

Abstract
Objective
To characterize the diversity and taxonomic relative abundance of the gut microbiota in patients with never-treated, recent-onset psoriatic arthritis (PsA).
Methods
High-throughput 16S rRNA pyrosequencing was utilized to compare community composition of gut microbiota in PsA patients (n=16), subjects with psoriasis of the skin (Ps) (n=15) and healthy, matched-controls (n=17). Samples were further assessed for the presence and levels of fecal and serum secretory immunoglobulin A (sIgA), pro-inflammatory proteins and fatty-acids.
Results
The gut microbiota observed in PsA and Ps patients was less diverse when compared to healthy controls. These could be attributed to the reduced presence of several taxa. While both groups showed a relative decrease in Coprococcus spp., PsA samples were characterized by a significant reduction in Akkermansia, Ruminococcus, and Pseudobutyrivibrio. Supernatants of fecal samples from PsA patients revealed an increase in sIgA and a decrease in receptor activator of nuclear factor kappa-B ligand (RANKL) levels. Fatty acid analysis revealed low levels of hexanoate and heptanoate in PsA and Ps patients.
Conclusion
PsA and Ps patients had a lower relative abundance of multiple intestinal bacteria. Although some genera were concomitantly decreased in both conditions, PsA samples had lower abundance of reportedly beneficial taxa. This gut microbiota profile in PsA was similar to that published for patients with IBD and was associated with changes in specific inflammatory proteins unique to this group, and distinct from Ps and controls. Thus, the role of gut microbiota in the continuum of Ps-PsA pathogenesis and the associated immune response merits further study.
Keywords: psoriatic arthritis, spondyloarthropathies, RANKL, microbiome, medium-chain fatty acids, hexanoate, heptanoate, inflammatory bowel disease, psoriasis
INTRODUCTION
Psoriatic arthritis (PsA) is a type of chronic spondyloarthritis (SpA), typically affecting individuals with pre-existing psoriasis of the skin (Ps). Despite recent advances in diagnosis and treatment, the pathogenesis of PsA remains unclear. The prevalent paradigm posits that in the presence of predisposing genetic factors (e.g., HLA-B*27, Cw6), individuals with Ps will develop PsA after exposure to yet-unidentified environmental factors (1,2). Previously proposed triggers include viruses, bacterial infections, trauma and stress. Interest has recently reemerged concerning the role of the gut microbiome (the totality of bacteria and their genes in a given biological niche) and associated gut inflammation in the pathogenesis of the SpA disease spectrum (3–6). HLA-B27 over-expressing rats, for instance, develop arthritis and colitis only in the presence of specific intestinal microbes (7). Similarly, SKG mice develop joint inflammation, enthesitis, skin inflammation and ileitis after injection with β-glucan, a major component of bacterial and fungal cell walls (8).
Strong epidemiologic evidence also suggests an intimate relationship between intestinal and joint inflammation in SpA. Patients with PsA, Ps and ankylosing spondylitis (AS) experience a much higher incidence of inflammatory bowel disease (IBD). Furthermore, articular manifestations are found in more than one-third of patients with known Crohn’s disease or ulcerative colitis (UC) (9). This has led to the consideration of IBD-related arthritis as part of the SpA spectrum.
Several studies have further associated intestinal mucosal inflammation and human SpA. Approximately 70% of all SpA patients show at least some ileocolonoscopic or histologic alterations in the intestinal epithelium. PsA has specifically been shown to be associated with both subclinical gut inflammation (10) and a significantly increased risk of subsequent Crohn’s disease (11).
Therefore, several animal and human studies substantiate the hypothesis pointing toward a biological link between (local) gut and (systemic) joint inflammation. This suggests a common etiology, but its precise nature remains unknown.
Among sites of exposure to bacterial antigens, the intestinal mucosa represents a unique environment for triggering of local and distal autoimmunity. The human intestinal microbiome contains roughly 100 trillion cells whose genomes encode ~3.3 million protein-coding genes (100-fold more than the human genome). The NIH Human Microbiome Project was recently launched to better understand and define this collective human-microbiome “supraorganism” in both health and disease.
Utilizing novel high-throughput DNA sequencing, it is now possible to identify bacteria in a given community, including unculturable or fastidious organisms, without the need for conventional microbiology techniques. Our study aimed to describe, for the first time, potential alterations in gut microbiota composition of patients with PsA and associated local inflammatory response, compared to Ps and healthy controls.
