believe that this is particularly relevant for China, for reasons developed in some detail below.
Guo et al. (2010) have documented that the combination of increasing fertilizer levels and crop yields over the last decades has resulted in a significant acidification of Chinese agricultural soils. Their calculations of proton balances and base cation removal for Chinese croplands indicate that an accelerating acidification will take place in the years to come if current agronomic practice is continued. This is possibly bad news regarding the future emissions of N2O from Chinese agricultural soils.
In soils, the only known sink for N2O is the enzyme N2O-reductase, which is one of four reductases used by denitrifying bacteria to sustain respiration under anoxic conditions by reducing NO3− to N2 (van Spanning et al., 2007). Empirical evidence for a negative effect of acidity on the rate of N2O reduction in soil has accumulated over the last five decades since it was first observed by Wijler & Delwiche (1954) and Nömmik (1956); as summarized by Šimek & Cooper (2002). The mechanisms involved were not well-understood, however, and the strength of this pH control over the N2O/(N2O+N2) product ratio of denitrification was not adequately determined when compared with other factors. Recent investigations of gene transcription and electron flow kinetics at the cellular level with the model organism Paracoccus denitrificans have indicated that the reason for low N2O reduction rate at low pH is a post transcriptional interference during the assembly of the enzyme N2O reductase (Bergaust et al., 2010; Bakken et al., 2012). Liu et al. (2010) conducted a series of experiments with soil from long-term liming experiments in Scandinavia, and demonstrated a strong negative correlation between soil pH and the N2O/(N2O+N2) ratio within the pH range 4–7. Measurements of gene transcription (nir, nor, and nosZ, coding for NO2−, NO. and N2O-reductase respectively), along with the gas kinetics demonstrated that the low N2O reductase activity in acid soils could not be ascribed to low number of nosZ transcripts. The authors suggested that the N2O/(N2O+N2) was negatively correlated with pH for exactly the same reason as in the model organism P. denitrificans. Thus, soil pH emerges as a master variable for N2O production in soils, and we are beginning to understand why. The phenomenon may have major implications for future emissions from Chinese agriculture, and this needs to be evaluated by investigating soils from relevant long-term experiments in China.
We hypothesized that the excessive application of nitrogen fertilizer in the intensive cropping system in China (i) results in soil acidification; (ii) that this soil acidification induces higher N2O/(N2O+N2) product ratio; (iii) that liming alleviates the effect of acidification on the N2O/(N2O+N2) ratio. Firstly, we sampled the soils from five Chinese long-term fertilization experiment sites, with various fertilizer N levels, and with or without manure input. The soils were incubated in a robotized incubation system (Molstad et al., 2007) under oxic and subsequently anoxic conditions [in helium (He) atmosphere], during which the O2, CO2, NO, N2O, and N2 gas kinetics were monitored. Secondly, the immediate effect of liming was tested using the same incubation procedure.
Materials and methods
Soils and field experiments
Five long-term field fertilization experimental sites with upland crops were selected from cold temperate to subtropical regions of the major agricultural areas in China (Fig.1). Gongzhuling (GZL) soils were taken from an experimental field established in 1980 at the Gongzhuling Research Farm of the Jilin Academy of Agricultural Sciences, Jilin Province. The soils are classified as Luvic Phaeozems in FAO (1988) and typic Black soil according to the Chinese Soil Classification System (Zhu et al., 2007; Zhang et al., 2009). Shangzhuang (SZ) soils were sampled from a newly established field experiment that was started in October 2006 at the Shangzhuang Research Station of China Agricultural University, suburban Beijing. The soil type is Calcareous Cambisol (FAO, 1988; Qiu et al., 2012). Qiyang (QY) soils were sampled from the plots of a long-term experiment established in September 1990 at the Qiyang Experiment Station of the Chinese Academy of Agricultural Sciences (CAAS), Hunan Province. The soils are classified as Ferralic Cambisol (FAO, 1988; Zhang et al., 2008). Changwu (CW) soils were taken from a long-term fertilization experiment started in 1984 at the Changwu Agri-ecological Station, Changwu County, Shaanxi Province. The soil is a calcareous soil, and referred to as a coarse-textured dark loessial soil following the Chinese Soil Classification System, which corresponds to a Calcarid Regosol based on the FAO System (FAO, 1988; Hao et al., 2007). Shouguang (SG) soils were taken from a greenhouse experiment started in 2004 at Luojia village, Shouguang, Shandong province. The soil is classified as Calcareous Cambisol (FAO, 1988; He et al., 2009). More information is described in Table1. Soil samples were taken throughout 2010 and 2011, transported to the laboratory in Norway where they were passed through a 4 mm sieve and stored at 4 °C in moist condition (pF = 2–3) until use (≤12 months after sampling).
Figure 1.
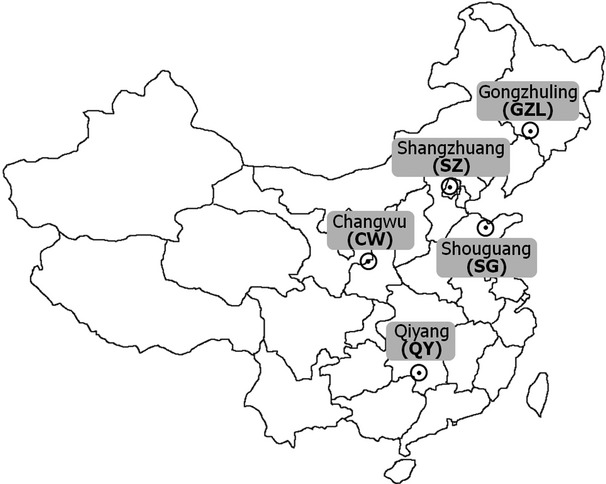
Map showing the sampling sites in China.
Table 1.
Field experimental details
| Gongzhuling (GZL) | Shangzhuang (SZ) | Qiyang (QY) | Changwu (CW) | Shouguang (SG) | |
|---|---|---|---|---|---|
| Latitude | 42°57′N | 39°48′N | 26°45′N | 35°14′N | 36°55′N |
| Longitude | 148°57′E | 116°28′E | 111°52′E | 107°41′E | 118°45′E |
| Altitude (m) | 150–220 | 40 | 120 | 1200 | 22 |
| Mean annual precipitation (mm) | 450–600 | 500–700 | 1255 | 565 | 558 |
| Mean annual temperature (°C) | 4–5 | 11.4 | 18 | 9.1 | 12.4 |
| Soil texture | Silt loam | Silt loam | Clay | Clay loam | Silt loam |
| 31% clay | 20% clay | 61.4% clay | 33% clay | 2% clay | |
| 30% silt | 57% silt | 34.9% silt | 21% silt | 52% silt | |
| 39% sand | 23% sand | 3.7% sand | 46% sand | 46% sand | |
| Organic C (g kg−1) | 12. 9–24.0 | 3.3–5.4 | 4.3–12.0 | 4.3–9.5 | 3.4–10.3 |
| pH (1:5 w/v in 0.01 mol l−1 CaCl2) | 7.3–7.7 | 6.7 | 3.7–5.7 | 7.9–8.0 | 6.2–7.3 |
| Cropping system | Maize | Wheat-maize | Wheat-maize | Wheat | Two-season tomato |
| Year of establishment | 1980 | 2006 | 1990 | 1984 | 2004 |
| Fertilizer treatment (N, P, K, M)* | |||||
| CK | 0, 0, 0, 0 | 0, 0, 0, 0 | 0, 0, 0, 0 | 0, 0, 0, 0 | 0, 0, 0, 0 |
| Nopt | 165, 36, 57, 0† | 220, 70, 75, 0 | 300, 53, 100, 0† | 120, 0, 0, 0 | – |
| Nhigh | – | 560, 70, 75, 0 | – | – | – |
| M | 0, 0, 0, 60 | – | 0, 0, 0, 27 | 0, 0, 0, 75 | 0, 0, 0, 36 |
| M Nopt | 165, 36, 57, 60† | – | 90, 53, 100, 19 | 120, 0, 0, 75 | 368, 79, 382, 36 |
| M Nhigh | – | – | – | – | 1290, 79, 382, 36 |
| Type of manure | Mixture of swine manure, plant residues, and soil. | – | Pig manure 1.1% N | Cow manure | Chicken manure 1.5% N |
| 10–20% C | 10% C | ||||
| 0.5–1% N | 0.26% N | ||||
The average annual amount of nitrogen (kg N ha−1), phosphorus (kg P ha−1), potassium (kg K ha−1), and farmyard manure (Mg ha−1) input.
A treatment without P and K (but identical N level) was included.
To simplify the presentation and discussion of treatment effects, we combined various treatments into broad categories with the following codes
CK: control, no manure, or chemical fertilizer input
Nhigh: high doses of chemical fertilizer N input (with or without P and K)
Nopt: moderate/optimal doses of chemical fertilizer N input (with or without P and K)
M: manure input
MNhigh/opt: manure and chemical fertilizer N input (with or without P and K)
Experimental details for each field experiment are given in Table S1
Soil pretreatment
To obtain a homogenous distribution and equal concentration of nitrate in all soils, the soil samples were treated by flooding and drainage with 2 mm KNO3 solution immediately before incubation. The soil samples were placed in 500 ml filter funnels with 4.5 cm diameter 0.45 μm Millipore membrane filters and flooded with 2 mm KNO3 solution. After 5 min the soils were drained by applying vacuum for 10 min. This flooding/drainage procedure was repeated three times, followed by a final evacuation step of 1 h. The soil moisture content, based on soil wet weight, ranged from 15% to 25% after the final drainage, depending on soil texture.
