

Abstract
Sepsis is a clinical syndrome characterized by a multisystem response to a pathogenic assault due to underlying infection that involves a combination of interconnected biochemical, cellular and organ–organ interactive networks. After the withdrawal of recombinant human-activated protein C (rAPC), researchers and physicians have continued to search for new therapeutic approaches and targets against sepsis, effective in both hypo- and hyperinflammatory states. Currently, statins are being evaluated as a viable option in clinical trials. Many agents that have shown favourable results in experimental sepsis are not clinically effective or have not been clinically evaluated. Apart from developing new therapeutic molecules, there is great scope for for developing a variety of drug delivery strategies, such as nanoparticulate carriers and phospholipid-based systems. These nanoparticulate carriers neutralize intracorporeal LPS as well as deliver therapeutic agents to targeted tissues and subcellular locations. Here, we review and critically discuss the present status and new experimental and clinical approaches for therapeutic intervention in sepsis.
Table of Links
| TARGETS | LIGANDS | ||
|---|---|---|---|
| C5a receptor | All trans-retinoic acid (ATRA), tretinoin | IL-6 | PAF, platelet activating factor |
| Caspase | Atorvastatin | IL-8 (CXCL8) | PMX53 |
| CXCL2 | C5a complement | IL-10 | Simvastatin |
| CXCR2 | Cathelicidin LL-37 | IL-12 | TNFα |
| ER-β, oestrogen receptor β | Chloroquine | IL-33 | |
| Haem oxygenase 1 | Curcumin | LPS | |
| HMG-CoA reductase | DHEA, dehydroepiandrosterone | Metyrapone | |
| NOS | Eritoran | Mifepristone (RU486) | |
| TLR4, Toll-like receptor 4 | IL-1β | PACAP, pituitary adenylate cyclase-activating polypeptide |
This Table lists key protein targets and ligands in this article which are hyperlinked to corresponding entries in http://www.guidetopharmacology.org, the common portal for data from the IUPHAR/BPS Guide to PHARMACOLOGY (Pawson et al., 2014) and are permanently archived in the Concise Guide to PHARMACOLOGY 2013/14 (Alexander et al., 2013a,b,c,d).
Introduction
Despite much effort to develop effective drugs, sepsis is the second most common cause of death in non-coronary intensive care units and is the tenth leading cause of death in the United States (Angus et al., 2001). Annually, over 750 000 people are affected by sepsis with high morbidity (more than 210 000 deaths) (Angus et al., 2001). According to recent studies, sepsis contributes over 6% of total deaths in the United States, with an average increment of 1.5% every year. Sepsis forecasts are predicting a catastrophic rise to about 1 110 000 cases annually by 2020 (Melamed and Sorvillo, 2009).
The mortality rates associated with the systemic inflammatory response syndrome (SIRS), sepsis, severe sepsis and septic shock are 7, 16, 20 and 46% respectively. Although the percentage of mortality with sepsis is higher in Europe than in the United States, the proportion of patients with single organ failures is higher in the United States (Levy et al., 2012).
In clinical terms, sepsis is basically a systemic inflammatory response to severe infection or tissue damage, leading to hyperactive and unbalanced levels of pro-inflammatory mediators. It leads to an increase in vascular permeability, changes in cardiac function and alteration of metabolic balance, which results in tissue necrosis, multiple organ failure, hypotension, hypoperfusion and finally death (Rittirsch et al., 2008). Sepsis occurs when the SIRS is accompanied by infections such as pneumonia and urinary tract infection. SIRS is generally defined by the presence of two or more of the clinical features listed in Figure 1. Sepsis followed by septic shock is the most severe form of sepsis and is associated with hypotension, which is refractory to adequate fluid resuscitation. In this review, emphasis has been laid on the discussion of pathophysiology of sepsis and current experimental, clinical and possible approaches being employed for the reduction of the clinical outcomes of sepsis and septic shock.
Figure 1.
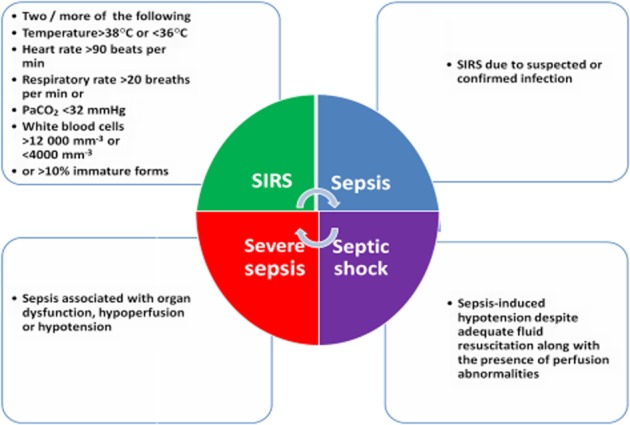
Consensus conference definitions of systemic inflammatory response syndrome, sepsis, severe sepsis and septic shock.
Pathophysiology of sepsis
Sepsis is inflammatory dys-regulation exhibited after bacterial infection. Microorganisms have special structural components known as pathogen-associated molecular patterns (PAMPS), such as LPS (endotoxin) in Gram-negative bacteria and peptidoglycans in Gram-positive bacteria. These foreign molecules interact with host cell receptors known as Toll-like receptors (TLRs) expressed by host immune response cells such as macrophages. Among TLRs, TLR4 selectively recognizes bacterial LPS or endotoxins (Beutler, 2002) and transduces their signal to the interior of cells through NF-κB, leading to transcription of pro-inflammatory cytokines such as TNF-α, IL-1 and IL-6 (Anas et al., 2010). The TLRs have intracellular domains, which interact with intracellular adaptor proteins, such as the myeloid differentiation factor 88 (MYD88), that relay the agonist signal and induce activation of NF-κB-mediated genes. This interaction between the macrophage and the foreign molecule causes the release of early mediators, such as TNF-α, IL-8, IL-1β and IL-6 (van der Poll and Opal, 2008) and late inflammatory mediators in order to protect the host from the microbial insult.
When exposed to a burst of these mediators secreted by macrophages, the endothelial cells undergo activation, causing the release of cytokines, pro-coagulant molecules, platelet-activating factor and NO, leading to the disruption of the endothelial barrier (Hack and Zeerleder, 2001). Endothelial activation promotes the migration of neutrophils and phagocytes to infected tissue, which leads to increased vascular permeability, changes of vascular tone, microvascular thrombosis, intravascular coagulation and finally shock (Hack and Zeerleder, 2001; Remick, 2007).
In severe stages of sepsis, the pro-inflammatory mediators also deregulate the coagulation cascade (both extrinsic and intrinsic clotting pathways) by impairing the activities of tissue factor pathway inhibitor (TFPI), antithrombin, the protein C–activated protein C (APC) system and fibrinolysis (van der Poll and Opal, 2008). TFPI is an endothelial cell-derived protease inhibitor that inhibits the activity of factor Xa. Antithrombin inhibits thrombin factors such as Xa and IXa. Protein C is a vitamin K-dependent plasma serine protease that has an anticoagulant activity (Griffin et al., 2007) and is activated when thrombin is bound to thrombomodulin co-factor. APC has the ability to inactivate the clotting co-factors Va and VIIIa and plasminogen activator inhibitor 1. Deficiency of protein C (due to reduction of thrombomodulin activity by TNF-α) results in abnormalities in coagulation. Therefore, administration of recombinant-activated protein C (rAPC) has been used as an approach to treat sepsis. High mobility group box-1 (HMGB-1), a novel late pro-inflammatory mediator that is bound to nuclear DNA, plays a major role in nucleosome stabilization, gene transcription and acts as a major mediator of systemic inflammation (Andersson and Tracey, 2003). HMGB-1 causes tissue injury and organ dysfunction at the late stage of lethal sepsis, acts on TLR4 and sustains the inflammatory response produced by early mediators by stimulating the release of TNF-α, IL-1β, IL-6 and other pro-inflammatory mediators from macrophages/monocytes (Andersson and Tracey, 2003). Although the initial release of inflammatory mediators by macrophages protects the host but their excessive release a hyper inflammatory state, which causes the activation of the complement system and the formation of the potent pro-inflammatory peptide, C5a (complement component 5a) and other complement products in the circulation, which leads to loss in control of complement activation. C5a has also the ability to induce coagulation cascade and release pro-inflammatory mediators, such as macrophage migration inhibitory factor (MIF) and HMGB-1, which play a prominent role in the pathogenesis of sepsis (Rittirsch et al., 2008). Interaction of C5a with its receptor C5aR on neutrophils triggers signalling pathways and releases powerful chemotactic attractants for neutrophils (LTB4) and induces aggregation of platelets, via TxA2. C5a also enhances the function of LPS, causing increased endothelial cell production of the IL-8 family of CXC-chemokines, other CC-chemokines and IL-6. In the presence of C5a, macrophages also produce pro-inflammatory mediators such as TNF-α and a variety of chemokines, on stimulation by LPS. The decrease in plasma levels of TNF and IL-6 after blockade of C5aR indicates that C5a, C5aR and IL-6 are all involved in the pathogenesis of sepsis (Czermak et al., 1999).
LPS and pro-inflammatory mediators, such as TNF-α, IFN-γ and C5a, are strong inducers of MIF production by leukocytes (Riedemann et al., 2004). MIF also enhances expression of TLR4 on macrophages, which recognize foreign molecules and release pro-inflammatory mediators. At high concentrations, MIF produces sustained inflammatory responses due to its inhibitory activity on p53-dependent apoptosis of activated macrophages. MIF has a special property that it links the immune system with the endocrine system. Another important effect of MIF contributing to the pathogenesis of sepsis is its antagonism of the anti-inflammatory effects of endogenous steroids (Calandra and Roger, 2003).
A recent study has shown that platelets were involved in the pathogenesis of sepsis by inducing the formation of neutrophil extracellular traps (NETs). NETs are nuclear- or mitochondrial-derived DNA strands formed and released by neutrophils, coated with histones and antimicrobial proteins and are capable of capturing and killing bacteria, fungi and parasites. During sepsis, both neutrophils and platelets are recruited to the small capillaries of the liver, lung and other organs. Platelets interact with and activate neutrophils to release NETs from it. Prior to interaction of platelets with neutrophils, platelets must recognize the bacterial surface molecule LPS through TLR4. Although the release of NETs from neutrophils leads to the killing of microbes, it simultaneously exposes dangerous neutrophil enzymes that can damage the endothelium and vital organs, especially the liver and lungs (Urban and Zychlinsky, 2007). The pathophysiology of sepsis is summarized in Figure 2.
Figure 2.

Pathophysiology and major targets for combating sepsis.
LPS removal by liver
Triglyceride (TG)-rich lipoproteins protect animals from endotoxemic death by two complementary mechanisms: (i) by binding endotoxin and inhibiting its activity directly and (ii) by accelerating endotoxin clearance from plasma. All lipoproteins, including TG-rich VLDL (very-low-density lipoprotein) and chylomicrons, and cholesterol ester-rich HDL (high-density lipoprotein) and LDL (low-density lipoprotein) (Shao et al., 2012) have been shown to bind and inactivate endotoxin in vitro (as measured by the Limulus assay). Lipoproteins also block the ability of endotoxin to induce secretion of the cytokines TNF, IL-1 and IL-6 from macrophages in cell culture (Feingold and Grunfeld, 2011). In caecal ligation and puncture (CLP) models of sepsis, the endotoxin bound to circulating TG-rich particles would be effectively masked from the reticuloendothelial system, hence reducing endotoxin-induced stimulation of macrophages and secretion of TNF-α.
TG-rich lipoproteins can also prevent macrophage activation, especially in liver, by redirecting endotoxin metabolism (Feingold and Grunfeld, 2011). Chylomicron-bound endotoxin is transported via the route of chylomicron transport rather than by the endotoxin molecule. Chylomicrons accelerate the clearance of endotoxin from plasma by increasing uptake of endotoxin by the liver, and chylomicron/endotoxin complexes, like chylomicrons alone (Shao et al., 2012), are rapidly cleared by hepatocytes, thereby bypassing the Kupffer cells, which finally results in lower peak serum TNF-α levels (Shao et al., 2012) (Figure 3). TG-rich lipoproteins can bind to LPS and thus modulate the host response to this toxic macromolecule by (i) inhibiting the activation of macrophages, monocytes and other LPS responsive cells; (ii) promoting the catabolism of LPS by the hepatic parenchymal cells; and (iii) inhibiting the response of hepatocytes to pro-inflammatory stimuli.
Figure 3.
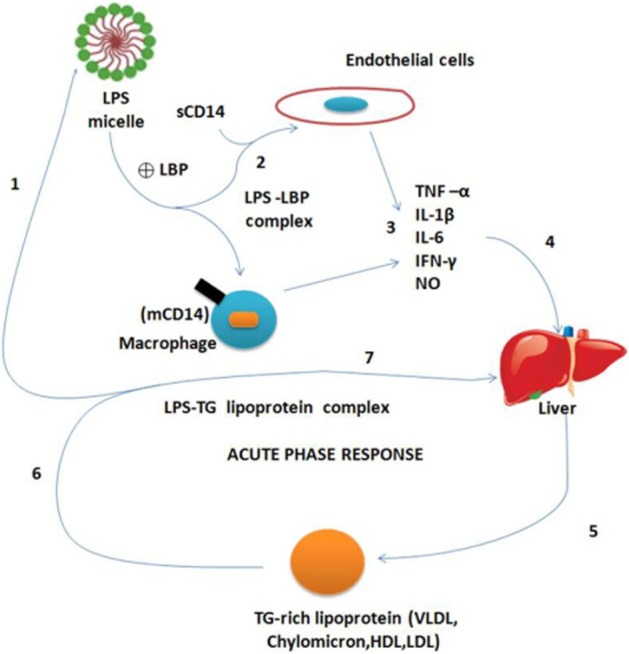
Possible mechanisms for modulation of the host inflammatory response by triglyceride (TG)-rich lipoproteins. (1) LPS enters in blood circulation and (2) stimulates macrophages and endothelial cells to (3) produce cytokines and other mediators of inflammation, including TNF-α, IL-1β and nitric oxide; LPS triggers cell activation by interaction with proteins, including LBP and membrane-bound and soluble forms of CD14 (mCD14). Cytokines secreted by LPS-stimulated cells (4) are responsible for hepatic stimulation, leading to a dramatic alteration in hepatic protein synthesis – ‘the acute phase response’. This cytokine-mediated change in gene expression includes (5) increased production and release of TG-rich lipoproteins (VLDL), and LBP, a 60 kDa protein with lipid transfer activity capable of (6) ‘binding’ LPS to lipoproteins. TG-rich lipoproteins modulate the host response to LPS by first, the formation of lipoprotein–LPS complexes which scavenge the LPS still in the circulation, thus neutralizing its toxicity. Secondly, the complexes are (7) cleared by the liver in a manner that dampens the hepatic response to further cytokine stimulation, thereby modulating the acute phase response. sCD14, soluble CD14.
Organ-specific pathophysiology
Sepsis induced multiple organ dysfunctions and injury is the leading cause of death among critically ill patients. Major organs affected by LPS-induced organ injury are liver, lung, kidney, and cardiovascular and microcirculatory systems. Excessive production of inflammatory mediators induced by LPS and underlying infection leads to coagulation, endothelial damage and microvascular leakage, resulting in organ dysfunction and injury.