METHODS
Study participants
Consecutive patients from rheumatology clinics and practice offices of New York University School of Medicine were screened for the presence of PsA [samples and sequences obtained from previously described cohort (12)] or psoriasis of the skin (Ps). After informed consent was signed, each patient’s medical history and medications were determined. A screening musculoskeletal examination, laboratory and radiographic assessments were also performed or reviewed. All PsA/Ps patients who met study criteria were offered enrollment. Non-arthritic healthy subjects were also identified from a recently published study (12) and enrolled as controls.
This study was approved by the Institutional Review Board of NYU School of Medicine, and written informed consent was obtained from all study participants.
Inclusion and exclusion criteria
Patients were included as recent-onset PsA if they met Classification Criteria for Psoriatic Arthritis (CASPAR) including presence of current active psoriasis of the skin (Ps) and arthritis, and had never been treated with systemic disease-modifying anti-rheumatic drugs (DMARDs; oral and/or biologic agents) or steroids. Patients were included as Ps without PsA if they were diagnosed by a dermatologist and had no arthritis, enthesitis or dactylitis (as assessed by a rheumatologist) at enrollment. Healthy controls were age-, sex-, and ethnicity-matched individuals with no personal history of psoriasis, autoimmune disease (including IBD) or inflammatory arthritis. Criteria for inclusion required that all subjects be age 18 years or older.
Exclusion criteria applied to all groups were as follows: recent (<3 months prior) use of any antibiotic therapy, current extreme diet (e.g., parenteral nutrition or macrobiotic diet), known history of malignancy or IBD, current consumption of probiotics, or any gastrointestinal tract surgery leaving permanent residua (e.g., gastrectomy, bariatric surgery, or colectomy).
Sample collection and DNA extraction
Fecal samples were obtained for all participants within 24 hours of production. DNA extraction, amplification of the V1–V2 16S rRNA gene region and 454 pyrosequencing were performed for all samples as recently published (13).
Sequence analysis
Sequencing data was compiled and processed using mothur software and converted to standard Fasta format as described in prior studies (13). Briefly, sequences were grouped into operational taxonomic units (OTUs) using the average neighbor algorithm. Sequences with a distance-based similarity of ≥97% were assigned to the same OTU. For each sample, microbial diversity was estimated by calculating the Shannon diversity index or Faith’s phylodiversity index. Phylogenetic classification was performed using the Bayesian classifier algorithm, with a boot-strap cutoff value of 60%. For microbiota comparison between samples, only 2718 sequences (number of high-quality sequences obtained from the sample with lowest counts) were used for all data analyses.
For UniFrac analysis, a phylogenetic tree was inferred using clearcut (14), on the 16S rRNA sequence alignment generated by mothur. Unweighted UniFrac was run using the resulting tree. Principal Coordinate of Analysis (PCoA) was performed on the resulting matrix of distances between each pair of samples.
Serum and fecal measurement of sIgA, proteins and cytokines
Protein concentration in feces was measured by the BCA method. ELISA assays were performed to determine serum and fecal concentrations of secretory IgA (sIgA; Immundiagnostik AG), receptor activator of nuclear factor kappa-B ligand (RANKL, Immundiagnostik AG), osteoprotegerin (OPG; Raybio), S-100A12 protein (S100, Circulex) utilizing a validated protocol (15).
Human leukocyte antigen (HLA) allele determination
Genomic DNA was isolated from peripheral blood of PsA patients using QIAamp Blood Mini Kit (Qiagen). HLA-B and C alleles were determined by Single Specific Primer- Polymerase Chain Reaction (SSP-PCR) methodologies (Weatherall Institute for Molecular Medicine, Oxford, UK) (12).
Fecal measurement of fatty acids (FAs)
Levels of fecal short-chain fatty acids (SCFAs) and medium-chain fatty acids (MCFAs) were measured by gas chromatography-mass spectrometry (GC-MS) at the University of Michigan Metabolomics Core. Samples were extracted by aqueous extraction solvent, homogenized and centrifuged. GC (Agilent 6890) separation was performed using a ZB-Wax plus column (Phenomenex). A single quadrupole mass spectrometer (Agilent, 5973 inert MSD) was used to identify and quantitate the SCFAs, using Agilent Chemstation software. The acquired GC-MS data were processed by Agilent mass hunter quantitative analysis software (version B.06). Retention times and characteristic masses of FAs were used for peak identification and quantification. Absolute quantities of FAs were normalized to the sample mass.
