
Figure 7. cAMP stimulated adiponectin secretion is Epac dependent and potentiated by Ca2+.
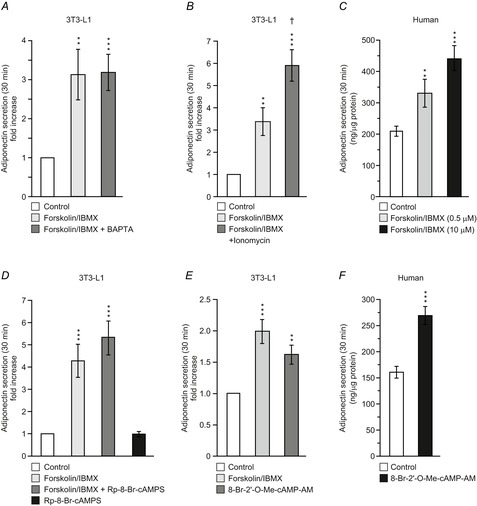
A, adiponectin secretion in 3T3-L1 adipocytes expressed as fold-increase compared to basal (5 mm glucose alone) during 30 min incubations (32°C) with forskolin (10 μm) + IBMX (200 μm) with or without BAPTA pre-treatment (50 μm). B, as in A, but cells incubated for 30 min with forsk–IBMX in the absence or presence of ionomycin (1 μm). C, adiponectin secretion during 30 min incubation of human primary subcutaneous adipocytes with forskolin (0.5 or 10 μm) together with IBMX (200 μm). D, effects of the cAMP antagonist Rp-8-Br-cAMPS (200 μm) on forsk–IBMX-stimulated adiponectin secretion in 3T3-L1 adipocytes. E, 3T3-L1 adipocyte adiponectin secretion stimulated during 30 min incubation with the Epac agonist 8-Br-2′-O-Me-cAMP-AM (20 μm). F, adiponectin secretion in human subcutaneous adipocytes stimulated with 8-Br-2′-O-Me-cAMP-AM (20 μm) for 30 min. Note that basal secretion (control) is somewhat lower than in C, but that the fold-increase produced by the agonist is of similar magnitude as that stimulated with forsk–IBMX. Data are mean values ± SEM of 14 (A), 8 (B), 10 (D) and 11 (E) experiments. Isolated adipocytes from four and three patients were used in C and F, respectively. **P < 0.01; ***P < 0.001.