Lung
Endothelial leakage of fluids and proteins into the interstitium, infiltration of activated neutrophils, loss of surfactant, mononuclear cell infiltration, type II pneumocyte proliferation and interstitial fibrosis are some of the major events that occur during early inflammatory process in lungs after infection (Greenlee et al., 2007). Reactive oxygen species (ROS) also play an important role in the pathogenesis of lung injury, which mainly affects inflammatory response through redox signalling (Abraham and Singer, 2007). Respiratory failure in patients may further exacerbate tissue damage to lung as well as other organs. Mechanical ventilation is essential in these patients to maintain normal blood oxygen levels in severe cases and may increase tissue damage by inducing release of pro-inflammatory cytokines (Pereira et al., 2013). These events also lead to polymorphonuclear leukocyte migration and activation, causing the release of proteases such as matrix metalloproteinases, which leads to active destruction of alveolar microstructure by degradation of elastin and collagen, worsening the gaseous exchange in lung, leading to respiratory failure (von Ranke et al., 2013).
Kidney
Sepsis is one of the leading causes of acute kidney injury (AKI). Patients with sepsis who develop AKI are at increased risk of death. Experimental studies indicate that endotoxemia-related AKI is associated with both haemodynamic and pro-inflammatory perturbations. Renal vasoconstriction markedly contrasts with sepsis-induced generalized systemic vasodilatation, which is strongly dependent upon NO (Boffa and Arendshorst, 2005). Hyperactivation of systemic renin-angiotensin system during sepsis is well known (Almeida et al., 2006). The pro-inflammatory cytokines, specifically TNF-α, produced by renal mesangial, tubular and endothelial cells, are involved in the renal damage (Donnahoo et al., 1999). NO may also be involved in sepsis-induced renal dysfunction that results in enhanced renal blood flow, particularly to the medulla. NO and atrial natriuretic peptide (ANP) may induce vascular relaxation by increasing the production of cGMP, an important mediator of vascular tone during sepsis. The decrease in ANP clearance was found to play a role in the pathogenesis of LPS-induced kidney injury (Bae et al., 2011). The dysfunction of epithelial barriers and endothelium has been reported in sepsis models (Abraham and Singer, 2007).
Liver
Initiation of hepatic injury usually results from decreased hepatic arterial blood flow after septic shock (primary dysfunction). This can lead to poor hepatic microcirculation, resulting in acute cellular and mitochondrial injury, leading to elevated hepatic transaminases, severe hypoglycaemia and reduced production of coagulation factors. These outcomes are usually restored as perfusion is restored (Dhainaut et al., 2001). Kupffer cells, which play an important role in bacterial products and inflammatory cytokines, are damaged during primary dysfunction, leading to reduced efficacy in removal of these products, resulting in the spillage of bacterial products and inflammatory cytokines to liver parenchyma, resulting in damaged sinusoidal endothelium and hepatocytes. This also leads to increased neutrophil infiltration. Excessive numbers of neutrophils in liver tissue can increase oxidative damage in the liver, leading to necrosis (Goya and Torisu, 1993)
Cardiovascular system
The heart is an important target organ that is structurally damaged by severe sepsis/septic shock. In sepsis, myocardial depression is observed in patients with low and elevated cardiac outputs. Myocardial depression is mainly due to elevated levels of TNF-α, IL-1β, IL-6, lysozyme C and NO. Along with these biochemical mediators, microvascular dysfunction, deregulation of β1-adrenoceptors and dys-regulated changes in Ca2+ ions in the cytoplasm of myocytes also play a very significant role in the reduction of myocardial function (Celes et al., 2013). Troponin I, a marker of myocardial injury, appears in the blood and its levels are increased during sepsis (Abraham and Singer, 2007). The myocardial changes in long-term human severe sepsis/septic shock can be classified within the group of specific cardiomyopathies and named inflammatory cardiomyopathy. Based upon experimental studies, cardiac pathological changes associated with reduced myocardial contractility in severe sepsis/septic shock can be ascribed to the defect in the mechanical connection between the intracellular cytoskeleton of cardiomyocytes and the extracellular matrix, mechanical and electrical/chemical uncoupling of cardiomyocytes and increased sarcolemmal permeability and structural changes in the contractile apparatus of cardiomyocytes. Therapeutic interventions leading to modulation of intracellular calcium and oxidative damage in cardiomyocytes can be a viable approach to provide beneficial effects on cardiovascular outcomes, leading to reduction in mortality induced by sepsis.
Microcirculatory failure
During sepsis/septic shock, in vivo microcirculation is substantially altered. These alterations include reduced capillary blood flow mainly due to systemic pressure reduction and local arteriolar constriction. In severe sepsis, the microcirculation is markedly decreased in the early phases, resulting in detrimental effects mainly caused by hypoperfusion and flare in tissue infection (Remick, 2007), ultimately resulting in widespread capillary dilation. Hypoperfusion occurs mainly in the microcirculatory system due to decreased perfusion pressure, decreased elasticity of red and white cells, constricted arterioles, circulating obstructive fragments (including haemoglobin) and plugging of microvessels with thrombus and adherence of cells to capillary and venular epithelial membranes resulting in increased resistance to flow, loss of fluid and plasma proteins through abnormal transcapillary exchange, differential vascular resistance changes between various beds (e.g. intestinal vs. muscle) and relative absence of regulatory neurohumoral control of small vessel segments of the circulation (Ince, 2005). Hypoperfusion in microcirculatory vessels during shock stops the normal passage of cellular elements, including erythrocytes and neutrophils. As a result of the predominance of many destructive factors, a subsequent round of tissue damage may occur. Because of prolonged capillary vascular stasis, deficient flow and factors released from injured cells, the microcirculation becomes a trap for uncontrolled bacterial growth enhanced by sustained hypoxaemia, acidosis and toxaemia. These events may combine to contribute to the loss of normal cell integrity and death of the host.
Metabolomics: advanced approach for assessment of mortality in sepsis
In case of sepsis, selection of optimized dose and duration of therapy in clinical settings and a readily measured biomarker is usually necessary (Marshall, 2014). Most of the biomarkers employed in preclinical models fail to clearly define the clinical outcomes in sepsis due to the complex nature of the pathophysiology of sepsis (Fry, 1996).
Metabolomics is a new biological technique that seems to be a viable strategy for the identification and validation of physiologically relevant biomarkers that are not covered by other profiling techniques such as proteomics and genomics. This actually focuses upon the identification and measurement of metabolite concentrations, employing NMR spectroscopy and MS coupled with other advanced techniques, leading to established quantitative data sets for multiple endogenous metabolites. It is an emerging component of the systems biology approach for the discovery of clinically relevant biomarkers and potential therapeutic targets (Stringer et al., 2011).
Recently, several reports demonstrated metabolomic approaches for better prediction of mortality outcomes in sepsis in experimental, as well as clinical, settings (Dyson and Singer, 2009; Marshall, 2014). Liu et al. (2010) identified characteristic metabolites, including hypoxanthine, indoxyl sulfate, glucuronic acid, gluconic acid, proline, uracil, nitrotyrosine, uric acid and trihydroxy cholanoic acid involved in oxidative stress and tissue damage, and might supply evidence for distinguishing burned septic rats from non-septic ones (Liu et al., 2010). In another study, septic rats were differentiated from non-septic rats on the basis of the presence of increased concentrations of alanine, creatine, phosphoethanolamine and myoinositol concentrations in lung tissues, and increased concentration of alanine, creatine, phosphoethanolamine and acetoacetate in serum (Izquierdo-Garcia et al., 2011).
CLP in murine models or LPS in vitro resulted in a significant decrease in hepatic oxygen consumption. There was a significant decrease in oxidative phosphorylation measured at 12 h. LPS also resulted in a significant increase in anaerobic respiration in hepatocytes. Interestingly, metabolomic analysis disclosed a metabolic shift in the liver from carbohydrate-based energy to utilization of fatty acids and amino acids. This included an increase in every tricarboxylic acid cycle intermediate and derivative, suggesting an increased flux into the cycle from fatty acid β-oxidation and anaplerotic contributions from amino acids (Park, 2013).
Approaches to therapeutic intervention in sepsis
The molecular pathophysiology of sepsis is highly complex and thus provides many molecular targets for therapeutic intervention (Figure 4). Approaches for therapeutic intervention of sepsis may be subdivided into two classes, namely experimental and clinical approaches (Table 1).
Figure 4.

Scheme of experimental and clinical approaches used in the therapeutic interventions in sepsis.
Table 1.
Approaches to therapeutic interventions in sepsis
| S. No. | Approach | Class/Drug | Mechanism of action | References |
|---|---|---|---|---|
| 1 | Experimental | LPS sequestrants | Neutralize LPS activity | Burns et al., 2005; Puddu et al., 2010; Vicent et al., 2010; Zheng et al., 2010 |
| 2 | HMGB-1 inhibitors | Block HMGB-1 activity | Ulloa et al., 2002; Matthay and Ware, 2004; Chen et al., 2005; Tang et al., 2008; 2009 | |
| 3 | MIF inhibitors | Act against MIF | Tohyama et al., 2008 | |
| 4 | Antimicrobial peptides | Antibacterial and LPS-neutralizing activity | Yibin et al., 2005; Shai et al., 2006 | |
| 5 | Curcumin | Inhibits the transcriptional activity of NF-κB | Kang et al., 2004 | |
| 6 | Diacerhein | Attenuates the synthesis and activity of pro-inflammatory cytokines | Calisto et al., 2012 | |
| 7 | Andrographolide sulfonate | Suppresses the expression of TNF-α, IL-1β, IL-6 | Guo et al., 2012 | |
| 8 | Clinical | Recombinant activated protein C | Blocks factors Va and VIIIa, inhibits cytokine production | Matthay, 2001 |
| 9 | TLR4 antagonists | Block TLR4 | Sha et al., 2007; Opal et al., 2013 | |
| 10 | C5a receptor (C5aR) antagonists and C1 inhibitors | Prevent activation of complement system | Strachan et al., 2000; Charchaflieh et al., 2012 | |
| 11 | Endotoxin removal devices | Bind and neutralize LPS activity | Davies and Cohen, 2011 | |
| 12 | Caspase inhibitors | Anti-apoptic activity | Weber et al., 2009 | |
| 13 | Oestrogen receptor-β agonists | Suppress transcription of pro-inflammatory genes | Cristofaro et al., 2006 | |
| 14 | Statins | Inhibit LPS-induced activation of NF-κB and antioxidant | Fraunberger et al., 2009 |
Experimental approaches
LPS sequestrants
The toxicity of LPS is mainly due to the glycolipid component, lipid A. Neutralization of lipid A by small molecules requires two protonable positive charges spaced so that the distance between them are equivalent to the distance between the two anionic phosphates on lipid A. Synthetic and natural peptides such as polymyxin B (PMB), peptoid 7, derivatives of anti-Limulus factor and antimicrobial peptides have been experimentally evaluated and found to be useful as LPS sequestrants by various research groups. Apart from these agents, spermine derivatives are also being used as experimental agents for LPS neutralization.
PMB is a membrane-active peptide antibiotic that sequesters LPS and neutralizes its activity. It has higher affinity for binding to LPS than any other peptide. However, the use of PMB has been limited due to severe systemic toxicity (Novelli et al., 2012). To overcome this limitation of PMB, several molecules have been developed experimentally that bind to LPS with high affinity but with less toxicity. Peptoid 7, a synthetic peptide derivative and novel LPS-neutralizing molecule conjugated with polyethylene glycol (PEG) to overcome the problems of poor aqueous solubility and non-specific toxicity, has demonstrated improved survival in a murine model of sepsis (Vicent et al., 2010). A cyclic peptide, CLP-19 (Limulus anti-LPS factor derivative), has been developed as a LPS-neutralizing agent to treat septic shock having good LPS binding activity. It also improves the survival of septic animal models by down-regulating LPS-induced release of TNF-α (Vallespi et al., 2003). Lactoferrin, a multifunctional protein belonging to the transferrin family, shows antibacterial, antiviral, LPS-neutralizing and immunological activities (Gifford et al., 2005). The antibacterial activity is mainly due to enhancement of bacterial permeability to various agents by disturbing bacterial membrane through binding to Ca2+ present on its surface (Farnaud and Evans, 2003). Lactoferrin also has the ability to neutralize free LPS directly, thereby preventing its action on TLR-4 and subsequent expression of pro-inflammatory cytokines (Mattsby-Baltzer et al., 1996; Elass et al., 2002; Puddu et al., 2010). The bactericidal/permeability-increasing protein (BPI) is an endogenous antibiotic protein that has potent anti-endotoxin and anti-cytotoxic activities against Gram-negative bacteria. Bactericidal neutralizing endotoxin protein (BNEP), a synthetic peptide derived from the human BPI, shows anti-endotoxin activity similar to BPI. Along with LPS-neutralizing activity, BNEP also reduces the levels of TNF-α and IL-6 by decreasing the circulating levels of endotoxin and protects animals from the effects of LPS. BNEP and PMB bound LPS with a similar affinity. Therefore, BNEP could protect animals from the lethal effects of LPS through decreasing plasma endotoxin and pro-inflammatory cytokines (Jiang et al., 2004).
Antimicrobial peptides, such as mesoparan and cathelicidins, have also found their usefulness due to their dual action, antibacterial and LPS-neutralizing activity. Mastoparan-1 (MP-1), a cationic amphiphilic tetradecapeptide toxin isolated from hornet venom, has both antibacterial and LPS-neutralizing activity. Its antibacterial action against Escherichia coli is due to the destruction of bacterial membrane structures. MP-1 has high affinity for LPS and lipid A and neutralizes LPS in a dose-dependent manner, but the affinity is lower than that of PMB. MP-1 also reduces the expression of TLR4, TNF-α, IL-6 mRNA and release of cytokines induced by LPS (Yibin et al., 2005).
Cathelicidins are cationic host defence peptides that show antimicrobial, LPS-neutralizing and immunomodulatory activities when C-terminal peptides are activated by proteolytic cleavage. The antimicrobial activity is mainly due to pore formation in the cell membrane and interaction with DNA, RNA and proteins. LPS-neutralizing activity was useful in treating sepsis when there are high levels of pro-inflammatory cytokines circulating in plasma, stimulated by LPS (Shai et al., 2006). Cathelicidins also prevent the LPS-induced apoptosis of endothelial cells. Cathelicidins, such as human LL-37 and bovine BMAP-28, have been shown to improve survival in mice with sepsis by decreasing LPS-mediated cytokine production (Giacometti et al., 2004; Cirioni et al., 2006).
A novel lipopolyamine, 1,3-di-oleoyloxy-2-(6-carboxy spermyl)-propylamide (DOSPER] has been described that binds to LPS with affinity of 1/10 that of PMB (David et al., 1999). Lysine–spermine conjugates neutralize bacterial LPS with the ε-amino terminus of the lysinyl moiety, which binds with long-chain aliphatic hydrophobic substituents in acryl or alkyl linkage (Burns et al., 2005).
Geniposide, a new anti-sepsis agent extracted and isolated from Gardenia jasminoides Ellis, shows a direct, neutralising action on LPS. Geniposide also down-regulates the expression of pro-inflammatory mediators following activation of TLR4 and also inhibits phosphorylation of p38 MAPK. It inhibits TNF-α mRNA expression but has no effect on the levels of TNF-α induced by CpG DNA (Zheng et al., 2010).