Soil incubation under oxic and anoxic conditions
We used the same incubation procedures as Raut et al. (2012), with minor modifications. After flooding and drainage, the soils were transferred to 120 ml serum vials (30 g fresh weight per vial), which were then sealed with air-tight butyl-rubber septa and aluminum crimp caps. Two or three replicate vials were prepared depending on the availability of the incubation positions in the incubation system. All the vials were transferred to the robotized incubation system at 15 °C to monitor gases (O2, CO2, NO, N2O, and N2) under oxic conditions for a period of 24–40 h to estimate the respiration rate. This period was named Phase Oxic.
After this oxic incubation, all the vials were repeatedly evacuated and filled with He (6 cycles) to remove O2 and N2. The vials were then transferred back to the incubation system and incubated at 15 °C to monitor headspace gases. This anoxic incubation period was named Phase I, which lasted 100–200 h depending on the denitrification rate; for soils with high denitrification rates, Phase I was ended when all NO3− was recovered as N2 or N2+N2O (ca. 100% recovery). For soils with lower rates of denitrification, Phase I was ended despite the presence of residual NO3− and NO2− in the soil, i.e. (N2O+N2)−N < initial NO3−N. Subsequently, nitrogen and glutamate was added by injecting a 0.5 ml solution (spread by dripping evenly on the soil surface) containing 26 mm KNO3 and 130 mm glutamic acid, with pH adjusted to 7 with sodium hydroxide (NaOH). This injection gave a final concentration of approximately 2 mm KNO3 and 10 mm glutamate in the soil liquid (if evenly distributed). The vials were then He washed once more and continually incubated anoxic until the N2 gas production stopped due to depletion of nitrate (such depletion was not reached for the soils with the lowest denitrification rates). This period was named Phase II. O2, CO2, NO, N2O, and N2 gases were monitored automatically during these two anoxic incubation periods.
The incubation experiments were done in a robotized incubation system similar to that described by Molstad et al. (2007), with moderate improvements. In short, the system monitored the headspace-concentrations of relevant gases (O2, CO2, NO, N2O, and N2) by repeated gas sampling through the butyl-rubber septa of the incubation vials. The gas samples were drawn by a peristaltic pump, which returned an equal volume of He. This ensured that the gas pressure was sustained near 1 atm despite repeated sampling, but diluted the headspace atmosphere with He (ca. 1%). This dilution was taken into account when calculating rates of production/consumption for each time increment [see Molstad et al. (2007) for details]. The sampling robot (Agilent GC Sampler 80) had a larger operating width than the previous version, and could sample from 44 incubation vials in the thermostatic water bath. NO concentrations were determined by the same instrument as in the original system (Chemoluminiscence NOx analyser Model 200A, Advanced Pollution Instrumentation, San Diego, USA), but the microGC with only thermal conductivity detectors (TCD) was replaced with a regular gas chromatograph (Agilent GC -7890A) with three detectors: a flame ionization detector (FID), a thermal conductivity detector (TCD), and an electron capture detector (ECD). N2O was detected by both the ECD (linear range 0–20 μl l−1) and TCD (for concentrations >20 μl l−1). The GC had two 30 m 0.53 mm id columns: a PLOT column for separation of CH4, CO2, and N2O, and a Molsieve column for separation of O2 and N2 (and Ar and Ne). Three vials with standard gases (known concentrations) were included in each incubation for calibration. These standard vials were prepared by evacuating 120 ml sealed vials and then filling them with Low standard gas (361 ppmv CO2, 1.89 ppmv CH4, and 0.585 ppmv N2O), High standard gas (1% CO2, 1% CH4, and 151 ppmv N2O) and NO standard gas (24.9 ppmv NO) respectively.
Gas kinetics
The oxic respiration rates were calculated from measured accumulation of CO2 [taking carbonate chemistry as a function of soil pH into account, see Molstad et al. (2007) for details]. The reason for not using O2 consumption rates was that estimated O2 consumption rates for single time increments were very variable due to low rates compared with the loss of O2 from the headspace by sampling. The denitrification rates were calculated by total (NO+N2O+N2)−N production during anoxic incubations. All these rates were calculated from changes in the headspace concentrations for each time increment between two samplings, taking into account the losses by sampling and minor leakage of N2 into the vials, as described in detail by Molstad et al. (2007).
To characterize the denitrification product ratio N2O/(N2O+N2) during anoxic incubations, we calculated an N2O production index (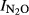 ), as described by Liu et al. (2010). The index was calculated for each vial and used as single observation in the statistical analyses. The formula for the index is
), as described by Liu et al. (2010). The index was calculated for each vial and used as single observation in the statistical analyses. The formula for the index is
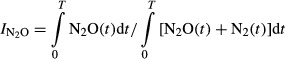 |
1 |
where N2O(t) is the accumulated flux of N2O at any time t, N2(t) is the accumulated flux of N2 at any time t, and T is the time when a specified amount of NO3−N had been recovered as (NO+N2O+N2)-N. To include all soils in a statistical evaluation of treatment effects, we calculated 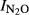 at T, where T = the time when 100 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N. For the soils with the lower denitrification rates, this was the maximum reached before termination of Phase I. The integrals were calculated by trapezoidal integration for each time interval (between two gas samplings). As shown in Fig.2, the accumulated N2O flux [i.e. N2O(t)] did not reach zero despite zero concentration in the vials. The reason for this is that a fraction of the N2O was removed by sampling, and the bacteria cannot reduce this N2O for obvious reasons.
at T, where T = the time when 100 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N. For the soils with the lower denitrification rates, this was the maximum reached before termination of Phase I. The integrals were calculated by trapezoidal integration for each time interval (between two gas samplings). As shown in Fig.2, the accumulated N2O flux [i.e. N2O(t)] did not reach zero despite zero concentration in the vials. The reason for this is that a fraction of the N2O was removed by sampling, and the bacteria cannot reduce this N2O for obvious reasons.
Figure 2.
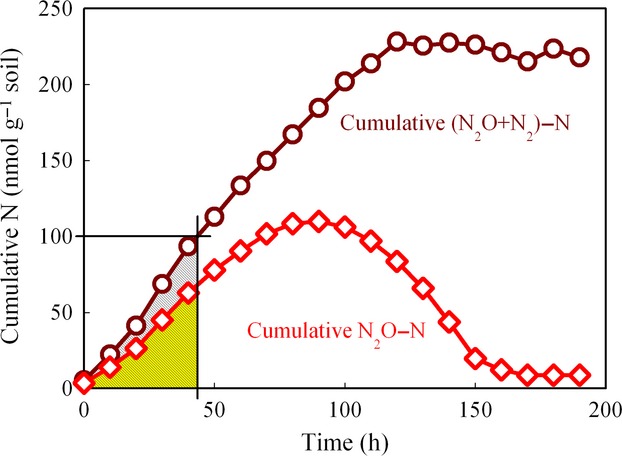
Demonstration of calculation of N2O production index (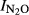 = ∫ N2O(t)dt/∫ [N2O(t)+N2(t)]dt). The area under the cumulative N2O−N is ∫ N2O(t)dt, and the area under the cumulative (N2O+N2)−N is ∫ [N2O(t)+N2(t)]dt, where N2O(t) is the accumulated flux of N2O at any time t, and N2(t) is the accumulated flux of N2 at any time t.
= ∫ N2O(t)dt/∫ [N2O(t)+N2(t)]dt). The area under the cumulative N2O−N is ∫ N2O(t)dt, and the area under the cumulative (N2O+N2)−N is ∫ [N2O(t)+N2(t)]dt, where N2O(t) is the accumulated flux of N2O at any time t, and N2(t) is the accumulated flux of N2 at any time t.
Statistical analyses were performed using Minitab 16 (Minitab Inc., State College, PA, USA). One-way anova was used to analyze the long-term fertilization effects on soil pH, soil respiration rates, denitrification rates, and N2O production indexes. General Regression Analysis was used to build linear regression models.
Liming experiment
Four soils were chosen for this experiment: two Chinese soils [MNhigh treatment in Shouguang (SG) and the CK treatment in Qiyang (QY)] and two peat soils (Fjaler soils, pH 4 and 6). The Fjaler peat soils were taken from one of the Norwegian long-term liming experiments analyzed by Liu et al.(2010). In this field experiment, the soil pH had been increased by adding shell sand in 1978. The experiment and the soils are described in detail by Mørkved et al. (2007).
In a preliminary experiment, we determined the amount of CaCO3 needed to raise the pH of each soil to ca. 7. This was done by monitoring soil pH over a period of 2 days after adding various amounts of CaCO3 (data not shown). The amounts of CaCO3 needed were 0.090, 0.038, 0.024, and 0.013 g CaCO3 g−1 dw soil for Fjaler soil with pH4 (F4), Fjaler soil with pH6 (F6), Shouguang soil (SG), and Qiyang soil (QY) respectively. The moist soils were then amended with these doses of CaCO3 and incubated under oxic conditions at 15 °C for 2 days to stabilize the soil pH (parallel soil samples without CaCO3 were included). The soils (with and without CaCO3) were then flooded and drained as described above. Two replicate samples of each treatment (soil with and without CaCO3) were flooded with 2 mm KNO3 solution; and two were flooded with a solution containing both 2 mm KNO3 and 10 mm sodium glutamate (pH adjusted to 7). Immediately after the last drainage, the soils were incubated in He-atmosphere as described above (the Phase Oxic was omitted in this experiment), for determination of denitrification rates and NO/N2O/N2 gas kinetics. The soil pH after adding CaCO3 and flooding/drainage are shown in Table2.