Statistical analysis
In order to identify differentially abundant bacterial taxa among groups, we applied the LefSe analytical method (16). In brief, LefSe [linear discriminant analysis (LDA) coupled with effect size measurements] is a metagenomic biomarker-discovery approach based on an algorithm that first performs a nonparametric Kruskal-Wallis test in order to identify bacterial taxa whose relative abundance is significantly different in a group of interest (e.g., PsA) compared to controls (i.e., healthy or Ps). Subsequently, LefSe applies LDA to those bacterial taxa identified as significantly different (P < 0.05) and further assesses the effect size of each differentially abundant taxon (16). Only those taxa that obtain a log LDA score >2 are ultimately considered. As a result, LefSe indicates those taxa and OTUs that better discriminate between phenotypes. In addition, since LefSe does not consider multiple hypothesis testing, we further applied the non-parametric Wilcoxon test for every taxon at every phylogenetic level whose average abundance was higher than 0.1% (in any of the three groups analyzed). We then applied the Benjamini and Hochberg false discovery rate test (FDR). Those bacterial taxa/OTUs with P < 0.05 and FDR q value lower than 0.2 were considered as the main bacterial taxa differentiating between groups of samples.
For cross-sectional analyses of baseline characteristics and comparison of diversity indexes between groups, differences were evaluated using Student’s t-test, Mann-Whitney U test, one-way ANOVA, AMOVA or chi-square test, as appropriate. Two-tailed tests were used for significance testing, and P values less than 0.05 were considered significant.
Correlation and network analyses
Spearman correlation between taxa/OTUs, FAs, sIgA and inflammatory proteins was performed using the statistical R package command cor.test. Correlations were performed only in those taxa/OTUs found to be statistically significant between groups by LefSe. P values under 0.05 were considered significant.
The Optimal Bayesian network structure was inferred through ‘high climbing’ algorithm implemented in the bnlearn R package (17). Regularized inference was carried out by rejecting those relations between nodes with an associated Spearman correlation p-value greater than 0.05.
RESULTS
Patients
Only patients with recent-onset, DMARD-naïve PsA were included in the study; 56% were female and mean age was 46.2 years (Table 1). Mean disease duration was 0.8 months and no patient had ever received steroids, oral DMARDs, or biologic drugs. All patients had active skin psoriatic lesions and clinical or radiographic evidence of arthritis at enrollment (25% presented with axial arthritis). Ps and healthy controls were age-, sex-, and ethnicity-matched to PsA subjects. Baseline characteristics are described in Table 1.
Table 1.
Demographic and clinical data among patients with recent-onset psoriatic arthritis (PsA), psoriasis of the skin (Ps) and healthy control participants.
| Characteristic | PsA (n=16) | Ps (n=15) | Healthy Controls (n=17) |
|---|---|---|---|
| Age, years, mean (median) | 46.2 (40) | 39.4 (37) | 42.2 (39) |
| Female, % | 56% | 53% | 64% |
| Ethnicity, white* | 62% | 66% | 59% |
| HLA-B27, % | 12% | n/a | n/a |
| HLA- Cw6, % | 18% | n/a | n/a |
| HLA-B27 and/or Cw6, % | 30% | n/a | n/a |
| Disease duration, months, mean (median) | 0.8 (0) | 16 (11) | n/a |
| Disease activity parameters (articular) | |||
| CRP, mg/l, mean (median) | 7.5 (0) | 1 (0) | 0 |
| DAS28, mean (median) | 4.8 (4.7) | n/a | n/a |
| Patient VAS pain, mm, mean (median) | 50.6 (45) | n/a | n/a |
| Active Joint Count, mean (median) | 4.7 (3) | n/a | n/a |
| Axial involvement, % | 25% | n/a | n/a |
| Disease activity parameters (skin) | |||
| PASI, mean (median) | 5.2 (3.8) | 6.3 (4.3) | n/a |
| Nail psoriasis, % | 75% | 69% | n/a |
| Medication use | |||
| NSAIDs, current, % | 75% | 0 | n/a |
| Methotrexate, % | 6%** | 0 | n/a |
| Prednisone, % | 0 | 0 | n/a |
| Biological agent, % | 0 | 0 | n/a |
Abbreviations: PsA, psoriatic arthritis; Ps, psoriasis of the skin only; CRP, C-reactive protein; DAS28, Disease Activity Score with 28 joint count; VAS, visual analog scale; PASI, Psoriasis Area and Severity Index; NSAIDs, non-steroidal anti-inflammatory drugs.