HMGB-1 inhibitors
HMGB-1 inhibitors have been found to be beneficial in experimental sepsis. Low MW compounds such as ethyl pyruvate, nicotine, stearoyl lysophosphatidylcholine, chloroquine, persicarin and isorhamnetin-3-O-galactoside and peptides such as pituitary adenylate cyclase-activating polypeptide (PACAP) inhibit HMGB-1 in cell cultures, as well as in appropriate animal models.
Ethyl pyruvate, a derivative of pyruvic acid, inhibits activation of the pro-inflammatory transcription factors, NF-κB and p38 MAPK, by blocking two signalling pathways that are essential for cytokine release. Ethyl pyruvate also decreased the expression of pro-inflammatory mediators including inducible nitric oxide synthase (iNOS), TNF-α, COX-2 and IL-6 in LPS-stimulated RAW 264.7 murine macrophage-like cells (Ulloa et al., 2002; Yang et al., 2002). During sepsis, inflammatory responses cause organ dysfunction and mortality. Treatment with ethyl pyruvate has been shown to improve survival and/or ameliorate organ dysfunction in animal models of severe sepsis. Yang et al. (2002) stated that ethyl pyruvate acts as an immunological modulator and ameliorated physical and immunological barrier dysfunction in sepsis (Yang et al., 2002).
Nicotine is an alkaloid that also shows anti-inflammatory action by binding to the nicotinic acetylcholine receptors present on the surface of macrophages. Blockade of these receptors affects the NF-κB pathway and attenuates the release of HMGB-1 (Matthay and Ware, 2004), TNF-α and iNOS via inhibiting haem oxygenase 1 (Tsoyi et al., 2011). Inhibitors of cholinesterase, such as physostigmine or neostigmine, activate the cholinergic anti-inflammatory pathway and decrease the levels of pro-inflammatory cytokines. Although these inhibitors improve survival in sepsis, when treatment was delayed, benefit in reduction of mortality was not observed (Fodale and Santamaria, 2008). Stearoyl lysophosphatidylcholine, another HMGB1 inhibitor that suppresses endotoxin, induced release of HMGB1 from macrophages. Additionally, it removes foreign microorganisms through H2O2-dependent mechanisms by stimulating neutrophils. Therefore, stearoyl lysophosphatidylcholine improves the survival of animal models in experimental sepsis (Chen et al., 2005). Quercetin also suppresses the release and pro-inflammatory activity of late HMGB1 cytokine (Tang et al., 2009).
Persicarin and isorhamnetin-3-O-galactoside inhibit LPS- and CLP-mediated HMGB-1 release and HMGB-1-mediated barrier disruption by increasing barrier integrity and by inhibiting the expression of cell adhesion molecules, thereby reducing leukocyte adhesion to HUVECs and leukocyte migration. The treatment was effective when given in two doses in early phase (12 h after CLP) and late phase of treatment (50 h after CLP) and was not effective in reducing mortality at single dose at 12 h after CLP (Kim et al., 2013a,b).
Chloroquine, as well as its derivative hydroxychloroquine, by inhibiting HMGB1 release has been demonstrated to have therapeutic potential for lethal sepsis and systemic inflammation in an animal model of lethal endotoxemia and sepsis in vivo (Yang et al., 2013).
Immune cells synthesize PACAP when they are exposed to endotoxin. PACAP is an endogenous neuropeptide that acts by attenuating the LPS-induced HMGB1, TNF-α and IL-6 release from macrophages. PACAP does not induce IL-10 directly, but increased production of IL-10 in LPS-stimulated macrophages indicates its anti-inflammatory activity. Therefore, PACAP attenuates the inflammatory response by modifying the release of various mediators and improve the survival in sepsis and septic shock (Tang et al., 2008).
Macrophage migratory factor (MIF) inhibitors
During inflammation, macrophages, T-cells and pituitary gland activate and release a pleiotropic cytokine MIF, which is an important mediator of septic shock. The corresponding monoclonal antibodies (mAbs) protect mice from septic shock by neutralizing MIF (Thierry Calandra et al., 2001). The main drawback with the mAbs is local and systemic inflammatory reactions during administration and, to avoid this complication, a DNA-based vaccine that protects from sepsis by generating endogenous anti-MIF antibodies has also been developed. Active immunization with the MIF/tetanus toxin DNA vaccine attenuates sepsis-related inflammation by decreasing the elevated levels of TNF, IL-1β and IL-6 especially in the lungs and protects the lung from acute injury (Tohyama et al., 2008).
NF-κB inhibitors and antioxidants
Polyphenols such as curcumin and flavonoids have been widely reported to have antioxidant and anti-inflammatory activities. There are numerous preclinical studies reporting the usefulness of treatment of sepsis in terms of enhancement of survival in various animal models of sepsis.
Curcumin. the yellow pigment present in the rhizomes of turmeric Curcuma longa Linn., Zingiberaceae, has been used for treating inflammatory and infectious diseases in traditional Chinese and Indian medicine by virtue of its anti-inflammatory, antioxidant, antiproliferative, and pro-apoptotic, antibacterial and anti-cancer activity (Sun et al., 2010). The anti-inflammatory activity of curcumin is mainly due to its ability to inhibit the transcriptional activity of NF-κB, activator protein-1, COX-2 expression and TREM-1 (triggering receptor expressed on myeloid cells-1) in macrophages (Yuan et al., 2012; Kang et al., 2004), which results in inhibition of pro-inflammatory cytokine release (Singh et al., 2013). Curcumin also bound to myeloid differentiation factor 2 (MD-2) and competitively inhibited binding of LPS to TLR4 and suppressed LPS-induced inflammatory response and improved survival in LPS-induced sepsis in mice (Gradisar et al., 2007).
Quercetin is a flavonoid derivative and shows anti-inflammatory action by reducing the levels of NO, IL-6 (via inhibition of LPS-induced activation of MAPK), TNF-α and HMGB1 through inhibition of the NF-κB pathway (Comalada et al., 2005), increases mRNA expression of anti-inflammatory enzyme haem oxygenase-1 (Heeba et al., 2014), thereby decreasing susceptibility of neutrophils to LPS (Liu et al., 2005).
In addition to curcumin and quercetin, polyphenols such as epigallocatechin-3-gallate and paeonol have also demonstrated beneficial effects by reducing inflammation induced by LPS. Epigallocatechin-3-gallate inhibited NF-κB, thereby reducing NO production and TNF-α and by inhibiting late (e.g. HMGB1) pro-inflammatory cytokines. Naringin, another flavonoid, was ameliorated LPS-induced sepsis in mice, via the NF-κB pathway, and to reduced LPS-induced lung inflammation by inhibiting MPS and iNOS activity (Liu et al., 2011b). Pretreatment with paeonol in RAW 264.7 macrophages and mice inhibited NF-κB and MAPK activities and cytokines (TNF-α, IL-6, IL-1β and IL-10) in vitro and rescued mice from lethal sepsis. Pro- and anti-inflammatory cytokines were significantly down-regulated, and thus the survival rates of LPS-challenged mice were improved by paeonol (Chen et al., 2014). Degradation products of polyphenols such as ferualdehyde also improve survival in murine model of sepsis through reduction in levels of TNF-α and anti-inflammatory cytokine IL-10 (Radnai et al., 2009).
Diacerhein is an anthraquinone derivative that has anti-inflammatory, analgesic and antipyretic activity. In sepsis, pro-inflammatory pathways are activated, releasing inflammatory mediators. Hypoglycaemia and insulin resistance are induced by excess production of these mediators, leading to a hormonal imbalance. Pro-inflammatory cytokines activate intracellular pathways involving IκB kinase/NF-κB and JNK, which are associated with insulin resistance (Marik and Raghavan, 2004). Diacerhein attenuates the synthesis and activity of pro-inflammatory cytokines such as TNF-α and IL-6 via inhibition of the activation of NF-κB and reduces the insulin resistance and higher levels of fasting plasma glucose in animals with sepsis (Calisto et al., 2012).
Andrographolide is a natural diterpenoid, from Andrographis paniculata, and it shows antibacterial, anti-inflammatory, anti-tumour, anti-angiogenic, antiviral, hepatoprotective and neuroprotective activities (Guo et al., 2012). Andrographolide sulfonate, a water-soluble form of andrographolide, protected against sepsis in animal models and reduced endotoxic shock and death of animals. Andrographolide sulfonate suppresses expression of TNF-α, IL-1β, IL-6 and iNOS and decreased the levels of TNF-α and IL-1β (Guo et al., 2012). It also inhibited the activation of MAPK, NF-κB and STAT3 (Guo et al., 2012), which are important in the transcriptional induction of several genes, such as those for iNOS,COX-2, TNF-α, IL-1β and IL-6, involved in inflammation.
Recently, an extracellular polysaccharide from Bacillus sp. strain LBP32 (EPS) has also been found to demonstrate inhibition of LPS-induced release of pro-inflammatory mediators, such as NO, ROS, IL-6 and TNF-α via inhibition of NF-κB activation. In vivo experiments were conducted and showed that EPS could greatly improve the outcome of mice with LPS-induced endotoxic shock (Diao et al., 2014).
Bis-N-norgliovictin, a low MW compound from a marine fungus, dose-dependently inhibited LPS-induced production of TNF-α, IL-6, IFN-β and CCL2 (MCP-1), but without suppressing macrophage viability and improves survival in sepsis via attenuating liver and lung damage (Song et al., 2013). Recently, therapeutic administration of lithium (25 and 50 mg kg−1) in a CLP model of sepsis in rats after 16 h has been reported to decrease IL-1β, IL-6 and TNF-α in the serum, and the 8-iso-prostaglandin F2α (8-ISO) levels, whereas increasing level of superoxide dismutase (SOD) and the total levels of GSH in the lung tissues of rats reduced the inflammation in the lung (Albayrak et al., 2013).
Clinical approaches
Recombinant activated protein C (rAPC)
rAPC was the first anti-sepsis agent to be approved for treating severe sepsis (Bernard et al., 2001b). This protein decreases the production of thrombin by blocking coagulation factors Va and VIIIa, resulting in inhibition of platelet activation, neutrophil migration and mast cell degranulation (Matthay, 2001). rAPC has anti-inflammatory properties such as inhibition of cytokine production by monocytes and cell adhesion. Clinical trials (phase II) of the rAPC (drotrecogin alfa; Xigris®, Eli Lilly) in patients with severe sepsis demonstrated that it decreased the coagulopathy and the sepsis-associated inflammatory response (Bernard et al., 2001a). Administration of rAPC decreased mortality and improved survival in patients with sepsis at high risk of death (Dhainaut et al., 2004). The U.S. Food and Drug Administration (FDA) and the European Medicines Agency approved the use of rAPC in 2001 (Toussaint and Gerlach, 2009). The major adverse effect with rAPC is bleeding and about 2.5% of patients had intracranial haemorrhage with open-label use of rAPC (Warren et al., 2002). Currently, rAPC is approved only for use in patients with sepsis who have severe organ compromise and the highest chances of death (Toussaint and Gerlach, 2009). In October 2011, Eli Lilly withdrew Xigris from the pharmaceutical market following the negative results of the placebo-controlled PROWESS SHOCK trial (Dolgin, 2012).
TLR4 antagonists
Endotoxin induces a pro-inflammatory response and systemic coagulation by acting on TLR4 receptors and modifies signalling pathways through the adaptor protein MD-2. TAK-242 is a novel cyclohexane derivative that directly binds to TLR4. This agent inhibits LPS-induced production of NO and cytokines such as TNF-α, IL-1, IL-6 and IL-10 (Sha et al., 2007). Even after successful completion of phase III clinical trials,Takeda discontinued TAK-242 in 2010, because it failed to decrease cytokine levels in patients with severe sepsis and septic shock. Eritoran tetrasodium (ETS) (a second-generation synthetic LPS) blocks LPS activity by competing for binding to MD-2, which, in turn, activates TLR4 receptor. This binding prevents intracellular signalling pathways, production of NF-κB and LPS-induced pro-inflammatory cytokines (Mullarkey et al., 2003). After a clinical trial (phase 3) on 2000 patients, Eritoran was withdrawn due to its failure to show therapeutic benefits, that is, reduction in 28 days and 1 year mortality (Opal et al., 2013). Failure of the eritoran trial demonstrated the importance of inflammatory pathways triggered by PAMPS, in addition to those targeted by LPS, in the mortality due to sepsis (Tse, 2013).
C5aR antagonists and complement system inhibitors
PMX53, a cyclic peptide, is a C5aR antagonist that inhibits C5a or LPS-induced neutropenia and cytokine (TNF-α and IL-6) production. The potent anti-inflammatory activities of this C5aR antagonist play a key role in C5a-induced sepsis (Strachan et al., 2000). A polyclonal IgG antibody against C5a has also demonstrated beneficial effects in CLP-induced sepsis (Czermak et al., 1999). The inflammatory responses due to the activation of the complement system can be inhibited by blocking C activation. The C1 inhibitor (C1-INH) is a serine proteinase inhibitor that blocks complement activation and improves survival in patients with sepsis but with lesser effects on organ dysfunction (Charchaflieh et al., 2012).
Endotoxin removal devices
Nowadays there is increasing emphasis on extracorporeal treatment, in which circulating endotoxins and inflammatory mediators in the blood are removed by appropriate adsorbents. Yin et al. (2012) developed l-serine grafted poly(vinylidene fluoride) membrane fibres for extracorporeal removal of endotoxin, TNF-α and IL-6 (Yin et al., 2012). As sepsis is mediated also by inflammatory cytokines, these were removed by haemodiafiltration using a cytokine-adsorbing haemofilter, made from polymethyl methacrylate (Hirasawa et al., 2012). Although PMB has the highest affinity for bacterial endotoxin, its clinical use has been limited because of its toxicity. To avoid this toxicity, investigators have developed an endotoxin removal extracorporeal device to remove endotoxin (Toramyxin®) which consists of an affinity column coated with PMB. Extracorporeal passage of blood through this column removes endotoxin from blood, without systemic toxicity. It has been approved and is marketed in Europe and Japan since 2002 and 1994, respectively, and is in phase III clinical trials in the United States and Canada (Davies and Cohen, 2011). Recently, Miltenyi Biotech has developed amphiphilic core-shell nanosorbents to remove endotoxin from protein preparations. These are magnetic nanoparticles that are coated with polycationic ligands that remove LPS from protein solutions with an efficiency of 99% (Cai et al., 2012).
Caspase inhibitors
The majority of people die with sepsis due to immunosupression or immunoparalysis, which is follows excessive apoptosis in the intestine, lymphoid organs and lymphocytes. Caspases are a subfamily of cysteine proteases that are critical for apoptosis in sepsis (Salvesen and Riedl, 2008) and caspase inhibitors should provide a good approach to intervention in sepsis. Recently, a novel caspase inhibitor, VX-166, which has potent anti-apoptotic activity, inhibited the release of IL-1β and IL-18 and improved the survival in LPS-induced sepsis (Weber et al., 2009).
Oestrogen receptor agonists
The biological activity of oestradiol is mediated through the oestrogen receptors (ER-α and -β) which modulate anti-inflammatory response. The ER-β is an important therapeutic target because it is not involved in the proliferative effect of oestrogens in mammary gland and uterus. ER-β agonists exhibit anti-inflammatory activities through suppression of transcription of pro-inflammatory genes (Cvoro et al., 2008). Cristofaro et al. (2006) showed that a non-steroidal selective ER-β agonist, WAY-202196, has protective activity against septic shock.