Table 2.
pH of the soils with/without liming, measured in 0.01 mol l−1 CaCl2 (1:5 w/v)
| Soil sample | No lime | Lime added | ||
|---|---|---|---|---|
| No glutamate | With glutamate | No glutamate | With glutamate | |
| Fjaler pH4 (F4) | 3.60 | 3.77 | 6.71 | 6.95 |
| Fjaler pH6 (F6) | 6.18 | 5.96 | 6.81 | 6.97 |
| Shouguang (SG) | 6.68 | 6.71 | 7.06 | 7.20 |
| Qiyang (QY) | 5.14 | 5.20 | 6.89 | 7.12 |
Results
Gas kinetics
A typical case of O2, CO2, NO, N2O, and N2 gas kinetics during incubation is shown in Fig.3. In this figure, cumulative CO2 and N2 are plotted [corrected for dilution and leakage, see Molstad et al. (2007) for details], whereas O2, NO, and N2O are plotted as measured because the accumulated flux of such intermediates (as plotted in Fig.2 for N2O) do not reach zero despite zero concentrations in the vials (as explained in relation to Fig.2).
Figure 3.

Demonstration of O2, CO2, NO, N2O, and N2 gas kinetics during three incubation phases (Phase Oxic, Phase I, and Phase II). The results shown are single vial results for a soil with high pH (Shouguang MNhigh treatment with pH 6.46; top panel) and a soil with low pH (Qiyang CK treatment, pH 4.88; lower panel). The O2 concentration during Phase Oxic is shown as vol% in the headspace, CO2 and N2 are shown as cumulative production (nmol g−1 soil dry weight), whereas the intermediate denitrification products NO and N2O are shown as measured (nmol g−1 soil dry weight).
Three phases during the incubations were defined as shown in Fig.3. For Phase Oxic, only O2 and CO2 concentrations are plotted. NO and N2O remained below 0.2 and 0.4 nmol N g−1 soil in this phase, respectively; N2-production could not be estimated due to the high N2 concentration in air. During 24–40 h aerobic incubation, oxic respiration consumed only marginal fractions of the headspace O2. On average, the molar ratio between O2 consumption and CO2 production was close to 1, but the estimated O2 consumption for single vials was variable since the respiratory oxygen consumption is marginal compared with the O2 lost by sampling. Thus, CO2 production rates were used to obtain single vial estimates of oxic respiration. In the anaerobic incubation Phase I and Phase II, the gas kinetics of soils with different pH showed profoundly different patterns. In the high pH soil (Fig.3a), NO and N2O peaked and declined to zero before or at the same time as the cumulative N2 reached a stable plateau, i.e. 100% recovery of NO3−N as N2. In the low pH soil (Fig.3b), NO reached a relatively stable low value that lasted throughout Phase I, whereas N2O concentrations increased gradually throughout Phase I. The cumulative N2 production remained very low until the very end of Phase I. In Phase II (after adding more nitrate and glutamate), the patterns of NO, N2O, and N2 gas kinetics were similar to those in Phase I in the high pH soil (Fig.3a), although the rates were faster. In Phase II for the low pH soil (Fig.3b), NO reached higher levels than in Phase I. The NO concentration remained stable for more than 100 h, and then dropped to zero. This coincided with the depletion of N2O and the reduction of N2-production rate (near 100% recovery of added NO3−N).
Denitrification rates vs. oxic respiration rates
To inspect the relationship between the rate of oxic respiration and denitrification (Phase Oxic and Phase I), we calculated the rate of electron flow to oxygen during the oxic phase (Re−) based on the accumulated CO2 production, and the rate of electron flow through denitrification (De−) during the first 20 h of the subsequent anoxic phase (Phase I) based on the sum of the net reduction of NO3− to NO, N2O, and N2. We assumed that the production of one CO2 -molecule consumed four electrons during oxic respiration. For denitrification, the reduction of one NO3− molecule to NO consumes three electrons; the reduction of two NO−molecule to N2O consumes two electrons, and the reduction of one N2O - molecule to N2 consumes two electrons. Plotting De− against Re− (Fig.4) demonstrates that De− was a linear function of Re−: De− = 0.55*Re− (r2 = 0.875, P < 0.001). The treatments with manure generally had both higher oxic respiration rates and denitrification rates than the other treatments, but the relationship between De− and Re− appears to be unaffected. The oxic respiration rates (Rox) correlated with the measured soil organic carbon (SOC), although one site (GZL) showed a lower ratio (Rox/SOC) than the other sites (see Fig. S1).
Figure 4.
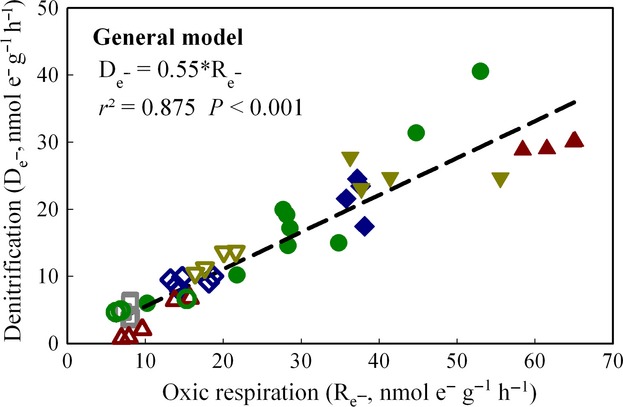
Relationship of the electron flow rates in Phase Oxic respiration (Re−) and in Phase I denitrification (De−). Data for single vials with soil from Gongzhuling (GZL,  ), Shangzhuang (SZ,
), Shangzhuang (SZ,  ), Qiyang (QY,
), Qiyang (QY,  ), Changwu (CW,
), Changwu (CW,  ), and Shouguang (SG,
), and Shouguang (SG,  ) are shown. The closed symbols show the treatments with manure; the open symbols show the treatments without manure. Linear regression function is shown (De− = 0.55*Re−; r2 = 0.875, P < 0.001).
) are shown. The closed symbols show the treatments with manure; the open symbols show the treatments without manure. Linear regression function is shown (De− = 0.55*Re−; r2 = 0.875, P < 0.001).
The ratio De−/Re− is a proxy for the fraction of respiring cells which are able to switch to anoxic respiration in response to oxygen depletion. We inspected if soil pH had any effect on this ratio by plotting the ratios against measured soil pH. The result (Fig.5) shows low ratios for some of the very acidic soils. Overall there seems to be a relationship between soil pH and the De−/Re− ratio, but this could be due to differences between sites regarding drainage for instance.
Figure 5.
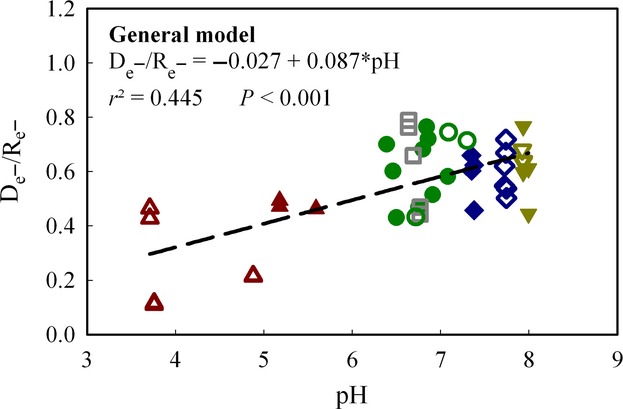
pH effect on the ratio of the electron flow rates in Phase Oxic respiration (Re−) and in Phase I denitrification (De−). Data for single vials with soil from Gongzhuling (GZL,), Shangzhuang (SZ, ), Qiyang (QY, ), Changwu (CW, ), and Shouguang (SG, ) are shown. The closed symbols show the treatments with manure; the open symbols show the treatments without manure.
Effects of long-term fertilization on soil pH and the N2O production index
Fertilization had influenced the soil pH in some but not all the field sites (Table3). In Gongzhuling (GZL), where the background soil pH was high (pH 7.74 ± 0.13 in CK), manure treatment (M and MN) had lowered the soil pH significantly (M and MN vs. CK) whereas mineral N alone had no effect. In Shangzhuang (SZ), fertilization had marginal effects on pH (the two fertilizer treatments had pH within ± 0.08 of that in CK, and none of the contrasts were significant), possibly reflecting the short duration of this field experiment (4 years). In Qiyang (QY), where the soil was naturally acidic (pH 4.86 ± 0.02 for CK), manure treatments had higher pH than in the control soil (CK), whereas mineral N had significantly lowered the pH both in the treatments with manure (MNopt vs. M) and in treatments without manure (Nopt vs. CK). In Changwu (CW), all treatments had high pH (7.92–7.96), apparently unaffected by the fertilization history. In Shouguang (SG), manure alone had not changed soil pH significantly (M vs. CK), but the combination of manure and high mineral N input (MNhigh) had lowered the pH significantly, despite the relatively short duration of this experiment (7 years).
Table 3.
Long-term fertilization effects on soil pH at each site
| Fertilization treatment | Gongzhuling (GZL) 30 years | Shangzhuang (SZ) 4 years | Qiyang (QY) 20 years | Changwu (CW) 26 years | Shouguang (SG) 7 years |
|---|---|---|---|---|---|
| CK | 7.74 ± 0.13*a† | 6.70 ± 0.01a | 4.86 ± 0.02c | 7.93 ± 0.06a | 7.04 ± 0.29a |
| Nopt | 7.73 ± 0.07a | 6.76 ± 0.01a | 3.75 ± 0.04d | 7.94 ± 0.09a | – |
| Nhigh | – | 6.62 ± 0.02a | – | – | – |
| M | 7.38 ± 0.04b | – | 5.66 ± 0.11a | 8.00 ± 0.06a | 6.93 ± 0.14a |
| MNopt | 7.35 ± 0.16b | – | 5.22 ± 0.06b | 7.94 ± 0.04a | 6.82 ± 0.06a |
| MNhigh | – | – | – | – | 6.45 ± 0.06b |
Mean ± Standard deviation
Different letter means significant difference (P < 0.05) among fertilization treatments.
