
Figure 1. Retinocollicular map development and the roles of EphBs.
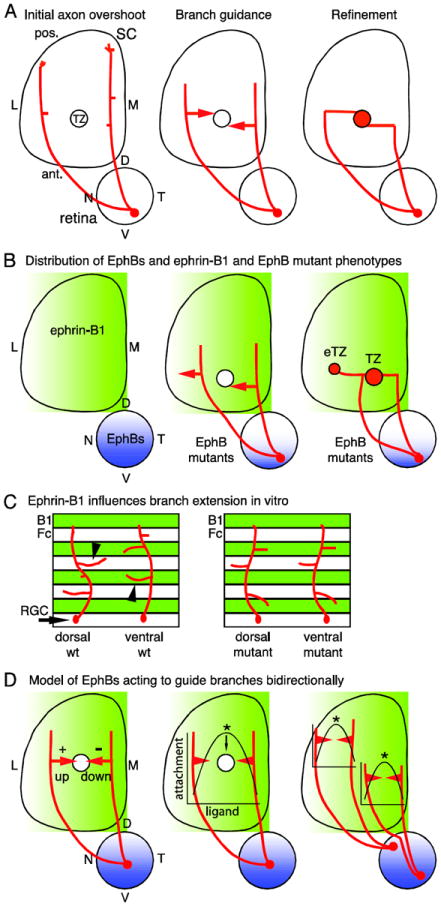
(A) Retinal ganglion cell (RGC) axons enter the superior colliculus (SC) at its anterior (ant.) border and overshoot the future termination zone (TZ) into posterior (pos.) SC (left). Axons then form branches which extend along the medial (M) - lateral (L) SC axis in a guided manner towards the TZ (middle). Branches that reach the nascent TZ arborize and contribute to a dense array of connections (right). D, dorsal; N, nasal; T, temporal; V, ventral. (B) EphB1, EphB2, EphB3, and EphB4 are distributed in RGCs in an overall high ventral - low dorsal gradient (blue) and ephrin-B1 is distributed in a high-M-to-low-L gradient (green) in the SC during retinocollicular map development. In EphB mutant mice, RGC axons from VT retina form a TZ in the appropriate location, but in addition form an ectopic TZ (eTZ) laterally (center). (C) In vitro, dorsal retinal axons prefer to extend branches on control lanes (Fc), avoiding ephrin-B1. Ventral retinal axons prefer to extend branches on ephrin-B1 lanes (B1). Both preferences are lost in EphB mutants. (D) A potential model for the roles of EphBs is informed by in vitro studies (Huynh-do et al., 1999). RGCs have a different point on the ephrin-B1 gradient at which adherence is maximal, dependent on their EphB level. Thus, distinct branch responses to the ephrin-B1 gradient are based not only on the EphB level of the parent RGC, but also the position of the branch on the gradient.