Statins
Statins are inhibitors of 3-hydroxy-3-methyl glutaryl coenzyme A (HMG-CoA) reductase, used to treat hyperlipidaemia and also for protection from cardiovascular dysfunction. They inhibit the synthesis of isoprenoids and geranyl-geranylpyrophosphate, which are formed from the mevalonate pathway. Statins have anti-inflammatory, anti-apoptotic, anti-proliferative, antithrombotic, antimicrobial and endothelium protecting activities. Because of these pleiotropic effects, statins have been used as adjunctive agents in the treatment of sepsis (Falagas et al., 2008). Statins modify the intercellular interactions and cellular chemotaxis of the immune system, and they attenuate the release of cytokines and acute phase proteins. They have antioxidant properties, even though they reduce ubiquinone levels, which is an endogenous antioxidant. Statins inhibit the expression of certain genes, modify the activity of cells and are involved in various mechanisms of the inflammatory response. Simvastatin showed a marked reduction in mortality by protection of cardiac function and reversal of inflammatory responses in a burn-induced sepsis model in mice (Beffa et al., 2011). Simvastatin inhibits LPS-induced activation of NF-κB and release of TNF by blocking the activity of HMG-CoA reductase (Fraunberger et al., 2009). Supporting the above study, Dobesh et al. (2009) demonstrated that statin treatment of patients, before the development of severe sepsis, appeared to produce a significant reduction in mortality, compared with patients who were not treated with statins.
Recent approaches for therapeutic interventions in sepsis
Pharmaceutical approaches
Targeting of therapeutic agents to specific tissues or cell types has been widely reported to enhance the therapeutic activity of molecules and reduce the toxicity in a range of diseases, such as cancer and metabolic diseases (Shukla and Srivastava, 2012). Marketed parenteral emulsions like Intralipid® exhibit many of the same properties as chylomicrons and may prevent septic death by a similar mechanism (Figure 4). Chylomicron and Intralipid particles are both rich in TG and are of similar size. Intralipid particles acquire apolipoproteins once in the circulation and, like chylomicrons, are rapidly cleared by hepatocytes (Shao et al., 2012). Synthetic TG-rich emulsions, as well as chylomicrons, have the capacity to inactivate endotoxin in vitro and to prevent death from a lethal dose of endotoxin in animals (Hultin et al., 1995). In porcine and equine models of sepsis, beneficial effects of phospholipid-enriched emulsion were clearly demonstrated by reduction in serum endotoxin, cytokines and TNF-α level. Liposomal delivery of ATP or NAD+ protects human endothelial cells from energy failure in a cell culture model of sepsis after 72 h of LPS/IFN exposure, indicating a potential approach in treatment of sepsis (Han et al., 2001).
Polymers such as chitosan and low-molecular-weight polyethylene glycol (LMW-PEG) can also affect the outcome of sepsis. Chitosan is a linear polymer consisting of 2-amino-2-deoxy-d-glucopyranose and 2-acetylamido-2-deoxy-d-glucopyranose residues linked through a β-1,4-glycosidic bond and have been used in various formulations to deliver wide range of drugs (Shukla et al., 2010; Singodia et al., 2010; Jain et al., 2014). Chitosan can directly interact with LPS and form a stable LPS–chitosan complex. The induction of TNF-α and IL-6 biosynthesis by LPS–chitosan complex is lower than that of the parent LPS (Yermak et al., 2006). Chitosan protected mice from sepsis by decreasing oxidative tissue damage and pro-inflammatory mediators (Qiao et al., 2011). Chitosan oligosaccharides have been reported to possess antimicrobial, anti-oxidative and anti-tumour, and immunopromoting activities and reduced inflammatory responses (release of TNF-α, IL-6) induced by LPS by blocking the binding of LPS to TLR4/MD-2 receptor (Mishra, 2011) and inhibiting MAPK and PI3K/Akt signalling pathways (Ma et al., 2011). Recently, chitosan-capped gold nanoparticles were shown to reduce LPS toxicity (Stefan et al., 2013). A derivative of chitosan, that is, zwitterionic chitosan, also reduces the pro-inflammatory activity of LPS by strongly binding to it and suppressing the release of pro-inflammatory mediators from macrophages (Bajaj et al., 2012). Kim et al. (2012) developed glycol chitosan-conjugated eicosapentaenoic acid nanoparticles, which showed anti-inflammatory properties by inhibiting NO production and IL-1β secretion induced by LPS (Kim et al., 2012).
LMW-PEG reduces the inflammatory response induced by bacterial endotoxin by decreasing the levels of cytokines (TNF-α and IL-6) in the circulation. LMW-PEG also inhibits the streptolysin-induced necrosis of human neutrophils and LPS-induced activation of pro-apoptotic pathways. It reduced the mortality by greater than 50% in both LPS and zymosan models of sepsis. LMW-PEG has potent anti-inflammatory properties and reduces mortality in models of severe systemic inflammation and sepsis (Ackland et al., 2010).
PEG-conjugated SOD has been demonstrated to have a very good antioxidant activity, leading to reduction of oxidative stress in endothelium, heart, lung, liver and kidney, and was able to protect the organs from damage and could be a promising approach to treatment of sepsis and sepsis-related organ damage (Veronese et al., 2002).
Recently, a group has synthesized Pi-PEG15-20 (phosphorylated-high molecular weight PEG) as an anti-virulence agent with an observation that inorganic phosphate (Pi) was able to reduce virulence in a variety of pathogens. This molecule limited the ability of colonizing pathogens to express virulence in response to host-derived and local environmental factors. Pi-PEG15-20 not only prevented colonization of the introduced pathogen community but also suppressed the propagation of opportunistic pathogen members of the commensal microflora and reduced pro-inflammatory potential of the caecal contents (Zaborin et al., 2014).
Mitochondria-targeted delivery of SOD by using a pH-sensitive polyketal polymer, poly(cyclohexane-1,4-diyl acetone dimethylene ketal (PCADK; Yang et al., 2008) has been proposed for effective therapeutic intervention of inflammatory disorders like sepsis. Polymer-encapsulated SOD was efficiently targeted to phagosomes (due to acidic pH-induced degradation of PCADK) and was able to reduce ROS generation by the mitochondria of macrophages. The main advantage of the polyketal polymer was that it has neutral degradation products, compared with polyanhydrides, and thus has a lower potential to enhance inflammation (Lee et al., 2006).
Liposomal delivery of therapeutic agents has also been reported to decrease the severity of sepsis and to limit organ injury. Liposomes loaded with dexamethasone and anti-TNF-α siRNA, TJU-2755 (Ponnappa et al., 2001), have been used for prophylaxis to alleviate LPS-induced tissue injury in sepsis (Suntres and Shek, 2000). Liposomal dichloromethylene-diphosphonate decreased plasma TNF-α, CXCL2 (MIP-2) and IL-10 in mice depleted of tissue macrophages, compared with control groups of mice, although it was not helpful in reducing the severity of bacteremia or survival of septic mice (Fujimoto et al., 2002). Delivery of proanthocyanidins via chitosan-coated liposomes induced a dose-dependent attenuation of iNOS and COX-2 expression in LPS-stimulated macrophages (Madrigal-Carballo et al., 2009). Brahmamdam et al. (2009) used a novel cyclodextrin polymer-based transferrin targeted delivery vehicle to administer siRNA targeted for two critical mediators of apoptosis, Bim and PUMA (cell death proteins), in a mouse model of CLP-induced sepsis. The study demonstrated that this targeted approach markedly decreased lymphocyte apoptosis and prevented the loss of splenic CD4 T- and B-cells (Brahmamdam et al., 2009).
Application of nanotechnology for drug delivery in sepsis faces a real challenge in any successful translation from preclinical phase to clinical settings (Opal and Patrozou, 2009). Currently, there are very few reports pertaining to application of nanotechnology for delivery of antisepsis agents ( Table 2). The main impediment for nanotherapeutics is the very slow translation from experimental stages to the clinic. So far, there are very few nanotechnology (i.e. Abraxane and Doxil)-based therapeutic agents approved by the FDA (Park, 2013) but the positive side is that the number of such agents in clinical trials have increased and it can be assumed that, in the near future, more nanotechnology-based drug delivery systems will be clinically available. The choice of appropriate nanocarriers is not clear, because of the lack of comparative studies, and suitable protocols to determine the optimal characteristics of nanocarriers remain to be defined. These issues are important because several factors may simultaneously affect biodistribution of nanocarriers in humans (Peer et al., 2007). Triggering of sensitization reactions by nanoparticulate systems should be thoroughly evaluated as there are reports of generation of antibodies against fullerenes (Chen and Hongjie, 2003) and dendrimers (Lee, 2001; 2004). Apart from this, there are numerous questions about nanotoxicological issues associated with nanotherapeutic agents (Huh and Kwon, 2011; Park, 2013). The challenge lies in proper evaluation of interaction of nanotherapeutic agents with tissues and organs in order to recalibrate the doses according to clinical needs and identification of proper administration procedures in order to obtain desired therapeutic effects.
Table 2.
Different nanoparticle-based approaches for experimental treatment of sepsis
| S. No. | Formulations | Therapeutic outcome | Reference |
|---|---|---|---|
| 1 | Liposomes loaded ATP or NAD+ | Protect human endothelium from energy failure in cell culture models | Han et al., 2001 |
| 2 | Ciprofloxacin-loaded lipid emulsion | Animal survival improved in LPS-induced sepsis mice model | Mishra, 2011 |
| 3 | Chitosan capped gold nanoparticles | Protective effects against LPS-induced toxicity in rats | Stefan et al., 2013 |
| 4 | Antisense oligonucleotides in pH-sensitive liposomes | LPS-induced production of TNF-α in rats | Ponnappa et al., 2001 |
| 5 | Liposomal dexamethasone | Prophylaxis against LPS-induced lung injuries in rats | Suntres and Shek, 2000 |
| 6 | Curcumin exosomes | Mice treated with exosomal curcumin have much lower mortality than mice treated with an equivalent concentration of liposomal curcumin | Sun et al., 2010 |
| 7 | Minocycline nanoliposomes | Reduction in LPS induces TNF-α by macrophages at lower doses | Liu and Yang, 2012 |
| 8 | Puerarin and curcumin loaded gold nanoparticles | Suppression of LPS-induced inflammation and cytotoxicity in rats | Singh et al., 2013 |
| 9 | Glycyrrhizic acid nanoparticles | Inhibition of LPS-induced inflammatory mediators in 264.7 mouse macrophages | Wang et al., 2013 |
| 10 | Red blood cell-mediated delivery of dexamethasone | Selective inhibition of NF-κB activation and TNF-α production in macrophages | Crinelli et al., 2000 |
| 11 | Ciprofloxacin-bearing cationic nanoemulsions | Reduction in LPS released from E. coli and reduction in cytokine secretion in E. coli-induced peritonitis in rats | Jain et al., 2014 |
| 12 | Parenteral triglyceride-rich emulsions | Have the capacity to inactivate endotoxin in vitro and to prevent death from a lethal dose of endotoxin in animals | Hultin et al., 1995 |
| 13 | Peptoid 7 PEG micelles | LPS neutralization in vivo improves survival in mice with sepsis | Vicent et al., 2010 |
Nanoparticles are used to improve bioavailability and pharmacokinetics or are employed for targeting loaded therapeutic agents to specific organs, which may improve therapeutic outcome in sepsis. However, the therapeutic efficacy at any stage of sepsis is dependent upon the mechanism of action of therapeutic agents. For example, the stages at which neutralization of LPS or TNF-α might benefit would differ from that stage, which might benefit from administration of granulocyte macrophage colony-stimulating factor (GM-CSF), a growth factor for myeloid cells or agents acting on HMGB-1, which is a late pro-inflammatory agent. Nanocarriers, that is, chitosan-coated gold nanoparticles (Stefan et al., 2013) and parenteral lipid nanoemulsion (Hultin et al., 1995), interacted with LPS in vivo to neutralize and redirect LPS to excretion pathways in the body.
Targeted delivery of anti-sepsis molecules to the site of action can be achieved by passive or active targeting to specific tissues, so that the effect of anti-sepsis molecules can be enhanced. Novel drug delivery approaches can be explored further for their effect on modification of the molecular pharmacology of anti-sepsis molecules.
Biomedical approaches
Stem cells possess a variety of anti-inflammatory, antimicrobial, anti-apoptotic and cytoprotective properties that could ameliorate sepsis-induced inflammation and organ injury. Mesenchymal stem cells (MSCs) are non-haematopoietic precursor cells derived from a variety of tissues, including bone marrow, adipose, placenta and umbilical cord. MSCs have the ability to repair injured tissues, paracrine signalling effects, immunomodulatory capacity and are also potential antimicrobial agents. These properties improve organ dysfunction and survival in animal models of sepsis. The TNF-stimulated gene is an anti-inflammatory factor secreted by MSCs, that interacts with the CD44 receptor of resident macrophages and decreases the expression of TNF-α via down-regulating NF-κB signalling pathways. The cathelicidin LL-37 peptide shows antimicrobial properties by disrupting bacterial membranes and is secreted by MSCs in response to E. coli (Wannemuehler et al., 2012). A recent study has shown that administration of human MSCs reduces mortality in mice with sepsis by enhancing bacterial clearance and phagocytic activity of monocytes in blood (Krasnodembskaya et al., 2012).
Agents inhibiting neutrophil migration
The clearance of pathogenic microorganisms from circulation depends upon neutrophil migration into the infectious site. The chemokine receptor CXCR2 is a key mediator controlling neutrophil migration from the systemic circulation to the site of infection. During sepsis, TLRs are activated and increase levels of G protein-coupled receptor kinase 2 (GRK2), a protein kinase that plays a major role in the phosphorylation and down-regulation of CXCR2, leading to inhibition of neutrophil migration. The cytokine IL-33 is an endogenous molecule belonging to the IL-1 family expressed by fibroblasts, epithelial, endothelial, mast and innate immune cells in response to infection. IL-33 prevents TLR-mediated down-regulation of CXCR2 and promotes neutrophil migration by suppressing the expression of GRK2. This phenomenon has been demonstrated experimentally, where IL-33-treated mice showed increased neutrophil influx into the peritoneal cavity and bacterial clearance compared with untreated mice. Therefore, increasing IL-33 would be an important therapeutic approach to attenuate sepsis (Alves-Filho et al., 2010).
Kukoamine B (a natural spermine alkaloid from the Chinese herb Cortex lycii) inhibits both LPS and CpG DNA, decreasing release of TNF-α and IL-6, which improves survival in sepsis compared with other drugs (PMB) that target only LPS (Liu et al., 2011a).
Translation of anti-sepsis therapy from bench to bedside: a major challenge
There are many reports indicating potential advantages of a variety of therapeutic agents in animal models of sepsis, but the majority of them have failed in clinical settings (Table 3). This may be attributed to lack of extrapolation of findings from animal models to clinical sepsis (Celes et al., 2013). To establish correlation between preclinical trials and subsequent clinical studies, we need a suitable animal model mimicking outcomes in clinical settings (Dyson and Singer, 2009). It is very unlikely that a single therapeutic molecule could ever be useful in treating the widespread dys-regulation of the inflammatory response that occurs during septic shock. Recently, sepsis has been demonstrated to have an early inflammatory phase and later immunosuppressive stages with higher risk of mortality in both phases. So, trials involving a combination of therapeutic intervention for both phases should be of more value in improving clinical outcomes. Mortality in the immunosuppressive phase are typically due to failure to control the primary infection or the acquisition of secondary hospital-acquired infections, often with opportunistic pathogens (Hotchkiss et al., 2013). In addition to this, patient-specific factors, such as age, gender, co-morbidities and genetic predisposition, also play key roles in determining the individual response to therapy.