The statistical analyses (general linear regression) of 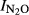 showed statistically significant differences between sites (P < 0.01) and significant effects of pH (P < 0.001) as well as a significant interaction between pH and site (P < 0.01). Fig.6 shows the N2O production index (
showed statistically significant differences between sites (P < 0.01) and significant effects of pH (P < 0.001) as well as a significant interaction between pH and site (P < 0.01). Fig.6 shows the N2O production index (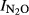 Phase I and Phase II) for all the treatments, plotted against the soil pH. For the field experiments where treatments affected the soil pH, we found that
Phase I and Phase II) for all the treatments, plotted against the soil pH. For the field experiments where treatments affected the soil pH, we found that 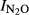 in Phase I was significantly correlated with pH. For the soils from QY, the estimated regression for
in Phase I was significantly correlated with pH. For the soils from QY, the estimated regression for 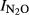 as a function of soil pH was
as a function of soil pH was 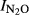 = 1.52–0.15*pH, (r2 = 0.607, P = 0.008). For the GZL soils the function was
= 1.52–0.15*pH, (r2 = 0.607, P = 0.008). For the GZL soils the function was 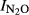 = 2.81–0.32*pH, (r2 = 0.764, P = 0.001); and for the SG soils the function was
= 2.81–0.32*pH, (r2 = 0.764, P = 0.001); and for the SG soils the function was 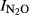 = 4.16–0.55*pH, (r2 = 0.715, P = 0.001). When including samples from all five sites, we estimated a general pH model (
= 4.16–0.55*pH, (r2 = 0.715, P = 0.001). When including samples from all five sites, we estimated a general pH model (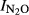 -Phase I vs. pH):
-Phase I vs. pH): 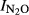 = 1.64–0.18*pH, (r2 = 0.759, P < 0.001). In Phase II, the most acid soil (QY) retained a high
= 1.64–0.18*pH, (r2 = 0.759, P < 0.001). In Phase II, the most acid soil (QY) retained a high 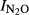 , which was correlated with pH [
, which was correlated with pH [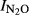 = 1.45–0.19*pH, (r2 = 0.49, P = 0.024)]. In the more alkaline soils, however, very low
= 1.45–0.19*pH, (r2 = 0.49, P = 0.024)]. In the more alkaline soils, however, very low 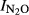 were recorded for Phase II, apparently independent of the pH. The similar relationships can be also found with indexes (
were recorded for Phase II, apparently independent of the pH. The similar relationships can be also found with indexes (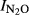 ) calculated for T Eqn (1) = the time when 50 nmol NO3-N g−1 soil and 200 nmol NO3-N g−1 soil had been recovered as (NO+N2O+N2)-N in either Phase I or Phase II (Fig. S2–S4).
) calculated for T Eqn (1) = the time when 50 nmol NO3-N g−1 soil and 200 nmol NO3-N g−1 soil had been recovered as (NO+N2O+N2)-N in either Phase I or Phase II (Fig. S2–S4).
Figure 6.
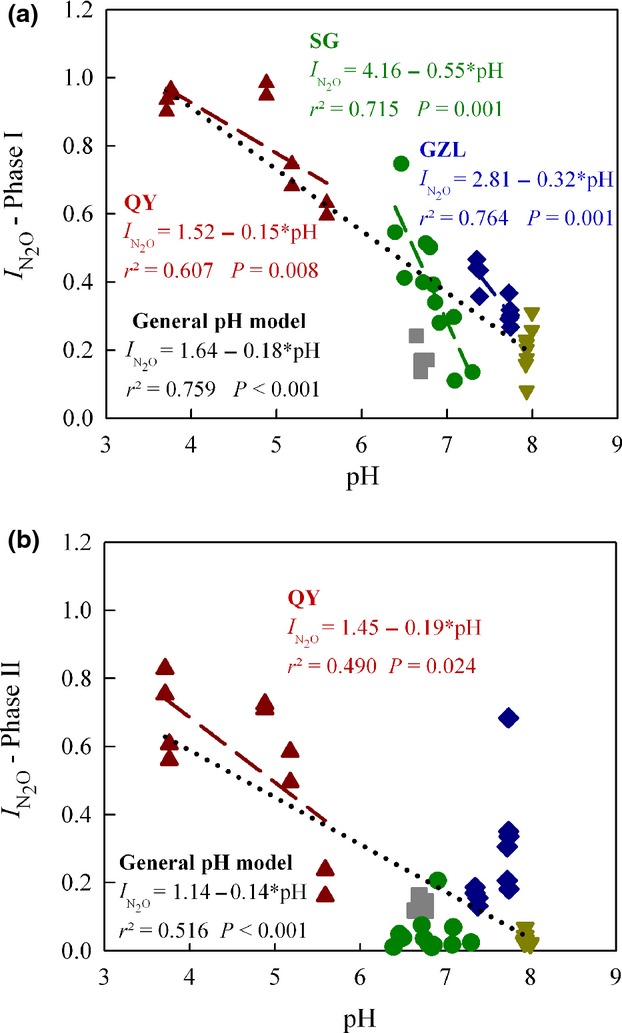
Relationship between soil pH and N2O production index (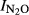 ) in Phase I (a) and Phase II (b). Data for single vials with soil from Gongzhuling (GZL, ), Shangzhuang (SZ, ), Qiyang (QY, ), Changwu (CW, ), and Shouguang (SG, ) are shown. Linear regressions for individual sites are shown, as well as the general pH regression model (including all samples).
) in Phase I (a) and Phase II (b). Data for single vials with soil from Gongzhuling (GZL, ), Shangzhuang (SZ, ), Qiyang (QY, ), Changwu (CW, ), and Shouguang (SG, ) are shown. Linear regressions for individual sites are shown, as well as the general pH regression model (including all samples).
We also investigated if the N input had an effect on N2O production index (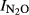 ), independent of pH. This was done by analyzing the residuals from the general pH model for phase I (
), independent of pH. This was done by analyzing the residuals from the general pH model for phase I (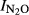 = 1.64–0.18*pH) against the nitrogen input (Fig.7). For the entire material, the residual was not significantly correlated with N input (r2 = 0.032), but in three sites (in GZL, SZ, and SG), there was a clear trend that the residual increased with increasing N input.
= 1.64–0.18*pH) against the nitrogen input (Fig.7). For the entire material, the residual was not significantly correlated with N input (r2 = 0.032), but in three sites (in GZL, SZ, and SG), there was a clear trend that the residual increased with increasing N input.
Figure 7.
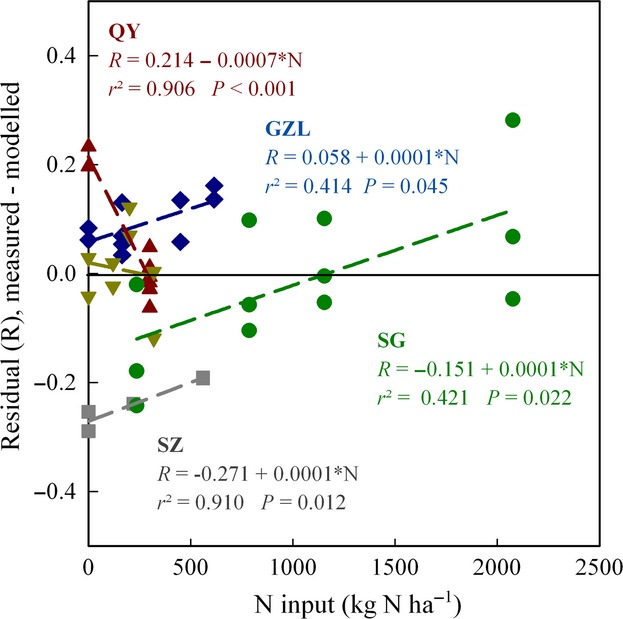
Residuals from the general regression if 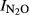 Phase I against pH (model:
Phase I against pH (model: 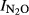 = 1.64–0.18*pH, see Fig6a), plotted against the annual input of total nitrogen (mineral N + organic N). Data for single vials with soil from Gongzhuling (GZL, ), Shangzhuang (SZ,), Qiyang (QY,), Changwu (CW,), and Shouguang (SG,) are shown. Trends are shown for individual sites.
= 1.64–0.18*pH, see Fig6a), plotted against the annual input of total nitrogen (mineral N + organic N). Data for single vials with soil from Gongzhuling (GZL, ), Shangzhuang (SZ,), Qiyang (QY,), Changwu (CW,), and Shouguang (SG,) are shown. Trends are shown for individual sites.
Fertilizer effects on NO accumulation
As shown in Fig.3, NO accumulated early during anoxic incubation (both in Phase I and II), but the maximum concentrations reached were much lower than that of N2O – notice that scales are different for the two gases. The maximum NO concentrations reached in the different treatments are shown in Fig. S5 for Phase I and Fig. S6 for Phase II. The values varied from 1–25 nmol NO-N g−1 dw soil and 2–52 nmol NO-N g−1 dw soil (1 nmol NO-N g−1 = 13.5 nM NO in liquid) for all soils in Phase I and Phase II, respectively, except for the most acidic soil with chemical nitrogen fertilizer input in the Qiyang experiment (QY, Nopt, pH 3.75; see Table3) which accumulated 178 nmol NO-N g−1 (2400 nM, in the liquid) in Phase I. We also plotted the maximum NO concentrations with pH for all soils (Fig. S7), but found no consistent relationship.