Table 3.
Current status of anti-sepsis agents in clinical trials
| S. No. | Drug | Company | Mechanism | Phase/Comments |
|---|---|---|---|---|
| 1 | Toraymyxin | Spectral Diagnostics | Removes endotoxin | IIIb |
| 2 | Alkaline phosphatase | AM-Pharma | Detoxifies bacterial Products | II |
| 4 | Eritoran (E5564) | Eisai | Antagonizes TLR4 | III; failed to reduce 28 day and 1 year sepsis-induced mortality |
| 5 | Lactoferrin-α | Agennix | Immunomodulatory and Antibacterial properties | II/IIIa |
| 6 | CytoFab (AZD9773) | Astra Zeneca | Fab fragments of IgG that bind to TNF-α | II; further clinical development has been stopped |
| 7 | Resatorvid (TAK 242) | Takeda | Antagonize TLR4 | III; further clinical development has been stopped |
| 8 | SRT2379 | GlaxoSmithKline | Sirtuin-1 activator | I |
| 9 | Bicizar | BioGenius | C1-esterase inhibitor blocks complement activation | III |
| 10 | Citrulline | Vanderbilt University | Precursor of NO | II |
| 11 | Hydrocortisone | Charite University, Berlin, Germany | Alters cytokine response | III |
| 12 | Recombinant human soluble thrombomodulin (ART-123) | Artisan Pharma, Inc. | Binds to thrombin | II |
| 13 | Anti-TF mAbs (ALT-836) | Altor Bioscience Corporation | Blocks TF induced inflammation | II |
| 14 | Atorvastatin | Hospital de Clinicas de Porto Alegre | Suppression of the TLR-dependent inflammatory pathway | II |
Data were obtained from http://www.clinicaltrials.gov (as of 22 October 2012).
The OASIS trial of talactoferrin alfa should be completed by 2014.
EUPHRATES trial of endotoxin removal by haemoperfusion through a polymyxin B column is scheduled for completion in 2013 (The Lancet Infectious Diseases, 2012).
When designing clinical trials in sepsis, critically ill patients should be stratified according to severity of illness in order to maximize the signal-to-noise ratio of new therapeutic agents. Defined human target populations and established severities of sepsis would allow for more realistic and applicable animal models (Riedemann et al., 2003). The focus of future clinical trials should be on more stratified populations with either specific diseases, that is, community-acquired pneumonia or peritonitis or specific biomarkers to identify patients who are likely to benefit from the intervention; for example, if one is seeking to test an antibody to TNF-α, one should enrol only patients with pathologically high circulating levels of TNF-α (Lopez et al., 2004; Tse, 2013). In most of the cases, correlation between pharmacokinetic/pharmacodynamic (PK/PD) data and experimental efficacy for anti-sepsis molecules is also lacking. This deficiency can also be a major contributor to the unsuccessful translation of experimental studies to clinical levels. Thus, gaining information about PK/PD correlation of a particular therapeutic intervention can be more relevant and may result in improving clinical outcomes (Fry, 1996).
Immunostimulants in sepsis-induced immunosuppression
Survival of patients after the early phase of sepsis (the cytokine storm) is improved by a better treatment protocol, leading to their entry into a protracted immunosuppressive phase. Survival in this later phase is usually dependent upon the degree of immunosuppression. Patients suffering from severe immunosuppression, as indicated by a reduced expression of HLA-DR (MHC class II cell surface receptor encoded by the human leukocyte antigen) in myeloid cells, after the early phase of sepsis, frequently have infectious foci despite the antimicrobial therapy, or reactivation of latent viral infections such as cytomegalovirus (Giamarellos-Bourboulis, 2013).
Improvement in survival was associated with decreased lymphocyte apoptosis, restoration of innate immune cell populations, increased pro-inflammatory cytokines and restoration of IFN-γ production by stimulated splenocytes on treatment with AS101 (ammonium trichloro-dioxoethylene-o,o′ tellurate) a compound blocking production of IL-10, a key mediator of immunosuppression in a mouse model of sepsis (Muenzer et al., 2010). Immunostimulatory therapy with biological molecules such as cytokines (IL-7 and IL-15) has demonstrated efficacy in models of infection including sepsis by preventing apoptosis-induced depletion of immune cells and diminishing immunosuppressive effect on phagocytic cells (Hotchkiss and Opal, 2010). GM-CSF shortened the time of mechanical ventilation as well as ICU stay by restoring monocyte HLA-DR expression and cytokine release in patients with sepsis and sepsis-associated immunosuppression, but improvement in 28 day survival was not evident (Meisel et al., 2009). All trans-retinoic acid (ATRA) has been reported to restore immunocompetence and improve the primary humoral immune response, even when immunosuppression was established first and ATRA was administered subsequently, mainly by modulating the number of leucocytes and the survival of myeloid-derived suppressor cells (Martire-Greco et al., 2014).
Benjamim et al. (2005) exhibited the usefulness of intrapulmonary administration of bone marrow-derived DCs in controlling pulmonary Aspergillus infection in immunocompromised post-septic mice. This effect was mediated by promoting the balance of pro-inflammatory and suppressive cytokines in the lung, thereby restoring the normal immune function in post-septic mice (Benjamim et al., 2005). Primary, secondary and T-cell-mediated immune responses were restored with mifepristone (RU486), an antagonist of glucocorticoid receptors (Rearte et al., 2010). Other modulators of glucocorticoids, such as dehydroepiandrosterone, a hormone with anti-glucocorticoid properties, or metyrapone, an inhibitor of corticosterone synthesis, were able to partially, but significantly, restore the humoral immune response in immunosuppressed mice (Rearte et al., 2013). A significant recovery of splenocyte proliferation and restoration of distinct cellular components leading to reorganization of spleen architecture was also observed, resulting in restoration of the hypersensitivity response, with these treatments.
The value of thymosin α1, a naturally occurring thymic peptide, as a promising beneficial immunomodulatory drug in therapeutic intervention of sepsis-related immunosupression was demonstated in a clinical trial. Thymosin α1 resulted in an increase in survival rates of patients having severe sepsis and improved mHLA-DR level in blood on the third and seventh day of treatment (Wu et al., 2013).
Expert opinion
Inflammation has been known to act as double-edged sword in case of severe infection and sepsis. Mediators, specifically pro-inflammatory cytokines, play a central role in the pathophysiology of sepsis by contributing to organ injury, shock and death. However, they are also vital components in the clearance of microbial infections from tissues and in case of innate immunity, specifically in late phase of sepsis where immunosuppression becomes an important aspect of pathophysiology. Failure of most of the experimental strategies in the therapy of sepsis in clinical settings may be ascribed to the fact that most of therapeutic strategies are focused on preventing early deaths by attenuation of hyperactive immune system at initial stages rather than combating the progressive development of immunosuppression in late phases of sepsis due to secondary bacterial, fungal and viral infections, which is the main cause of death in late phases of sepsis.
These facts underlie the need of a dual-action compound or a combination of agents to provide a therapy that reduces mortality by acting on both late and early phases of sepsis. preventing the early over-activation of immune response and reversing the immunosuppression in late phases of sepsis. This goal can be achieved by opting for a logical therapeutic regimen comprising a combination of different agents for early and late phases of sepsis in clinical settings to achieve a better therapeutic response. Agents such as curcumin could provide better therapeutic outcomes in clinical settings because of its effects on several targets in the pathophysiological pathway of sepsis.
It is also likely that binding of LPS by extracellular molecules may not be enough to target the bound complex to the liver parenchyma and metabolic inactivation and therefore, it is necessary to elucidate the pathway of targeting the bound complex to the liver, using pharmaceutical approaches. In our laboratory, we have obtained good evidence for anti-endotoxin treatments. As serotype-specificity or epitope-specificity limits the therapeutic applications, it would be beneficial to have a compound capable of non-specifically binding to one or more sites on the LPS chain by employing well-known, safe excipients to achieve hydrophobic as well as electrostatic interaction with LPS. Use of novel drug delivery systems, such as nanoparticles and emulsions, can be tested in clinical settings to improve therapeutic outcome in sepsis by improving bioavailability and tissue distribution affected by sepsis-induced damage. Evaluation of the PK and PD parameters of a new therapeutic intervention has been minimal in experimental and clinical settings. Initial dosing is typically inferred from results in animal models and the final dose selected by an optimistic interpretation of mortality data from a phase II trial. Proper evaluation of PK and PD parameters and their correlation may lead to better monitoring of therapy in experimental and clinical settings.
Conclusion
Sepsis is an inflammatory condition involving a complex interaction of multiple pathways, which leads to fatal outcomes. The array of outcomes due to sepsis may lead to multiple organ failure and ultimately death. Pharmacological agents currently being employed in the intervention of sepsis and related conditions in both clinical and experimental settings are statins, Bicizar® (complement C1-esterase inhibitor) and phenolic compounds such as curcumin and flavonoids. The main problem with most of the experimental agents is their poor PK properties, which may present a significant hurdle in the development of these agents into a potential clinical candidate. Other reasons are the poor correlation in outcomes between preclinical studies and clinical trials hindering the development of experimental anti-sepsis molecules. Extracorporeal removal of endotoxin is still a preferred approach to achieve better survival in clinical settings but there is an urgent need to develop intracorporeal approaches specifically for clinical settings.
Risk of mortality is also high in a hypo-immune condition after the cytokine storm, because of the increased probability of secondary infections, which limits the scope of anti-inflammatory agents in the therapeutic intervention of sepsis. Treatment of secondary infection is still a challenging task in the hypo-immune conditions after sepsis. This clearly dictates the need of agents that are capable of maintaining normal immune function in hyper- as well as hypo-immune states.
Although targeted drug delivery in sepsis is still is in its infancy, there are reports indicating improvement in the therapeutic performance of anti-sepsis molecules, via more efficient drug delivery strategies and this approach still has promise in improving outcomes in clinical settings. Pharmaceutical excipients such as chitosan and PEG have sufficient activity against LPS, which may improve the outcome of sepsis therapy. Phospholipids in the form of emulsions can serve as carriers for anti-sepsis molecules, providing nutrition as well as enhancing removal of endotoxin. To further improve the outcomes via therapeutic agents, targeted drug delivery approaches should be considered as one of the integral components in the drug development programmes for sepsis.
Glossary
- AKI
acute kidney injury
- ANP
atrial natriuretic peptide
- BNEP
bactericidal neutralizing endotoxin protein
- BPI
bactericidal/permeability-increasing protein
- C5aR
complement component 5a receptor
- CLP
caecal ligation puncture
- DHEA
dehydroepiandrosterone
- GM-CSF
granulocyte macrophage colony-stimulating factor
- GRK2
G protein-coupled receptor kinase 2
- HDL
high-density lipoprotein
- HLA-DR
MHC class II cell surface receptor encoded by the human leukocyte antigen
- HMGB-1
high mobility group box-1
- HMG-CoA
3-hydroxy-3-methyl glutaryl coenzyme A
- iNOS
inducible nitric oxide synthase
- LBP
lipopolysaccharide binding protein
- LDL
low-density lipoprotein
- LMW-PEG
low-molecular-weight polyethylene glycol
- mAbs
monoclonal antibodies
- MD-2
myeloid differentiation factor 2
- MIF
migration inhibitory factor
- MP-1
Mastoparan-1
- MSCs
mesenchymal stem cells
- NETs
neutrophil extracellular traps
- PACAP
pituitary adenylate cyclase-activating polypeptide
- PAMPS
pathogen-associated molecular patterns
- PCADK
poly(cyclohexane-1,4-diyl acetone dimethyleneketal
- PEG
polyethylene glycol
- PMB
polymyxin B
- rAPC
recombinant-activated protein C
- ROS
reactive oxygen species
- SIRS
systemic inflammatory response syndrome
- SOD
superoxide dismutase
- TFPI
tissue factor pathway inhibitor
- TG-rich lipoproteins
triglyceride-rich lipoprotein
- TLR
Toll-like receptor
- TREM-1
triggering receptor expressed on myeloid cells-1
- VLDL
very-low-density lipoprotein
Author contributions
P. R. M. and P. S. were actively involved in conceptualization and writing of the manuscript. N. M. and G. M. R. actively participated in drawing graphical representations and tables for the article. G. P., S. S. and R. S. managed search of latest studies and were actively involved in checking manuscript for typographical and grammatical errors. P. R. M., P. S., N. M., G. M. R., G. P., S. S. and R. S. all reviewed and critically appraised the manuscript. All authors agreed on the final version.
Conflict of interest
The authors report no conflict of interest.