Liming experiment
The NO, N2O, and N2 gas kinetics of soils with and without liming were compared for some of the Chinese soils, as well as soils from a long-term liming experiment on peat soil in Norway [Fjaler, Liu et al. (2010)]. Fig.8 shows single vial results for one of the soils (Fjaler pH4 soil). For the original soil with low pH (unlimed, without glutamate, pH 3.60, shown in Fig.8a), NO was produced immediately when anaerobic incubation started. The measured concentration of NO peaked at 18 h, followed by a slow reduction toward a stable level throughout the incubation. The amounts of N2O (measured concentration) and N2 (accumulated net production) both increased linearly but relatively slowly throughout the 150 h incubation. In the same soil with CaCO3 added to raise the pH (limed, without glutamate, pH 6.71, shown in Fig.8b), the gas kinetics was profoundly different. For the first 35 h, the productions of the three N-gases were similar to that in the unlimed soil. After that the liming clearly induced higher denitrification activity and a sharp increase in the rate of N2O reduction to N2, although this lagged behind the onset of rapid N2O production. As a result of this, the limed soil accumulated more N2O than the unlimed soil for a period lasting from 50 to 90 h after the start of the anoxic incubation.
Figure 8.
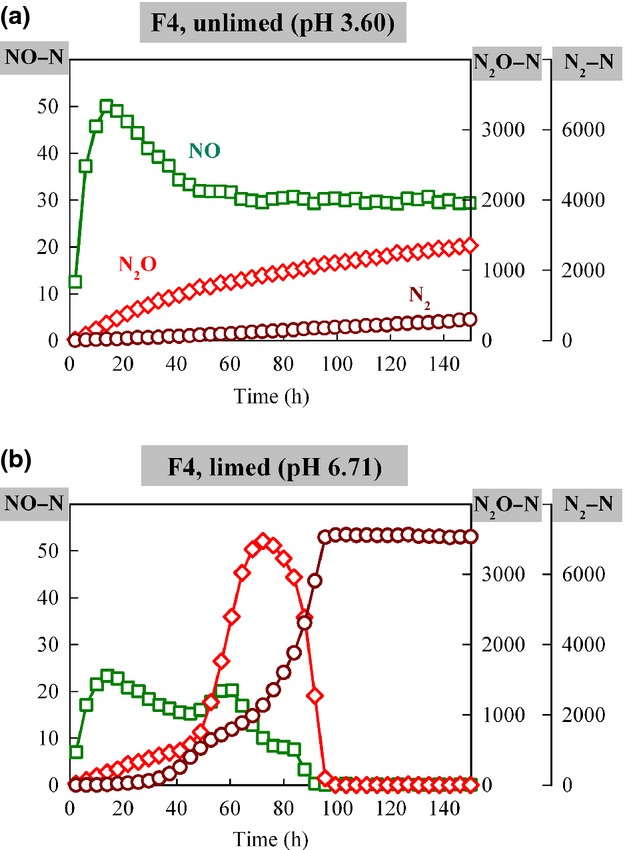
Demonstration of liming effect on NO, N2O, and N2 gas kinetic during anoxic incubation. Panel a shows the gas kinetics of the original soil (Peat soil, F4, unlimed, without glutamate, pH 3.60), panel b shows the gas kinetics of the same soil after liming (pH 6.71). NO and N2O are shown as measured (nmol g−1 soil dry weight), whereas N2 is shown as cumulative production (nmol g−1 soil dry weight).
The results for all soils tested are summarized in Fig.9 which plots the N2O production index (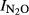 ) against pH, in pairwise comparisons of limed and unlimed soils. The raw data behind this are shown in (Fig. S8–S11), plotted in the same way as in Fig.8; the differences in final N2 concentration (nmol g−1 soil) between peat soils (F4 and F6) and mineral soils (QY and SG) are due to different amounts of water (containing 2 mm NO3−) per g soil [see Table S2)]. CaCO3 induced increasing pH in all soils, most pronounced in the naturally low pH soils. Along with the increase of the soil pH, the N2O production index dropped for all soils, both for treatments with and without glutamate. We also plotted the index for other values of T Eqn (1), and similar responses to liming were observed (results can be found in Fig. S12–S17).
) against pH, in pairwise comparisons of limed and unlimed soils. The raw data behind this are shown in (Fig. S8–S11), plotted in the same way as in Fig.8; the differences in final N2 concentration (nmol g−1 soil) between peat soils (F4 and F6) and mineral soils (QY and SG) are due to different amounts of water (containing 2 mm NO3−) per g soil [see Table S2)]. CaCO3 induced increasing pH in all soils, most pronounced in the naturally low pH soils. Along with the increase of the soil pH, the N2O production index dropped for all soils, both for treatments with and without glutamate. We also plotted the index for other values of T Eqn (1), and similar responses to liming were observed (results can be found in Fig. S12–S17).
Figure 9.
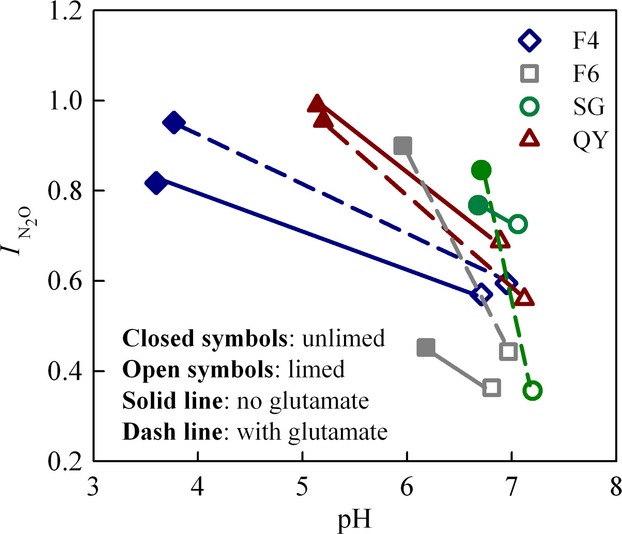
Relationship between soil pH and N2O production index in the liming experiment. The four soils limed were peat soil from Fjaler (Norway) with pH = 4 (F4), peat soil from Fjaler with pH with pH = 6 (F6), the Shouguang soil (SG) and the Qiyang soil (QY). The lines connect the results for limed (open symbols) and unlimed (closed symbols) for each soil and treatment (solid lines: no glutamate added, dashed lines: glutamate added). For the mineral soils (QY and SG), the indexes (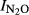 ) were calculated for the time period from 0 to T (Eqn 1), where T is the time when 100 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)−N. For the peat soils (F4 and F6), the indexes (
) were calculated for the time period from 0 to T (Eqn 1), where T is the time when 100 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)−N. For the peat soils (F4 and F6), the indexes (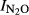 ) were calculated for T = the time when 3000 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N.
) were calculated for T = the time when 3000 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N.
Discussion
A recent survey of Chinese farmers' fertilizer practice showed a widespread excessive N use: grain crops receive 220–270 kg N ha−1, but remove only 120–160 kg N ha−1; fruits and vegetables receive 400–600 kg N ha−1 but remove only 83–130 kg N ha−1 (Zhang et al., 2013). Guo et al. (2010) concluded that these high nitrogen inputs have already acidified Chinese croplands significantly, except for high pH soils. We found a significant nitrogen-induced acidification in two of the field experiments that had run for more than 20 years, whereas the third long-term field experiment (CW, original pH > 7) showed no sign of fertilizer-induced acidification. The two sites that had run for less than 20 years (SZ and SG) showed lowered pH values in response to high N input, but this was statistically significant for only one treatment. The effect of manure on soil pH appears to differ between the field experiments – acidification in GZL, alkalinization in QY. This may reflect contrasting manure quality (see Table1). Thus, our results corroborate the findings of the nationwide study by Guo et al. (2010): high nitrogen fertilizer levels leads to a gradual acidification of Chinese cropland soils except for soils with high pH.
Our second hypothesis was that this acidification would lead to a higher propensity of the soils to emit N2O. To assess this, we should take both potential denitrification rate and denitrification product ratio N2O/(N2O+N2) into account. The potential denitrification rate was strongly correlated with the oxic respiration rate of the soils, as demonstrated by plotting the electron flow rate to denitrification during anoxic incubation (De−, Phase I) against the electron flow rate to oxygen during the preceding oxic incubation (Re−, Phase Oxic) (Fig.4). Manure inputs clearly increased the Re− and induced a similar relative increase in De−. Thus, the potential denitrification in the soils largely reflect the available carbon substrates in the soil for heterotrophic microbial activity, which depend on the long-term history of the soil as well as the recent history of organic carbon inputs. The De−/Re− ratio can possibly be used as a proxy for the percentage of respiring organisms that were able to switch to anoxic respiration. One would expect the De−/Re− ratio to be strongly correlation with soil pH, based on the common notion that denitrifying organisms are favored by high pH (Čuhel & Šimek, 2011). We do find a correlation when including all soil samples, but within the individual experiments there is little evidence for a strong pH control of the De−/Re− ratio (Fig.5). In conclusion, potential denitrification rates in the soils appear to be controlled largely by the same factors that control their oxic respiration rates; the annual input of organic material is evidently of great importance here.