References
- Abraham E, Singer M. Mechanisms of sepsis-induced organ dysfunction. Crit Care Med. 2007;35:2408–2416. doi: 10.1097/01.ccm.0000282072.56245.91. [DOI] [PubMed] [Google Scholar]
- Ackland GL, Gutierrez Del Arroyo A, Yao ST, Stephens RC, Dyson A, Klein NJ, et al. Low-molecular-weight polyethylene glycol improves survival in experimental sepsis. Crit Care Med. 2010;38:629–636. doi: 10.1097/CCM.0b013e3181c8fcd0. [DOI] [PubMed] [Google Scholar]
- Albayrak A, Halici Z, Polat B, Karakus E, Cadirci E, Bayir Y, et al. Protective effects of lithium: a new look at an old drug with potential antioxidative and anti-inflammatory effects in an animal model of sepsis. Int Immunopharmacol. 2013;16:35–40. doi: 10.1016/j.intimp.2013.03.018. [DOI] [PubMed] [Google Scholar]
- Alexander SPH, Benson HE, Faccenda E, Pawson AJ, Sharman JL, Spedding M, et al. The Concise Guide to PHARMACOLOGY 2013/14: G Protein-Coupled Receptors. Br J Pharmacol. 2013a;170:1459–1581. doi: 10.1111/bph.12445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander SPH, Benson HE, Faccenda E, Pawson AJ, Sharman JL, Spedding M, et al. The Concise Guide to PHARMACOLOGY 2013/14: Enzymes. Br J Pharmacol. 2013b;170:1797–1867. doi: 10.1111/bph.12451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander SPH, Benson HE, Faccenda E, Pawson AJ, Sharman JL, Spedding M, et al. The Concise Guide to PHARMACOLOGY 2013/14: Nuclear Hormone Receptors. Br J Pharmacol. 2013c;170:1652–1675. doi: 10.1111/bph.12448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander SPH, Benson HE, Faccenda E, Pawson AJ, Sharman JL, Spedding M, et al. The Concise Guide to PHARMACOLOGY 2013/14: Catalytic Receptors. Br J Pharmacol. 2013d;170:1676–1705. doi: 10.1111/bph.12449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almeida WS, Maciel TT, Di Marco GS, Casarini DE, Campos AH, Schor N. Escherichia coli lipopolysaccharide inhibits renin activity in human mesangial cells. Kidney Int. 2006;69:974–980. doi: 10.1038/sj.ki.5000134. [DOI] [PubMed] [Google Scholar]
- Alves-Filho JC, Sonego F, Souto FO, Freitas A, Verri WA, Jr, Auxiliadora-Martins M, et al. Interleukin-33 attenuates sepsis by enhancing neutrophil influx to the site of infection. Nat Med. 2010;16:708–712. doi: 10.1038/nm.2156. [DOI] [PubMed] [Google Scholar]
- Anas AA, Wiersinga WJ, de Vos AF, van der Poll T. Recent insights into the pathogenesis of bacterial sepsis. Neth J Med. 2010;68:147–152. [PubMed] [Google Scholar]
- Andersson U, Tracey KJ. HMGB1 in sepsis. Scand J Infect Dis. 2003;35:577–584. doi: 10.1080/00365540310016286. [DOI] [PubMed] [Google Scholar]
- Angus DC, Linde-Zwirble WT, Lidicker J, Clermont G, Carcillo J, Pinsky MR. Epidemiology of severe sepsis in the United States: analysis of incidence, outcome, and associated costs of care. Crit Care Med. 2001;29:1303–1310. doi: 10.1097/00003246-200107000-00002. [DOI] [PubMed] [Google Scholar]
- Bae EH, Kim IJ, Ma SK, Lee JU, Kim SW. Altered regulation of renal nitric oxide and atrial natriuretic peptide systems in lipopolysaccharide-induced kidney injury. Korean J Physiol Pharmacol. 2011;15:273–277. doi: 10.4196/kjpp.2011.15.5.273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bajaj G, Van Alstine WG, Yeo Y. Zwitterionic chitosan derivative, a new biocompatible pharmaceutical excipient, prevents endotoxin-mediated cytokine release. PLoS ONE. 2012;7:e30899. doi: 10.1371/journal.pone.0030899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beffa DC, Fischman AJ, Fagan SP, Hamrahi VF, Paul KW, Kaneki M, et al. Simvastatin treatment improves survival in a murine model of burn sepsis: role of interleukin 6. Burns. 2011;37:222–226. doi: 10.1016/j.burns.2010.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benjamim CF, Lundy SK, Lukacs NW, Hogaboam CM, Kunkel SL. Reversal of long-term sepsis-induced immunosuppression by dendritic cells. Blood. 2005;105:3588–3595. doi: 10.1182/blood-2004-08-3251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernard GR, Ely EW, Wright TJ, Fraiz J, Stasek JE, Jr, Russell JA, et al. Safety and dose relationship of recombinant human activated protein C for coagulopathy in severe sepsis. Crit Care Med. 2001a;29:2051–2059. doi: 10.1097/00003246-200111000-00003. [DOI] [PubMed] [Google Scholar]
- Bernard GR, Vincent JL, Laterre PF, LaRosa SP, Dhainaut JF, Lopez-Rodriguez A, et al. Efficacy and safety of recombinant human activated protein C for severe sepsis. N Engl J Med. 2001b;344:699–709. doi: 10.1056/NEJM200103083441001. [DOI] [PubMed] [Google Scholar]
- Beutler B. TLR4 as the mammalian endotoxin sensor. Curr Top Microbiol Immunol. 2002;270:109–120. doi: 10.1007/978-3-642-59430-4_7. [DOI] [PubMed] [Google Scholar]
- Boffa JJ, Arendshorst WJ. Maintenance of renal vascular reactivity contributes to acute renal failure during endotoxemic shock. J Am Soc Nephrol. 2005;16:117–124. doi: 10.1681/ASN.2004060441. [DOI] [PubMed] [Google Scholar]
- Brahmamdam P, Watanabe E, Unsinger J, Chang KC, Schierding W, Hoekzema AS, et al. Targeted delivery of siRNA to cell death proteins in sepsis. Shock. 2009;32:131–139. doi: 10.1097/SHK.0b013e318194bcee. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burns MR, Wood SJ, Miller KA, Nguyen T, Cromer JR, David SA. Lysine-spermine conjugates: hydrophobic polyamine amides as potent lipopolysaccharide sequestrants. Bioorg Med Chem. 2005;13:2523–2536. doi: 10.1016/j.bmc.2005.01.038. [DOI] [PubMed] [Google Scholar]
- Cai C, Hsieh CL, Gao S, Kannan A, Bhansali M, Govardhan K, et al. Soluble guanylyl cyclase alpha1 and p53 cytoplasmic sequestration and down-regulation in prostate cancer. Mol Endocrinol. 2012;26:292–307. doi: 10.1210/me.2010-0394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calandra T, Roger T. Macrophage migration inhibitory factor: a regulator of innate immunity. Nat Rev Immunol. 2003;3:791–800. doi: 10.1038/nri1200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calisto KL, Camacho AC, Mittestainer FC, Carvalho BM, Guadagnini D, Carvalheira JB, et al. Diacerhein attenuates the inflammatory response and improves survival in a model of severe sepsis. Critical care. 2012;16:R158. doi: 10.1186/cc11478. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Celes MRN, Prado CM, Rossi MA. Sepsis: going to the heart of the matter. Pathobiology. 2013;80:70–86. doi: 10.1159/000341640. [DOI] [PubMed] [Google Scholar]
- Charchaflieh J, Wei J, Labaze G, Hou YJ, Babarsh B, Stutz H, et al. The role of complement system in septic shock. Clin Dev Immunol. 2012;2012:407324. doi: 10.1155/2012/407324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen G, Li J, Qiang X, Czura CJ, Ochani M, Ochani K, et al. Suppression of HMGB1 release by stearoyl lysophosphatidylcholine: an additional mechanism for its therapeutic effects in experimental sepsis. J Lipid Res. 2005;46:623–627. doi: 10.1194/jlr.C400018-JLR200. [DOI] [PubMed] [Google Scholar]
- Chen N, Liu D, Soromou LW, Sun J, Zhong W, Guo W, et al. Paeonol suppresses lipopolysaccharide-induced inflammatory cytokines in macrophage cells and protects mice from lethal endotoxin shock. Fundam Clin Pharmacol. 2014;28:268–276. doi: 10.1111/fcp.12019. [DOI] [PubMed] [Google Scholar]
- Chen RJ, Hongjie D. Noncovalent functionalization of carbon nanotubes for highly specific electronic biosensors. Proc Natl Acad Sci USA. 2003;100:4984–4989. doi: 10.1073/pnas.0837064100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cirioni O, Giacometti A, Ghiselli R, Bergnach C, Orlando F, Silvestri C, et al. LL-37 protects rats against lethal sepsis caused by gram-negative bacteria. Antimicrob Agents Chemother. 2006;50:1672–1679. doi: 10.1128/AAC.50.5.1672-1679.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Comalada M, Camuesco D, Sierra S, Ballester I, Xaus J, Galvez J, et al. In vivo quercitrin anti-inflammatory effect involves release of quercetin, which inhibits inflammation through down-regulation of the NF-kappaB pathway. Eur J Immunol. 2005;35:584–592. doi: 10.1002/eji.200425778. [DOI] [PubMed] [Google Scholar]
- Crinelli R, Antonelli A, Bianchi M, Gentilini L, Scaramucci S, Magnani M. Selective inhibition of NF-κB activation and TNF-α production in macrophages by red blood cell-mediated delivery of dexamethasone. Blood Cells Mol Dis. 2000;26:211–222. doi: 10.1006/bcmd.2000.0298. [DOI] [PubMed] [Google Scholar]
- Cristofaro PA, Opal SM, Palardy JE, Parejo NA, Jhung J, Keith JC, Jr, et al. WAY-202196, a selective estrogen receptor-beta agonist, protects against death in experimental septic shock. Crit Care Med. 2006;34:2188–2193. doi: 10.1097/01.CCM.0000227173.13497.56. [DOI] [PubMed] [Google Scholar]
- Cvoro A, Tatomer D, Tee MK, Zogovic T, Harris HA, Leitman DC. Selective estrogen receptor-beta agonists repress transcription of proinflammatory genes. J Immunol. 2008;180:630–636. doi: 10.4049/jimmunol.180.1.630. [DOI] [PubMed] [Google Scholar]
- Czermak BJ, Sarma V, Pierson CL, Warner RL, Huber-Lang M, Bless NM, et al. Protective effects of C5a blockade in sepsis. Nat Med. 1999;5:788–792. doi: 10.1038/10512. [DOI] [PubMed] [Google Scholar]
- David SA, Silverstein R, Amura CR, Kielian T, Morrison DC. Lipopolyamines: novel antiendotoxin compounds that reduce mortality in experimental sepsis caused by gram-negative bacteria. Antimicrob Agents Chemother. 1999;43:912–919. doi: 10.1128/aac.43.4.912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies B, Cohen J. Endotoxin removal devices for the treatment of sepsis and septic shock. Lancet Infect Dis. 2011;11:65–71. doi: 10.1016/S1473-3099(10)70220-6. [DOI] [PubMed] [Google Scholar]
- Dhainaut J-F, Marin N, Mignon A, Vinsonneau C. Hepatic response to sepsis: interaction between coagulation and inflammatory processes. Crit Care Med. 2001;29:S42–S47. doi: 10.1097/00003246-200107001-00016. [DOI] [PubMed] [Google Scholar]
- Dhainaut JF, Yan SB, Claessens YE. Protein C/activated protein C pathway: overview of clinical trial results in severe sepsis. Crit Care Med. 2004;32(5 Suppl):S194–S201. doi: 10.1097/01.ccm.0000128035.64448.45. [DOI] [PubMed] [Google Scholar]
- Diao Y, Xin Y, Zhou Y, Li N, Pan X, Qi S, et al. Extracellular polysaccharide from Bacillus sp. strain LBP32 prevents LPS-induced inflammation in RAW 264.7 macrophages by inhibiting NF-κB and MAPKs activation and ROS production. Int Immunopharmacol. 2014;18:12–19. doi: 10.1016/j.intimp.2013.10.021. [DOI] [PubMed] [Google Scholar]
- Dobesh PP, Klepser DG, McGuire TR, Morgan CW, Olsen KM. Reduction in mortality associated with statin therapy in patients with severe sepsis. Pharmacotherapy. 2009;29:621–630. doi: 10.1592/phco.29.6.621. [DOI] [PubMed] [Google Scholar]
- Dolgin E. Trial failure prompts soul-searching for critical-care specialists. Nat Med. 2012;18:1000. doi: 10.1038/nm0712-1000. [DOI] [PubMed] [Google Scholar]
- Donnahoo KK, Meng X, Ayala A, Cain MP, Harken AH, Meldrum DR. Early kidney TNF-alpha expression mediates neutrophil infiltration and injury after renal ischemia-reperfusion. Am J Physiol. 1999;277(3 Pt 2):R922–R929. doi: 10.1152/ajpregu.1999.277.3.R922. [DOI] [PubMed] [Google Scholar]
- Dyson A, Singer M. Animal models of sepsis: why does preclinical efficacy fail to translate to the clinical setting? Crit Care Med. 2009;37:S30–S37. doi: 10.1097/CCM.0b013e3181922bd3. [DOI] [PubMed] [Google Scholar]
- Elass E, Masson M, Mazurier J, Legrand D. Lactoferrin inhibits the lipopolysaccharide-induced expression and proteoglycan-binding ability of interleukin-8 in human endothelial cells. Infect Immun. 2002;70:1860–1866. doi: 10.1128/IAI.70.4.1860-1866.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falagas ME, Makris GC, Matthaiou DK, Rafailidis PI. Statins for infection and sepsis: a systematic review of the clinical evidence. J Antimicrob Chemother. 2008;61:774–785. doi: 10.1093/jac/dkn019. [DOI] [PubMed] [Google Scholar]
- Farnaud S, Evans RW. Lactoferrin – a multifunctional protein with antimicrobial properties. Mol Immunol. 2003;40:395–405. doi: 10.1016/s0161-5890(03)00152-4. [DOI] [PubMed] [Google Scholar]
- Feingold KR, Grunfeld C. The role of HDL in innate immunity. J Lipid Res. 2011;52:1–3. doi: 10.1194/jlr.E012138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fodale V, Santamaria LB. Cholinesterase inhibitors improve survival in experimental sepsis: a new way to activate the cholinergic anti-inflammatory pathway. Crit Care Med. 2008;36:622–623. doi: 10.1097/CCM.0B013E31816297CE. [DOI] [PubMed] [Google Scholar]
- Fraunberger P, Grone E, Grone HJ, Walli AK. Simvastatin reduces endotoxin-induced nuclear factor kappaB activation and mortality in guinea pigs despite lowering circulating low-density lipoprotein cholesterol. Shock. 2009;32:159–163. doi: 10.1097/SHK.0b013e318193c514. [DOI] [PubMed] [Google Scholar]
- Fry DE. The importance of antibiotic pharmacokinetics in critical illness. Am J Surg. 1996;172:20s–25s. doi: 10.1016/s0002-9610(96)00346-7. [DOI] [PubMed] [Google Scholar]
- Fujimoto J, Wiener-Kronish JP, Hashimoto S, Sawa T. Effects of CL2MDP-encapsulating liposomes in a murine model of Pseudomonas aeruginosa-induced sepsis. J Liposome Res. 2002;12:239–257. doi: 10.1081/lpr-120014760. [DOI] [PubMed] [Google Scholar]
- Giacometti A, Cirioni O, Ghiselli R, Bergnach C, Orlando F, D'Amato G, et al. The antimicrobial peptide BMAP-28 reduces lethality in mouse models of staphylococcal sepsis. Crit Care Med. 2004;32:2485–2490. doi: 10.1097/01.ccm.0000148221.09704.22. [DOI] [PubMed] [Google Scholar]