To compare the ‘intrinsic’ N2O/(N2O+N2) product ratios of the soils, we used the N2O production index (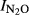 ) rather than the product ratio at any specific time during the anoxic phase. The index expresses the fraction of denitrified N which is prone to be emitted if the denitrification occurred in an anoxic microenvironment in the soil (in situ). The plots of
) rather than the product ratio at any specific time during the anoxic phase. The index expresses the fraction of denitrified N which is prone to be emitted if the denitrification occurred in an anoxic microenvironment in the soil (in situ). The plots of 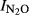 against pH (Fig.6) show a strong overall correlation with soil pH, and the estimated regression function (
against pH (Fig.6) show a strong overall correlation with soil pH, and the estimated regression function (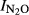 = 1.64–0.18*pH, r2 = 0.759) was strikingly similar to that determined by Liu et al. (2010) for soils taken from a long-term liming experiment in organic soil in Norway; regression function based on a combination of all data for their first anoxic incubation period is
= 1.64–0.18*pH, r2 = 0.759) was strikingly similar to that determined by Liu et al. (2010) for soils taken from a long-term liming experiment in organic soil in Norway; regression function based on a combination of all data for their first anoxic incubation period is 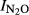 = 1.6–0.15*pH (r2 = 0.62, n = 29). The results are also in good agreement with the observations by Raut et al. (2012), who documented that the ongoing intensification of Nepalese agriculture acidifies the soils and increases the N2O/(N2O+N2) product ratios of denitrification. The regression model for
= 1.6–0.15*pH (r2 = 0.62, n = 29). The results are also in good agreement with the observations by Raut et al. (2012), who documented that the ongoing intensification of Nepalese agriculture acidifies the soils and increases the N2O/(N2O+N2) product ratios of denitrification. The regression model for 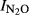 based on their data for Nepalese upland soils is:
based on their data for Nepalese upland soils is: 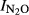 = 1.82–0.22*pH (r2 = 0.33, n = 32).
= 1.82–0.22*pH (r2 = 0.33, n = 32).
With one notable exception (Shangzhuang, SZ), the results for the individual sites all fit reasonably well with the general regression function, and we find strong evidence for a pH effect within those sites where acidification had taken place (QY, SG and GZL). For the sites where pH was not significantly affected by pH (SZ and CW), 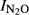 all fell within a narrow range, apparently unaffected by fertilization practice.
all fell within a narrow range, apparently unaffected by fertilization practice.
In theory, the nitrogen level could have a direct effect on 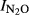 , independent of soil pH. The reason for this is that extremely high nitrogen inputs, such as that in Shouguang (2000 kg N ha−1 yr−1) would provide an environment where denitrifying organisms would hardly ever experience shortage of nitrate or nitrite during anoxic spells. This could favor organisms which do not express N2O reductase, because the expression of N2O reductase would be a waste of energy in situations with ample amounts of nitrate and nitrite. We attempted to shed some light on this by exploring the residuals from the general regression of
, independent of soil pH. The reason for this is that extremely high nitrogen inputs, such as that in Shouguang (2000 kg N ha−1 yr−1) would provide an environment where denitrifying organisms would hardly ever experience shortage of nitrate or nitrite during anoxic spells. This could favor organisms which do not express N2O reductase, because the expression of N2O reductase would be a waste of energy in situations with ample amounts of nitrate and nitrite. We attempted to shed some light on this by exploring the residuals from the general regression of 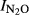 against soil pH. When plotting these residuals against the annual input of nitrogen (Fig.7), we do indeed see a positive trend for three of the experimental sites. This result is not conclusive, however, because N-level and pH are correlated, and the trends demonstrated may reflect the fact that the ‘true’ response of
against soil pH. When plotting these residuals against the annual input of nitrogen (Fig.7), we do indeed see a positive trend for three of the experimental sites. This result is not conclusive, however, because N-level and pH are correlated, and the trends demonstrated may reflect the fact that the ‘true’ response of 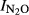 to soil pH is not necessarily linear over the entire pH range investigated.
to soil pH is not necessarily linear over the entire pH range investigated.
We included measurements of NO in this study because it is a toxic intermediate in denitrifiation, and a strong inducer of some denitrification genes (van Spanning et al., 2007). NO emissions from soil is of environmental concern because NO interferes with troposphere chemistry by scavenging hydroxyl radicals (Crutzen & Zimmermann, 1991).The results showed that NO concentrations could reach micromolar concentrations in the most acidic soil (QY), but for all the other soils the maximum NO concentrations reached were 1–52 nmol N g−1 soil (13–700 nM, in the liquid), which is comparable to the characteristic concentration for balanced expression of denitrification in model organisms with homeostatic control of NO during denitrification (Bergaust et al., 2011).
In our studies, the soils were incubated in the closed vials. The N2O production index can only indicate the inherent propensity of soils to emit N2O from denitrification as controlled at the cellular level. Under field conditions, the fraction of N2O escaping to the atmosphere will depend on the diffusion constraints, which is a function of texture and air-filled porosity. This implies that field emissions across landscapes with variable soil texture and drainage are not necessarily positively correlated with acidity (Gu et al., 2013). Ongoing field experiments have shown, however, that within single fields which are homogenous with respect to texture and drainage, the spatial variation of N2O emissions is strongly correlated with the pH and the N2O production index (Lundon, Dörsch and Bakken, manuscripts in preparation).
In conclusion, our results provide strong evidence that the soil acidification driven by high fertilizer levels in China affects the N2O/(N2+N2O) product ratio of denitrification, and hence the propensity of the soils to emit N2O.
What are the mechanisms behind the strong effect of acidification on the product ratio? The most plausible explanation is a direct effect of low pH on the organisms' ability to produce a functional N2O reductase. The first documentation of this was provided through a study of gene transcription and denitrification kinetics in the model organism Paracoccus denitrificans (Bergaust et al., 2010). It was demonstrated that this organism was unable to produce functional N2O reductases at pH ≤ 6.1, and the most plausible explanation was interference with the assembly of the protein in the periplasm. They further demonstrated that the N2O reductase successfully assembled at pH 7 was fully functional at lower pH levels. Subsequently, Liu et al. (2010) provided circumstantial evidence for a similar effect of low pH in soils: soils from a long-term liming experiment showed a pervasive control of soil pH on the N2O/(N2O+N2) product ratio, although the relative amount of nosZ genes and their transcripts appeared unaffected by pH. More stringent evidence was provided recently through experiments with cells extracted from soils by density gradient centrifugation. The extracted cells (from soils with pH ranging from 4 to 8) were incubated at pH ranging from 5.7 to 7.6, while monitoring NO, N2O, and N2 gas kinetics and gene transcription under anoxic conditions. The results showed that none of the cells were able to express functional N2OR at pH ≤ 6.1, despite significant transcription of the nosZ gene; but the N2O reductase synthesized at pH 7 was fully functional at lower pH levels (Liu, Frostegård and Bakken, manuscript in preparation).
An alternative explanation to the pH-control of the N2O/(N2O+N2) product ratio is that soil pH controls the composition of the denitrifying community – ‘distal control’ as defined by Čuhel & Šimek (2011). However, if a community shift was the explanation to the high N2O/(N2O+N2) product ratio in the acidified soils, liming is unlikely to affect the product ratio immediately. Our liming experiments with acidified Chinese soils showed a dramatic reduction in the N2O product ratio after 2 days of oxic preincubation, and equivalent results were observed for the two soil samples from the Norwegian long-term liming experiments (Fig.9). Similar results were obtained by Čuhel & Šimek (2011) who investigated the effect of liming on the N2O/(N2O+N2) product ratio of denitrification in pasture soils with a long history of contrasting pH management; adequate N2O reductase activity was restored immediately by increasing the soil pH. Van den Heuvel et al. (2011) did a similar investigation of pH adjustments of riparian soils. The soils were incubated anoxically as slurries adjusted to a pH range from 4 to 7, and they found a negative exponential relationship between pH and the N2O/N2 product ratio. However, if we transform their data to N2O/(N2O+N2) product ratios, we find a linear relationship between this ratio and the pH for the pH range 4–6.
The immediate result of liming, as demonstrated in Fig.8, is not necessarily to reduce the N2O emission; after 80 h of anoxic incubation, the N2O concentration was much higher in the limed than in the unlimed soil, despite the fact that the N2O/(N2O+N2) ratio at this time was clearly lowered by the liming. The reason is that liming induced a much higher denitrification rate, most likely by inducing higher metabolic activity through enhancing the availability of soil organic carbon (Bååth & Arnebrant, 1994). Similarly, Baggs et al. (2010) found that the immediate effect of liming was to stimulate nitrification and denitrification, and the N2O emission from both, as differentiated by tracer technique. Such immediate dramatic enhancement of the metabolic activity is likely to be transient, however (Kemmitt et al., 2006).
We scanned previous field experiments and laboratory experiments with oxic incubation of soils and found variable effects of liming on measured emission of N2O: The effect of liming pastoral soils in New Zealand was investigated in a series of field- and laboratory experiments (Zaman et al., 2007, 2008; Zaman & Nguyen, 2010), demonstrating that the immediate effect of liming was increased N2 emissions (estimated by acetylene inhibition), lowered N2O/N2 emission ratios, but practically unaltered emission of N2O. Clough et al. (2004) measured N2 and N2O emissions from microcosms (oxic incubations) for 80 days after adjusting the soil pH to a range of pH levels (4.7–7.2), and found that liming lowered the N2O/N2 emission ratio and the net emission of N2O from soil at low soil moisture content but not from water saturated soil. Khan et al. (2011) measured N2O emissions induced by bovine urine 2 weeks after manipulating the pH by liming, and found that the accumulated emissions increased with liming. Goodroad et al. (1984) measured N2O emissions in field experiments with long-term contrasting pH management (4.7–6.7). They found no statistically significant effect of pH due to large spatial and temporal variation in N2O emission (CV 50–200%). Similarly, Galbally et al. (2010) measured N2O emissions in liming experiments 2 years after the lime had been applied. The average daily emission was 0.96 mg N m−1 d−1 in the acid (unlimed) plots (pH 4) and 0.88 in the limed plots, but the difference was not statistically significant. The authors concluded, however, that the liming increased plant productivity by 25%, thus the emission per unit of plant products was lowered by liming. Borken & Brumme (1997) measured N2O emissions from various forest soils that had been limed >10 years prior to the measurements. They observed that liming reduced the annual emissions by 9–62% compared with unlimed plots. Mkhabela et al. (2006) studied the immediate effect of liming an acid loam soil (pH 4.71) to reach pH 6.3 and 7.4, by oxic incubations of soil (amended with manure) for a period of 21 days. The accumulated N2O emissions (expressed as g N2O-N ha−1) were 547, 46 and 134 g for unlimed, moderately limed (pH 6.3) and strongly limed (pH 7.4) respectively. The effects were statistically significant.