- Giamarellos-Bourboulis EJ. Failure of treatments based on the cytokine storm theory of sepsis: time for a novel approach. Immunotherapy. 2013;5:207–209. doi: 10.2217/imt.13.8. [DOI] [PubMed] [Google Scholar]
- Gifford JL, Hunter HN, Vogel HJ. Lactoferricin: a lactoferrin-derived peptide with antimicrobial, antiviral, antitumor and immunological properties. Cell Mol Life Sci. 2005;62:2588–2598. doi: 10.1007/s00018-005-5373-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goya T, Torisu M. Potential role of hepatic macrophages in neutrophil-mediated liver injury in rats with sepsis. Hepatology. 1993;17:1086–1094. [PubMed] [Google Scholar]
- Gradisar H, Keber MM, Pristovsek P, Jerala R. MD-2 as the target of curcumin in the inhibition of response to LPS. J Leukoc Biol. 2007;82:968–974. doi: 10.1189/jlb.1206727. [DOI] [PubMed] [Google Scholar]
- Greenlee KJ, Werb Z, Kheradmand F. Matrix metalloproteinases in lung: multiple, multifarious, and multifaceted. Physiol Rev. 2007;87:69–98. doi: 10.1152/physrev.00022.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffin JH, Fernandez JA, Gale AJ, Mosnier LO. Activated protein C. J Thromb Haemost. 2007;5(Suppl. 1):73–80. doi: 10.1111/j.1538-7836.2007.02491.x. [DOI] [PubMed] [Google Scholar]
- Guo W, Liu W, Chen G, Hong S, Qian C, Xie N, et al. Water-soluble andrographolide sulfonate exerts anti-sepsis action in mice through down-regulating p38 MAPK, STAT3 and NF-kappaB pathways. Int Immunopharmacol. 2012;14:613–619. doi: 10.1016/j.intimp.2012.09.002. [DOI] [PubMed] [Google Scholar]
- Hack CE, Zeerleder S. The endothelium in sepsis: source of and a target for inflammation. Crit Care Med. 2001;29(7 Suppl):S21–S27. doi: 10.1097/00003246-200107001-00011. [DOI] [PubMed] [Google Scholar]
- Han YY, Huang L, Jackson EK, Dubey RK, Gillepsie DG, Carcillo JA. Liposomal ATP or NAD+ protects human endothelial cells from energy failure in a cell culture model of sepsis. Res Commun Mol Pathol Pharmacol. 2001;110:107–116. [PubMed] [Google Scholar]
- Heeba GH, Mahmoud ME, El Hanafy AA. Anti-inflammatory potential of curcumin and quercetin in rats: role of oxidative stress, heme oxygenase-1 and TNF-alpha. Toxicol Ind Health. 2014;30:551–560. doi: 10.1177/0748233712462444. [DOI] [PubMed] [Google Scholar]
- Hirasawa H, Oda S, Nakamura M, Watanabe E, Shiga H, Matsuda K. Continuous hemodiafiltration with a cytokine-adsorbing hemofilter for sepsis. Blood Purif. 2012;34:164–170. doi: 10.1159/000342379. [DOI] [PubMed] [Google Scholar]
- Hotchkiss RS, Monneret G, Payen D. Sepsis-induced immunosuppression: from cellular dysfunctions to immunotherapy. Nat Rev Immunol. 2013;13:862–874. doi: 10.1038/nri3552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hotchkiss RS, Opal S. Immunotherapy for sepsis – a new approach against an ancient foe. N Engl J Med. 2010;363:87–89. doi: 10.1056/NEJMcibr1004371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huh AJ, Kwon YJ. ‘Nanoantibiotics’: a new paradigm for treating infectious diseases using nanomaterials in the antibiotics resistant era. J Control Release. 2011;156:128–145. doi: 10.1016/j.jconrel.2011.07.002. [DOI] [PubMed] [Google Scholar]
- Hultin M, Carneheim C, Rosenqvist K, Olivecrona T. Intravenous lipid emulsions: removal mechanisms as compared to chylomicrons. J Lipid Res. 1995;36:2174–2184. [PubMed] [Google Scholar]
- Ince C. The microcirculation is the motor of sepsis. Crit Care. 2005;9:S13–S19. doi: 10.1186/cc3753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Izquierdo-Garcia JL, Nin N, Ruiz-Cabello J, Rojas Y, de Paula M, Lopez-Cuenca S, et al. A metabolomic approach for diagnosis of experimental sepsis. Intensive Care Med. 2011;37:2023–2032. doi: 10.1007/s00134-011-2359-1. [DOI] [PubMed] [Google Scholar]
- Jain V, Shukla P, Pal R, Mishra PR. Cationic nanoemulsions bearing ciprofloxacin surf-plexes enhances its therapeutic efficacy in conditions of E. coli induced peritonitis and sepsis. Pharm Res. 2014 doi: 10.1007/s11095-014-1360-0. Available at: http://dx.doi.org/10.1007/s11095-014-1360-0 (accessed 26/07/2014) [DOI] [PubMed] [Google Scholar]
- Jiang Z, Hong Z, Guo W, Xiaoyun G, Gengfa L, Yongning L, et al. A synthetic peptide derived from bactericidal/permeability-increasing protein neutralizes endotoxin in vitro and in vivo. Int Immunopharmacol. 2004;4:527–537. doi: 10.1016/j.intimp.2004.02.004. [DOI] [PubMed] [Google Scholar]
- Kang G, Kong PJ, Yuh YJ, Lim SY, Yim SV, Chun W, et al. Curcumin suppresses lipopolysaccharide-induced cyclooxygenase-2 expression by inhibiting activator protein 1 and nuclear factor kappaB bindings in BV2 microglial cells. J Pharmacol Sci. 2004;94:325–328. doi: 10.1254/jphs.94.325. [DOI] [PubMed] [Google Scholar]
- Kim J, Lee CM, Jeong HJ, Kim DW, Lee KY. Elevated anti-inflammatory effects of eicosapentaenoic acid based self-aggregated glycol chitosan nanoparticles. J Nanosci Nanotechnol. 2012;12:2672–2678. doi: 10.1166/jnn.2012.5685. [DOI] [PubMed] [Google Scholar]
- Kim TH, Ku S-K, Bae J-S. Anti-inflammatory activities of isorhamnetin-3-O-galactoside against HMGB1-induced inflammatory responses in both HUVECs and CLP-induced septic mice. J Cell Biochem. 2013a;114:336–345. doi: 10.1002/jcb.24361. [DOI] [PubMed] [Google Scholar]
- Kim TH, Ku S-K, Bae J-S. Persicarin is anti-inflammatory mediator against HMGB1-induced inflammatory responses in HUVECs and in CLP-induced sepsis mice. J Cell Physiol. 2013b;228:696–703. doi: 10.1002/jcp.24214. [DOI] [PubMed] [Google Scholar]
- Krasnodembskaya A, Samarani G, Song Y, Zhuo H, Su X, Lee JW, et al. Human mesenchymal stem cells reduce mortality and bacteremia in gram-negative sepsis in mice in part by enhancing the phagocytic activity of blood monocytes. Am J Physiol Lung Cell Mol Physiol. 2012;302:L1003–L1013. doi: 10.1152/ajplung.00180.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee S. Recognition properties of antibodies to PAMAM dendrimers and their use in immune detection of dendrimers. Biomed Microdevices. 2001;3:53–59. [Google Scholar]
- Lee S, Yang SC, Heffernan MJ, Taylor WR, Murthy N. Polyketal microparticles: a new delivery vehicle for superoxide dismutase. Bioconjug Chem. 2006;18:4–7. doi: 10.1021/bc060259s. [DOI] [PubMed] [Google Scholar]
- Lee SC. Biochemical and immunological properties of cytokines conjugated to dendritic polymers. Biomed Microdevices. 2004;6:191–202. doi: 10.1023/B:BMMD.0000042048.18186.ff. [DOI] [PubMed] [Google Scholar]
- Levy MM, Artigas A, Phillips GS, Rhodes A, Beale R, Osborn T, et al. Outcomes of the Surviving Sepsis Campaign in intensive care units in the USA and Europe: a prospective cohort study. Lancet Infect Dis. 2012;12:919–924. doi: 10.1016/S1473-3099(12)70239-6. [DOI] [PubMed] [Google Scholar]
- Liu D, Yang PS. Minocycline hydrochloride nanoliposomes inhibit the production of TNF-alpha in LPS-stimulated macrophages. Int J Nanomedicine. 2012;7:4769–4775. doi: 10.2147/IJN.S34036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu JJ, Song CW, Yue Y, Duan CG, Yang J, He T, et al. Quercetin inhibits LPS-induced delay in spontaneous apoptosis and activation of neutrophils. Inflamm Res. 2005;54:500–507. doi: 10.1007/s00011-005-1385-2. [DOI] [PubMed] [Google Scholar]
- Liu X, Zheng X, Wang N, Cao H, Lu Y, Long Y, et al. Kukoamine B, a novel dual inhibitor of LPS and CpG DNA, is a potential candidate for sepsis treatment. Br J Pharmacol. 2011a;162:1274–1290. doi: 10.1111/j.1476-5381.2010.01114.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu XR, Zheng XF, Ji SZ, Lv YH, Zheng DY, Xia ZF, et al. Metabolomic analysis of thermally injured and/or septic rats. Burns. 2010;36:992–998. doi: 10.1016/j.burns.2010.03.015. [DOI] [PubMed] [Google Scholar]
- Liu Y, Wu H, Nie Y-C, Chen J-L, Su W-W, Li P-B. Naringin attenuates acute lung injury in LPS-treated mice by inhibiting NF-κB pathway. Int Immunopharmacol. 2011b;11:1606–1612. doi: 10.1016/j.intimp.2011.05.022. [DOI] [PubMed] [Google Scholar]
- Lopez A, Lorente JA, Steingrub J, Bakker J, McLuckie A, Willatts S, et al. Multiple-center, randomized, placebo-controlled, double-blind study of the nitric oxide synthase inhibitor 546C88: effect on survival in patients with septic shock. Crit Care Med. 2004;32:21–30. doi: 10.1097/01.CCM.0000105581.01815.C6. [DOI] [PubMed] [Google Scholar]
- Ma P, Liu H-T, Wei P, Xu Q-S, Bai X-F, Du Y-G, et al. Chitosan oligosaccharides inhibit LPS-induced over-expression of IL-6 and TNF-α in RAW264.7 macrophage cells through blockade of mitogen-activated protein kinase (MAPK) and PI3K/Akt signaling pathways. Carbohydr Poly. 2011;84:1391–1398. [Google Scholar]
- Madrigal-Carballo S, Rodríguez G, Sibaja M, Reed JD, Vila AO, Molina F. Chitosomes loaded with cranberry proanthocyanidins attenuate the bacterial lipopolysaccharide-induced expression of iNOS and COX-2 in raw 264.7 macrophages. J Liposome Res. 2009;19:189–196. doi: 10.1080/08982100902729436. [DOI] [PubMed] [Google Scholar]
- Marik PE, Raghavan M. Stress-hyperglycemia, insulin and immunomodulation in sepsis. Intensive Care Med. 2004;30:748–756. doi: 10.1007/s00134-004-2167-y. [DOI] [PubMed] [Google Scholar]
- Marshall JC. Why have clinical trials in sepsis failed? Trends Mol Med. 2014;20:195–203. doi: 10.1016/j.molmed.2014.01.007. [DOI] [PubMed] [Google Scholar]
- Martire-Greco D, Landoni VI, Chiarella P, Rodriguez-Rodrigues N, Schierloh P, Rearte B, et al. all-trans-Retinoic acid improves immunocompetence in a murine model of lipopolysaccharide-induced immunosuppression. Clin Sci (Lond) 2014;126:355–365. doi: 10.1042/CS20130236. [DOI] [PubMed] [Google Scholar]
- Matthay MA. Severe sepsis – a new treatment with both anticoagulant and antiinflammatory properties. N Engl J Med. 2001;344:759–762. doi: 10.1056/NEJM200103083441009. [DOI] [PubMed] [Google Scholar]
- Matthay MA, Ware LB. Can nicotine treat sepsis? Nat Med. 2004;10:1161–1162. doi: 10.1038/nm1104-1161. [DOI] [PubMed] [Google Scholar]
- Mattsby-Baltzer I, Roseanu A, Motas C, Elverfors J, Engberg I, Hanson LA. Lactoferrin or a fragment thereof inhibits the endotoxin-induced interleukin-6 response in human monocytic cells. Pediatr Res. 1996;40:257–262. doi: 10.1203/00006450-199608000-00011. [DOI] [PubMed] [Google Scholar]
- Meisel C, Schefold JC, Pschowski R, Baumann T, Hetzger K, Gregor J, et al. Granulocyte-macrophage colony-stimulating factor to reverse sepsis-associated immunosuppression. Am J Respir Crit Care Med. 2009;180:640–648. doi: 10.1164/rccm.200903-0363OC. [DOI] [PubMed] [Google Scholar]
- Melamed A, Sorvillo FJ. The burden of sepsis-associated mortality in the United States from 1999 to 2005: an analysis of multiple-cause-of-death data. Crit Care. 2009;13:R28. doi: 10.1186/cc7733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mishra PR. An investigation on the approach to target lipopolysaccharide through polymeric capped nano-structured formulation for the management of sepsis. J Biomed Nanotechnol. 2011;7:47–49. doi: 10.1166/jbn.2011.1195. [DOI] [PubMed] [Google Scholar]
- Muenzer JT, Davis CG, Chang K, Schmidt RE, Dunne WM, Coopersmith CM, et al. Characterization and modulation of the immunosuppressive phase of sepsis. Infect Immun. 2010;78:1582–1592. doi: 10.1128/IAI.01213-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mullarkey M, Rose JR, Bristol J, Kawata T, Kimura A, Kobayashi S, et al. Inhibition of endotoxin response by e5564, a novel Toll-like receptor 4-directed endotoxin antagonist. J Pharmacol Exp Ther. 2003;304:1093–1102. doi: 10.1124/jpet.102.044487. [DOI] [PubMed] [Google Scholar]
- Novelli G, Morabito V, Ferretti G, Novelli S, Ruberto F, Pernice M, et al. Polymixin-B hemoperfusion in a case intolerant to conventional antibiotic therapy. Journal of Biomed Sci Eng. 2012;5:101–103. [Google Scholar]
- Opal SM, Patrozou E. Translational research in the development of novel sepsis therapeutics: logical deductive reasoning or mission impossible? Crit Care Med. 2009;37:S10–S15. doi: 10.1097/CCM.0b013e3181921497. [DOI] [PubMed] [Google Scholar]
- Opal SM, Laterre P, Francois B, La Rosa SP, Angus DC, Mira J, et al. Effect of eritoran, an antagonist of md2-tlr4, on mortality in patients with severe sepsis: the access randomized trial. JAMA. 2013;309:1154–1162. doi: 10.1001/jama.2013.2194. [DOI] [PubMed] [Google Scholar]
- Park K. Facing the truth about nanotechnology in drug delivery. ACS Nano. 2013;7:7442–7447. doi: 10.1021/nn404501g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pawson AJ, Sharman JL, Benson HE, Faccenda E, Alexander SP, Buneman OP, Davenport AP, McGrath JC, Peters JA, Southan C, Spedding M, Yu W, Harmar AJ NC-IUPHAR. The IUPHAR/BPS Guide to PHARMACOLOGY: an expert-driven knowledge base of drug targets and their ligands. Nucl Acids Res. 2014;42(Database Issue):D1098–1106. doi: 10.1093/nar/gkt1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peer D, Karp JM, Hong S, Farokhzad OC, Margalit R, Langer R. Nanocarriers as an emerging platform for cancer therapy. Nat Nano. 2007;2:751–760. doi: 10.1038/nnano.2007.387. [DOI] [PubMed] [Google Scholar]
- Pereira V, Serpa Neto A, Cardoso S, Manetta J, Esposito D, de Oliveira Prado Pasqualucci M, et al. Association between use of lung-protective ventilation with lower tidal volumes and risk of acute lung injury, mortality, pulmonary infection, and atelectasis: a meta-analysis. Critical care. 2013;17(Suppl. 3):P38. [Google Scholar]
- van der Poll T, Opal SM. Host-pathogen interactions in sepsis. Lancet Infect Dis. 2008;8:32–43. doi: 10.1016/S1473-3099(07)70265-7. [DOI] [PubMed] [Google Scholar]