On this background, it is hardly surprising that management of soil pH is not on IPCC's list of mitigation options for reducing N2O emissions (IPCC, 2007). But the observations cited above cannot be taken as a ‘proof of absence’, i.e. that pH-management will not have a consistent effect on N2O emissions on large temporal and spatial scale. The few studies where both N2 and N2O emissions have been measured all show a lowered N2O/(N2O+N2) emission ratio with increasing soil pH. Thus, they corroborate our findings based on anoxic incubations of soils; the N2O/(N2O+N2) product ratio of denitrification is pervasively controlled by soil pH. It appears likely, therefore that the overall effect of soil acidification will be higher N2O emissions, and this can be counteracted by increasing the pH. It may take some effort to test this rigorously through field experiments, because N2O emissions in field experiments are notoriously variable in both time and space. But it would probably be worth the effort, considering the grave implications for future N2O emissions from developing countries such as China.
Acknowledgments
We would like to thank Lars Molstad from the Norwegian UMB-Nitrogen-group (http://www.umb.no/nitrogengroup/) for designing and programming of our robotized incubation system for analyzing gas kinetics. Thanks also to XU Minggang, Institute of Agricultural Resources and Zonation, Chinese Academy of Agricultural Sciences, for collecting soil samples from Gongzhuling and Qiyang. This research was part of the project ‘Fertilizer Technologies for Reduced N2O Emission from Food Production’, which is a collaborative project involving Yara International, Norway and China, and financed by the Norwegian Research council. One of the authors (Z. Qu) is grateful to the China Scholarship Council (CSC) for the financial support as a recipient of CSC scholarship.
Supporting Information
Additional Supporting Information may be found in the online version of this article
Relationship of the oxic respiration rate (ROX) and soil organic carbon (SOC).
Figure S2. Relationship between soil pH and N2O production index (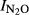 ) in Phase I calculated with T Eqn (1) = the time when 50 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N in the soil comparison experiment.
) in Phase I calculated with T Eqn (1) = the time when 50 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N in the soil comparison experiment.
Figure S3. Relationship between soil pH and N2O production index (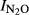 ) in Phase I calculated with T Eqn (1) = the time when 200 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N in the soil comparison experiment.
) in Phase I calculated with T Eqn (1) = the time when 200 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N in the soil comparison experiment.
Figure S4. Relationship between soil pH and N2O production index (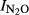 ) in Phase II calculated with T Eqn (1) = the time when 200 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N in the soil comparison experiment.
) in Phase II calculated with T Eqn (1) = the time when 200 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N in the soil comparison experiment.
Figure S5. Effects of long-term fertilization on maximum NO concentration (nmol N g−1 soil) in Phase I for each site.
Figure S6. Effects of long-term fertilization on maximum NO concentration (nmol N g−1 soil) in Phase II for each site.
Figure S7. Relationship between soil pH and maximum NO concentration (nmol N g−1 soil) in both Phase I (a) and Phase II (b).
Figure S8. NO, N2O, and N2 gas kinetics during anoxic incubation for Fjaler pH4 soil (F4) in the liming experiment.
Figure S9. NO, N2O, and N2 gas kinetics during anoxic incubation for Fjaler pH6 soil (F6) in the liming experiment.
Figure S10. NO, N2O, and N2 gas kinetics during anoxic incubation for Qiyang soil (QY) in the liming experiment.
Figure S11. NO, N2O, and N2 gas kinetics during anoxic incubation for Shouguang soil (SG) in the liming experiment.
Figure S12. Relationship between soil pH and N2O production index in the liming experiment for mineral soils (QY and SG). Indexes (IN2O) were calculated at T Eqn (1), with T Eqn (1) = the time when 50 nmol NO3-N g-1 soil had been recovered as (NO+N2O+N2)-N
Figure S13. Relationship between soil pH and N2O production index in the liming experiment for mineral soils (QY and SG). Indexes (IN2O) were calculated at T Eqn (1), with T Eqn (1) = the time when 200 nmol NO3-N g-1 soil had been recovered as (NO+N2O+N2)-N
Figure S14. Relationship between soil pH and N2O production index in the liming experiment for mineral soils (QY and SG). Indexes (IN2O) were calculated at T Eqn (1), with T Eqn (1) = the time when 300 nmol NO3-N g-1 soil had been recovered as (NO+N2O+N2)-N
Figure S15. Relationship between soil pH and N2O production index in the liming experiment for peat soils (F4 and F6). Indexes (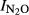 ) were calculated at T Eqn (1), with T Eqn (1) = the time when 1000 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N.
) were calculated at T Eqn (1), with T Eqn (1) = the time when 1000 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N.
Figure S16. Relationship between soil pH and N2O production index in the liming experiment for peat soils (F4 and F6). Indexes (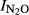 ) were calculated at T Eqn (1), with T Eqn (1) = the time when 2000 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N.
) were calculated at T Eqn (1), with T Eqn (1) = the time when 2000 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N.
Figure S17. Relationship between soil pH and N2O production index in the liming experiment for peat soils (F4 and F6). Indexes (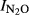 ) were calculated at T Eqn (1), with T Eqn (1) = the time when 4000 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N.
) were calculated at T Eqn (1), with T Eqn (1) = the time when 4000 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N.
Table S1. The experimental treatments for each field experiment.
Table S2. Water content of soils (based on soil fresh weight) in the liming experiment.
References
- Bååth E, Arnebrant K. Growth rate and response of bacterial communities to pH in limed and ash treated forest soils. Soil Biology and Biochemistry. 1994;26:995–1001. [Google Scholar]
- Baggs EM, Smales CL, Bateman EJ. Changing pH shifts the microbial sourceas well as the magnitude of N2O emission from soil. Biology and Fertility of Soils. 2010;46:793–805. [Google Scholar]
- Bakken LR, Bergaust L, Liu B, Frostegård Å. Regulation of denitrification at the cellular level: a clue to the understanding of N2O emissions from soils. Philosophical Transactions of the Royal Society B: Biological Sciences. 2012;367:1226–1234. doi: 10.1098/rstb.2011.0321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergaust L, Mao Y, Bakken LR, Frostegård Å. Denitrification response patterns during the transition to anoxic respiration and posttranscriptional effects of suboptimal pH on nitrogen oxide reductase in Paracoccus denitrificans. Applied and environmental microbiology. 2010;76:6387–6396. doi: 10.1128/AEM.00608-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergaust L, Bakken L, Frostegård Å. Denitrification regulatory phenotype, a new term for the characterization of denitrifying bacteria. Biochemical Society Transactions. 2011;39:207–212. doi: 10.1042/BST0390207. [DOI] [PubMed] [Google Scholar]
- Borken W, Brumme R. Liming practice in temperate forest ecosystems and the effects on CO2, N2O and CH4 fluxes. Soil Use and Management. 1997;13:251–257. [Google Scholar]
- Clough TJ, Kelliher FM, Sherlock RR, Ford CD. Lime and soil moisture effects on nitrous oxide emissions from a urine patch. Soil Science Society of America Journal. 2004;68:1600–1609. [Google Scholar]
- Crutzen PJ, Zimmermann PH. The changing photochemistry of the troposphere. Tellus B. 1991;43:136–151. [Google Scholar]
- Čuhel J, Šimek M. Proximal and distal control by pH of denitrification rate in a pasture soil. Agriculture, Ecosystems & Environment. 2011;141:230–233. [Google Scholar]
- Cui S, Shi Y, Groffman PM, Schlesinger WH, Zhu YG. Centennial-scale analysis of the creation and fate of reactive nitrogen in China (1910-2010) Proceedings of the National Academy of Sciences of the United States of America. 2013;110:2053–2057. doi: 10.1073/pnas.1221638110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan T, Stewart B, Yong W, Junjie L, Guangye Z. Long-term fertilization effects on grain yield, water-use efficiency and soil fertility in the dryland of Loess Plateau in China. Agriculture, Ecosystems & Environment. 2005;106:313–329. [Google Scholar]
- FAO. World Soil Resources Report 60. FAO, Rome, Italy: 1988. Soil Map of the World.Revised Legend. Reprinted with corrections. [Google Scholar]
- Galbally IE, Meyer MC, Wang Y-P, Smith CJ, Weeks IA. Nitrous oxide emissions from a legume pasture and the influences of liming and urine addition. Agriculture, Ecosystems & Environment. 2010;136:262–272. [Google Scholar]
- Goodroad L, Keeney D, Peterson L. Nitrous oxide emissions from agricultural soils in Wisconsin. Journal of Environmental Quality. 1984;13:557–561. [Google Scholar]
- Gu J, Nicoullaud B, Rochette P, Grossel A, Henault C, Cellier P, Richard G. A regional experiment suggest that soil texture is a major control of N2O emissions from tile drained winter wheat fields during the firtilization period. Soil Biology and Biochemistry. 2013;60:131–141. [Google Scholar]