- Ponnappa BC, Dey I, Tu G-C, Zhou F, Aini M, Cao Q-N, et al. In vivo delivery of antisense oligonucleotides in pH-sensitive liposomes inhibits lipopolysaccharide-induced production of tumor necrosis factor-α in rats. J Pharmacol Exp Ther. 2001;297:1129–1136. [PubMed] [Google Scholar]
- Puddu P, Latorre D, Valenti P, Gessani S. Immunoregulatory role of lactoferrin-lipopolysaccharide interactions. Biometals. 2010;23:387–397. doi: 10.1007/s10534-010-9307-3. [DOI] [PubMed] [Google Scholar]
- Qiao Y, Bai XF, Du YG. Chitosan oligosaccharides protect mice from LPS challenge by attenuation of inflammation and oxidative stress. Int Immunopharmacol. 2011;11:121–127. doi: 10.1016/j.intimp.2010.10.016. [DOI] [PubMed] [Google Scholar]
- Radnai B, Tucsek Z, Bognar Z, Antus C, Mark L, Berente Z, et al. Ferulaldehyde, a water-soluble degradation product of polyphenols, inhibits the lipopolysaccharide-induced inflammatory response in mice. J Nutr. 2009;139:291–297. doi: 10.3945/jn.108.097386. [DOI] [PubMed] [Google Scholar]
- von Ranke FM, Zanetti G, Hochhegger B, Marchiori E. Infectious diseases causing diffuse alveolar hemorrhage in immunocompetent patients: a state-of-the-art review. Lung. 2013;191:9–18. doi: 10.1007/s00408-012-9431-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rearte B, Maglioco A, Balboa L, Bruzzo J, Landoni VI, Laborde EA, et al. Mifepristone (RU486) restores humoral and T cell-mediated immune response in endotoxin immunosuppressed mice. Clin Exp Immunol. 2010;162:568–577. doi: 10.1111/j.1365-2249.2010.04262.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rearte B, Maglioco A, Machuca D, Greco DM, Landoni VI, Rodriguez-Rodrigues N, et al. Dehydroepiandrosterone and metyrapone partially restore the adaptive humoral and cellular immune response in endotoxin immunosuppressed mice. Innate Immunity. 2013;20:585–597. doi: 10.1177/1753425913502243. [DOI] [PubMed] [Google Scholar]
- Remick DG. Pathophysiology of sepsis. Am J Pathol. 2007;170:1435–1444. doi: 10.2353/ajpath.2007.060872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riedemann NC, Guo R-F, Ward PA. The enigma of sepsis. J Clin Invest. 2003;112:460–467. doi: 10.1172/JCI19523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riedemann NC, Guo RF, Gao H, Sun L, Hoesel M, Hollmann TJ, et al. Regulatory role of C5a on macrophage migration inhibitory factor release from neutrophils. J Immunol. 2004;173:1355–1359. doi: 10.4049/jimmunol.173.2.1355. [DOI] [PubMed] [Google Scholar]
- Rittirsch D, Flierl MA, Ward PA. Harmful molecular mechanisms in sepsis. Nat Rev Immunol. 2008;8:776–787. doi: 10.1038/nri2402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salvesen GS, Riedl SJ. Caspase mechanisms. Adv Exp Med Biol. 2008;615:13–23. doi: 10.1007/978-1-4020-6554-5_2. [DOI] [PubMed] [Google Scholar]
- Sha T, Sunamoto M, Kitazaki T, Sato J, Ii M, Iizawa Y. Therapeutic effects of TAK-242, a novel selective Toll-like receptor 4 signal transduction inhibitor, in mouse endotoxin shock model. Eur J Pharmacol. 2007;571:231–239. doi: 10.1016/j.ejphar.2007.06.027. [DOI] [PubMed] [Google Scholar]
- Shai Y, Makovitzky A, Avrahami D. Host defense peptides and lipopeptides: modes of action and potential candidates for the treatment of bacterial and fungal infections. Curr Protein Peptide Sci. 2006;7:479–486. doi: 10.2174/138920306779025620. [DOI] [PubMed] [Google Scholar]
- Shao B, Munford RS, Kitchens R, Varley AW. Hepatic uptake and deacylation of the LPS in bloodborne LPS-lipoprotein complexes. Innate Immun. 2012;18:825–833. doi: 10.1177/1753425912442431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shukla P, Srivastava AK. Different fabrication processes and polymer blends for sustained release matrix tablets of highly water soluble drug (metoprolol succinate): an in vitro evaluation. Inventi Impact: Pharm Tech. 2012 Available at: http://www.inventi.in/Article/ppt/463/12.aspx (accessed 9/3/2014) [Google Scholar]
- Shukla P, Gupta G, Singodia D, Shukla R, Verma AK, Dwivedi P, et al. Emerging trend in nano-engineered polyelectrolyte-based surrogate carriers for delivery of bioactives. Expert Opin Drug Deliv. 2010;7:993–1011. doi: 10.1517/17425247.2010.510830. [DOI] [PubMed] [Google Scholar]
- Singh AK, Jiang Y, Gupta S, Younus M, Ramzan M. Anti-inflammatory potency of nano-formulated puerarin and curcumin in rats subjected to the lipopolysaccharide-induced inflammation. J Med Food. 2013;16:899–911. doi: 10.1089/jmf.2012.0049. [DOI] [PubMed] [Google Scholar]
- Singodia D, Kansal S, Verma A, Shukla R, Shukla P, Dwivedi P, et al. Development of nevirapine loaded novel surfactant free polymeric emulsion and investigations for its suitability as drug delivery vehicle. J Bionanosci. 2010;4:66–73. [Google Scholar]
- Song Y, Dou H, Gong W, Liu X, Yu Z, Li E, et al. Bis-N-norgliovictin, a small-molecule compound from marine fungus, inhibits LPS-induced inflammation in macrophages and improves survival in sepsis. Eur J Pharmacol. 2013;705:49–60. doi: 10.1016/j.ejphar.2013.02.008. [DOI] [PubMed] [Google Scholar]
- Stefan M, Melnig V, Pricop D, Neagu A, Mihasan M, Tartau L, et al. Attenuated effects of chitosan-capped gold nanoparticles on LPS-induced toxicity in laboratory rats. Mater Sci Eng. 2013;33:550–556. doi: 10.1016/j.msec.2012.09.031. [DOI] [PubMed] [Google Scholar]
- Strachan AJ, Woodruff TM, Haaima G, Fairlie DP, Taylor SM. A new small molecule C5a receptor antagonist inhibits the reverse-passive Arthus reaction and endotoxic shock in rats. J Immunol. 2000;164:6560–6565. doi: 10.4049/jimmunol.164.12.6560. [DOI] [PubMed] [Google Scholar]
- Stringer KA, Serkova NJ, Karnovsky A, Guire K, Paine R, 3rd, Standiford TJ. Metabolic consequences of sepsis-induced acute lung injury revealed by plasma (1)H-nuclear magnetic resonance quantitative metabolomics and computational analysis. Am J Physiol Lung Cell Mol Physiol. 2011;300:L4–L11. doi: 10.1152/ajplung.00231.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun D, Zhuang X, Xiang X, Liu Y, Zhang S, Liu C, et al. A novel nanoparticle drug delivery system: the anti-inflammatory activity of curcumin is enhanced when encapsulated in exosomes. Molecular Therapy. 2010;18:1606–1614. doi: 10.1038/mt.2010.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suntres ZE, Shek PN. Prophylaxis against lipopolysaccharide-induced lung injuries by liposome-entrapped dexamethasone in rats. Biochem Pharmacol. 2000;59:1155–1161. doi: 10.1016/s0006-2952(99)00411-6. [DOI] [PubMed] [Google Scholar]
- Tang D, Kang R, Xiao W, Zhang H, Lotze MT, Wang H, et al. Quercetin prevents LPS-induced high-mobility group box 1 release and proinflammatory function. Am J Respir Cell Mol Biol. 2009;41:651–660. doi: 10.1165/rcmb.2008-0119OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang Y, Lv B, Wang H, Xiao X, Zuo X. PACAP inhibit the release and cytokine activity of HMGB1 and improve the survival during lethal endotoxemia. Int Immunopharmacol. 2008;8:1646–1651. doi: 10.1016/j.intimp.2008.07.014. [DOI] [PubMed] [Google Scholar]
- The Lancet Infectious Diseases. For sepsis, the drugs don't work. Lancet Infect Dis. 2012;12:89. doi: 10.1016/S1473-3099(12)70020-8. [DOI] [PubMed] [Google Scholar]
- Thierry Calandra BE, Le Roy D, Pugin J, Metz CN, Hültner L, Heumann D, et al. Protection from septic shock by neutralization of macrophage migration inhibitory factor. Pediatr Infect Dis J. 2001;20:470–471. doi: 10.1038/72262. [DOI] [PubMed] [Google Scholar]
- Tohyama S, Onodera S, Tohyama H, Yasuda K, Nishihira J, Mizue Y, et al. A novel DNA vaccine-targeting macrophage migration inhibitory factor improves the survival of mice with sepsis. Gene Ther. 2008;15:1513–1522. doi: 10.1038/gt.2008.112. [DOI] [PubMed] [Google Scholar]
- Toussaint S, Gerlach H. Activated protein C for sepsis. N Engl J Med. 2009;361:2646–2652. doi: 10.1056/NEJMct0808063. [DOI] [PubMed] [Google Scholar]
- Tse MT. Trial watch: sepsis study failure highlights need for trial design rethink. Nat Rev Drug Discov. 2013;12:334–334. doi: 10.1038/nrd4016. [DOI] [PubMed] [Google Scholar]
- Tsoyi K, Jang HJ, Kim JW, Chang HK, Lee YS, Pae HO, et al. Stimulation of alpha7 nicotinic acetylcholine receptor by nicotine attenuates inflammatory response in macrophages and improves survival in experimental model of sepsis through heme oxygenase-1 induction. Antioxid Redox Signal. 2011;14:2057–2070. doi: 10.1089/ars.2010.3555. [DOI] [PubMed] [Google Scholar]
- Ulloa L, Ochani M, Yang H, Tanovic M, Halperin D, Yang R, et al. Ethyl pyruvate prevents lethality in mice with established lethal sepsis and systemic inflammation. Proc Natl Acad Sci U S A. 2002;99:12351–12356. doi: 10.1073/pnas.192222999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urban C, Zychlinsky A. Netting bacteria in sepsis. Nat Med. 2007;13:403–404. doi: 10.1038/nm0407-403. [DOI] [PubMed] [Google Scholar]
- Vallespi MG, Alvarez-Obregon JC, Rodriguez-Alonso I, Montero T, Garay H, Reyes O, et al. A Limulus anti-LPS factor-derived peptide modulates cytokine gene expression and promotes resolution of bacterial acute infection in mice. Int Immunopharmacol. 2003;3:247–256. doi: 10.1016/S1567-5769(02)00277-1. [DOI] [PubMed] [Google Scholar]
- Veronese FM, Caliceti P, Schiavon O, Sergi M. Polyethylene glycol-superoxide dismutase, a conjugate in search of exploitation. Adv Drug Deliv Rev. 2002;54:587–606. doi: 10.1016/s0169-409x(02)00029-7. [DOI] [PubMed] [Google Scholar]
- Vicent MJ, Cascales L, Carbajo RJ, Cortes N, Messeguer A, Perez Paya E. Nanoconjugates as intracorporeal neutralizers of bacterial endotoxins. J Control Release. 2010;142:277–285. doi: 10.1016/j.jconrel.2009.10.026. [DOI] [PubMed] [Google Scholar]
- Wang W, Luo M, Fu Y, Wang S, Efferth T, Zu Y. Glycyrrhizic acid nanoparticles inhibit LPS-induced inflammatory mediators in 264.7 mouse macrophages compared with unprocessed glycyrrhizic acid. Int J Nanomedicine. 2013;8:1377–1383. doi: 10.2147/IJN.S37788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wannemuehler TJ, Manukyan MC, Brewster BD, Rouch J, Poynter JA, Wang Y, et al. Advances in mesenchymal stem cell research in sepsis. J Surg Res. 2012;173:113–126. doi: 10.1016/j.jss.2011.09.053. [DOI] [PubMed] [Google Scholar]
- Warren HS, Suffredini AF, Eichacker PQ, Munford RS. Risks and benefits of activated protein C treatment for severe sepsis. N Engl J Med. 2002;347:1027–1030. doi: 10.1056/NEJMsb020574. [DOI] [PubMed] [Google Scholar]
- Weber P, Wang P, Maddens S, Wang P, Wu R, Miksa M, et al. VX-166: a novel potent small molecule caspase inhibitor as a potential therapy for sepsis. Critical Care. 2009;13:R146. doi: 10.1186/cc8041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu J, Zhou L, Liu J, Ma G, Kou Q, He Z, et al. The efficacy of thymosin alpha 1 for severe sepsis (ETASS): a multicenter, single-blind, randomized and controlled trial. Critical Care. 2013;17:R8. doi: 10.1186/cc11932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang M, Cao L, Xie M, Yu Y, Kang R, Yang L, et al. Chloroquine inhibits HMGB1 inflammatory signaling and protects mice from lethal sepsis. Biochem Pharmacol. 2013;86:410–418. doi: 10.1016/j.bcp.2013.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang R, Gallo DJ, Baust JJ, Uchiyama T, Watkins SK, Delude RL, et al. Ethyl pyruvate modulates inflammatory gene expression in mice subjected to hemorrhagic shock. Am J Physiol Gastrointest Liver Physiol. 2002;283:G212–G221. doi: 10.1152/ajpgi.00022.2002. [DOI] [PubMed] [Google Scholar]
- Yang SC, Bhide M, Crispe IN, Pierce RH, Murthy N. Polyketal copolymers: a new acid-sensitive delivery vehicle for treating acute inflammatory diseases. Bioconjug Chem. 2008;19:1164–1169. doi: 10.1021/bc700442g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yermak IM, Davidova VN, Gorbach VI, Luk'yanov PA, Solov'eva TF, Ulmer AJ, et al. Forming and immunological properties of some lipopolysaccharide-chitosan complexes. Biochimie. 2006;88:23–30. doi: 10.1016/j.biochi.2005.07.004. [DOI] [PubMed] [Google Scholar]
- Yibin G, Jiang Z, Hong Z, Gengfa L, Liangxi W, Guo W, et al. A synthesized cationic tetradecapeptide from hornet venom kills bacteria and neutralizes lipopolysaccharide in vivo and in vitro. Biochem Pharmacol. 2005;70:209–219. doi: 10.1016/j.bcp.2005.04.040. [DOI] [PubMed] [Google Scholar]
- Yin JP, Su ZL, Wang YM, Wang T, Tian SS, Xu XX, et al. Release of HMGB1 by LPS-treated cardiac fibroblasts and its contribution to the production of collagen type I and III] Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2012;28:785–788. [PubMed] [Google Scholar]
- Yuan Z, Syed MA, Panchal D, Rogers D, Joo M, Sadikot RT. Curcumin mediated epigenetic modulation inhibits TREM-1 expression in response to lipopolysaccharide. Int J Biochem Cell Biol. 2012;44:2032–2043. doi: 10.1016/j.biocel.2012.08.001. [DOI] [PubMed] [Google Scholar]
- Zaborin A, Defazio JR, Kade M, Kaiser BLD, Belogortseva N, Camp DG, et al. Phosphate-containing polyethylene glycol polymers prevent lethal sepsis from multidrug-resistant pathogens. Antimicrob Agents Chemother. 2014;58:966–977. doi: 10.1128/AAC.02183-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng X, Yang D, Liu X, Wang N, Li B, Cao H, et al. Identification of a new anti-LPS agent, geniposide, from Gardenia jasminoides Ellis, and its ability of direct binding and neutralization of lipopolysaccharide in vitro and in vivo. Int Immunopharmacol. 2010;10:1209–1219. doi: 10.1016/j.intimp.2010.07.001. [DOI] [PubMed] [Google Scholar]