- Guo J, Liu X, Zhang Y, et al. Significant acidification in major Chinese croplands. Science. 2010;327:1008–1010. doi: 10.1126/science.1182570. [DOI] [PubMed] [Google Scholar]
- Hao MD, Fan J, Wang QJ, Dang TH, Guo SL, Wang JJ. Wheat grain yield and yield stability in a long-term fertilization experiment on the Loess Plateau. Pedosphere. 2007;17:257–264. [Google Scholar]
- He F, Jiang R, Chen Q, Zhang F, Su F. Nitrous oxide emissions from an intensively managed greenhouse vegetable cropping system in Northern China. Environmental Pollution. 2009;157:1666–1672. doi: 10.1016/j.envpol.2008.12.017. [DOI] [PubMed] [Google Scholar]
- IPCC. 2006 IPCC Guidelines for National Greenhouse Gas Inventories, Prepared by the National Greenhouse Gas Inventories Programme. Volume 4: Agriculture, Forestry and Other Land Use. Japan: IGES; 2006. Chapter 11: N2O emissions from managed soils, and Co2 emissions from lime and urea application; pp. 11.1–11.54. Eggleston HS, Buendia L, Miwa K, Ngara T, Tanabe K, [Google Scholar]
- IPCC. Climate Change 2007: Mitigation. Contribution of Working Group III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press; 2007. Summary for Policymakers; pp. 1–23. Metz B, Davidson OR, Bosch PR, Dave R, Meyer LA, [Google Scholar]
- Jiang D, Hengsdijk H, Dai TB, De Boer W, Jing Q, CAO WX. Long-term effects of manure and inorganic fertilizers on yield and soil fertility for a winter wheat-maize system in Jiangsu, China. Pedosphere. 2006;16:25–32. [Google Scholar]
- Ju X, Liu X, Zhang F, Roelcke M. Nitrogen fertilization, soil nitrate accumulation, and policy recommendations in several agricultural regions of China. Ambio: A Journal of the Human Environment. 2004;33:300–305. doi: 10.1579/0044-7447-33.6.300. [DOI] [PubMed] [Google Scholar]
- Kemmitt SJ, Wright D, Goulding KW, Jones DL. pH regulation of carbon and nitrogen dynamics in two agricultural soils. Soil Biology and Biochemistry. 2006;38:898–911. [Google Scholar]
- Khan S, Clough T, Goh K, Sherlock R. Influence of soil pH on NOx and N2O emissions from bovine urine applied to soil columns. New Zealand Journal of Agricultural Research. 2011;54:285–301. [Google Scholar]
- Liu B, Mørkved PT, Frostegård Å, Bakken LR. Denitrification gene pools, transcription and kinetics of NO, N2O and N2 production as affected by soil pH. FEMS Microbiology Ecology. 2010;72:407–417. doi: 10.1111/j.1574-6941.2010.00856.x. [DOI] [PubMed] [Google Scholar]
- Mkhabela M, Gordon R, Burton D, Madani A, Hart W. Effect of lime, dicyandiamide and soil water content on ammonia and nitrous oxide emissions following application of liquid hog manure to a marshland soil. Plant and Soil. 2006;284:351–361. [Google Scholar]
- Molstad L, Dörsch P, Bakken LR. Robotized incubation system for monitoring gases (O2, NO, N2O N2) in denitrifying cultures. Journal of Microbiological Methods. 2007;71:202–211. doi: 10.1016/j.mimet.2007.08.011. [DOI] [PubMed] [Google Scholar]
- Mørkved PT, Dörsch P, Bakken LR. The N2O product ratio of nitrification and its dependence on long-term changes in soil pH. Soil Biology and Biochemistry. 2007;39:2048–2057. [Google Scholar]
- Nömmik H. Investigations on denitrification in soil. Acta Agriculturae Scandinavica. 1956;6:195–228. [Google Scholar]
- Qiu S, Ju X, Lu X, et al. Improved Nitrogen Management for an Intensive Winter Wheat/Summer Maize Double-cropping System. Soil Science Society of America Journal. 2012;76:286–297. [Google Scholar]
- Raut N, Dörsch P, Sitaula BK, Bakken LR. Soil acidification by intensified crop production in South Asia results in higher N2O/(N2 +N2O) product ratio of denitrification. Soil Biology and Biochemistry. 2012;55:104–112. [Google Scholar]
- Šimek M, Cooper J. The influence of soil pH on denitrification: progress towards the understanding of this interaction over the last 50 years. European Journal of Soil Science. 2002;53:345–354. [Google Scholar]
- van Spanning RJ, Richardson DJ, Ferguson SJ. Introduction to the biochemistry and molecular biology of denitrification. In: Bothe H, Ferguson S, Newton WE, editors. Biology of the Nitrogen Cycle. Amsterdam, The Netherlands: Elsevier; 2007. pp. 382–395. [Google Scholar]
- Van den Heuvel RN, Bakker SE, Jetten MSM, Hefting MM. Decreased N2O reduction by low soil pH causes high emissions in a riparian ecosystem. Geobiology. 2011;9:294–300. doi: 10.1111/j.1472-4669.2011.00276.x. [DOI] [PubMed] [Google Scholar]
- Wijler J, Delwiche C. Investigations on the denitrifying process in soil. Plant and Soil. 1954;5:155–169. [Google Scholar]
- Zaman M, Nguyen M. Effect of lime or zeolite on N2O and N2 emissions from a pastoral soil treated with urine or nitrate-N fertilizer under field conditions. Agriculture, Ecosystems & Environment. 2010;136:254–261. [Google Scholar]
- Zaman M, Nguyen M, Matheson F, Blennerhassett J, Quin B. Can soil amendments (zeolite or lime) shift the balance between nitrous oxide and dinitrogen emissions from pasture and wetland soils receiving urine or urea-N? Soil Research. 2007;45:543–553. [Google Scholar]
- Zaman M, Nguyen M, Saggar S. N2O and N2 emissions from pasture and wetland soils with and without amendments of nitrate, lime and zeolite under laboratory condition. Soil Research. 2008;46:526–534. [Google Scholar]
- Zhang H, Wang B, Xu M. Effects of inorganic fertilizer inputs on grain yields and soil properties in a long-term wheat-corn cropping system in South China. Communications in Soil Science and Plant Analysis. 2008;39:1583–1599. [Google Scholar]
- Zhang H, Xu M, Zhang F. Long-term effects of manure application on grain yield under different cropping systems and ecological conditions in China. The Journal of Agricultural Science. 2009;147:31–42. [Google Scholar]
- Zhang WF, Dou ZX, He P, et al. New technologies reduce greenhouse gas emissions from nitrogenous fertilizer in China. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:8375–8380. doi: 10.1073/pnas.1210447110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao RF, Chen XP, Zhang FS, Zhang H, Schroder J, Römheld V. Fertilization and nitrogen balance in a wheat-maize rotation system in North China. Agronomy Journal. 2006;98:938–945. [Google Scholar]
- Zhu P, Ren J, Wang L, Zhang X, Yang X, MacTavish D. Long-term fertilization impacts on corn yields and soil organic matter on a clay-loam soil in Northeast China. Journal of Plant Nutrition and Soil Science. 2007;170:219–223. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Relationship of the oxic respiration rate (ROX) and soil organic carbon (SOC).
Figure S2. Relationship between soil pH and N2O production index () in Phase I calculated with T Eqn (1) = the time when 50 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N in the soil comparison experiment.
Figure S3. Relationship between soil pH and N2O production index () in Phase I calculated with T Eqn (1) = the time when 200 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N in the soil comparison experiment.
Figure S4. Relationship between soil pH and N2O production index () in Phase II calculated with T Eqn (1) = the time when 200 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N in the soil comparison experiment.
Figure S5. Effects of long-term fertilization on maximum NO concentration (nmol N g−1 soil) in Phase I for each site.
Figure S6. Effects of long-term fertilization on maximum NO concentration (nmol N g−1 soil) in Phase II for each site.
Figure S7. Relationship between soil pH and maximum NO concentration (nmol N g−1 soil) in both Phase I (a) and Phase II (b).
Figure S8. NO, N2O, and N2 gas kinetics during anoxic incubation for Fjaler pH4 soil (F4) in the liming experiment.
Figure S9. NO, N2O, and N2 gas kinetics during anoxic incubation for Fjaler pH6 soil (F6) in the liming experiment.
Figure S10. NO, N2O, and N2 gas kinetics during anoxic incubation for Qiyang soil (QY) in the liming experiment.
Figure S11. NO, N2O, and N2 gas kinetics during anoxic incubation for Shouguang soil (SG) in the liming experiment.
Figure S12. Relationship between soil pH and N2O production index in the liming experiment for mineral soils (QY and SG). Indexes (IN2O) were calculated at T Eqn (1), with T Eqn (1) = the time when 50 nmol NO3-N g-1 soil had been recovered as (NO+N2O+N2)-N
Figure S13. Relationship between soil pH and N2O production index in the liming experiment for mineral soils (QY and SG). Indexes (IN2O) were calculated at T Eqn (1), with T Eqn (1) = the time when 200 nmol NO3-N g-1 soil had been recovered as (NO+N2O+N2)-N
Figure S14. Relationship between soil pH and N2O production index in the liming experiment for mineral soils (QY and SG). Indexes (IN2O) were calculated at T Eqn (1), with T Eqn (1) = the time when 300 nmol NO3-N g-1 soil had been recovered as (NO+N2O+N2)-N
Figure S15. Relationship between soil pH and N2O production index in the liming experiment for peat soils (F4 and F6). Indexes () were calculated at T Eqn (1), with T Eqn (1) = the time when 1000 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N.
Figure S16. Relationship between soil pH and N2O production index in the liming experiment for peat soils (F4 and F6). Indexes () were calculated at T Eqn (1), with T Eqn (1) = the time when 2000 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N.
Figure S17. Relationship between soil pH and N2O production index in the liming experiment for peat soils (F4 and F6). Indexes () were calculated at T Eqn (1), with T Eqn (1) = the time when 4000 nmol NO3−N g−1 soil had been recovered as (NO+N2O+N2)-N.
Table S1. The experimental treatments for each field experiment.
Table S2. Water content of soils (based on soil fresh weight) in the liming experiment.