Including Hispanic whites.
One patient had received one dose of methotrexate the week prior to enrollment.
Decreased diversity in PsA and Ps gut microbiota
A total of 48 fecal samples were obtained from PsA, Ps and healthy subjects for sequencing. Using a distance-based similarity of ≥97% for operational taxonomic units (OTU) assignment, a total of 2835 OTUs were identified. When compared to healthy subjects, microbial diversity was significantly reduced in PsA and Ps samples, as calculated by the Shannon diversity index and Faith’s phylodiversity index (Supplementary Figures 1A, B). Subsequently, we analyzed whether the overall structure of the microbiota of healthy samples differed from that of Ps and PsA and quantified the similarity by applying the UniFrac phylogenetic distance. We further applied PCoA to cluster samples along orthogonal axes of maximal variance. As shown in Supplementary Figure 1C, PC1 axes discriminated most healthy samples from the majority of Ps and PsA samples. Moreover, analysis of molecular variance (AMOVA) of the obtained UniFrac distances between samples revealed that overall microbiota structure was also significantly different when comparing PsA to Ps samples (Supplementary Figure 1).
Lower relative abundance of Akkermansia and Ruminoccocus is characteristic of PsA gut microbiota
To further investigate which bacterial taxa were distinct among groups, LefSe analysis was applied (see Methods). Interestingly, while no bacterial taxa were found to be enriched in PsA patients, relative abundance of several microbial clades were decreased in both PsA and Ps, and therefore enriched in healthy controls (Figure 1 and Supplementary Figure 2). Within these identified components of the intestinal microbiota, Akkermansia, Ruminococcus and Pseudobutyrivibrio were considered the most relevant genera that discriminated PsA microbiota from healthy controls (Figure 1A, D). At other levels of taxonomic classification, unclassified Clostridia and the parental taxonomic levels of Akkermansia (Verrucomicrobia, Verrucomicrobiae and Verrucomicrobiales) were also significantly decreased in PsA. The Ps gut microbiota was characterized by a reduced relative abundance of the genera Parabacteroides and Coprobacillus (Figure 1B, D). The comparison between PsA and Ps groups revealed that the higher taxonomic levels for Akkermansia and Ruminoccocus (including Firmicutes/Clostridiales and Verrucomicrobiales, respectively) were significantly less abundant in PsA patients (Figure 1C, D), while Bacteroidetes phylum and Coprobacillus genus were less abundant in Ps samples. Akkermansia and Ruminoccocus per se were also relatively decreased in PsA (Figure 1C, Supplementary Figure 2).
Figure 1. Bacterial taxa differences between PsA, Ps and healthy controls.

Bacterial taxa significantly enriched in healthy controls (HLT) as compared to psoriatic arthritis (PsA) patients were detected by LefSe (P <0.05, LDA >2logs) and after false-discovery rate (FDR). (A) Overall, no taxon was found to be enriched in PsA (red bars) compared to HLT (blue bars) or (B) Ps patients (green bars). Taxa are arranged in descending order according to their LDA score and marked with an asterisk (*) when significance remained after FDR correction. (C) Taxa differentiating PsA from Ps samples. (D) Box-plots with relative abundance (parts per unit) of specific taxa underrepresented in PsA and Ps subjects. Only those with statistical differences after FDR correction are shown.
Our ability to analyze the microbiota beyond the genus level allowed us to investigate the various OTUs that were underrepresented in patients with PsA. Several OTUs had a decreased relative abundance compared to healthy controls, including OTUs 43 (Coprococcus), 31 (Pseudobutyrivibrio), 26 (Parabacteroides), 83 (unclassified_Ruminococcaceae), 65 (Alistipes), and 85 (Akkermansia) (Figure 2A, 2D and Supplementary Figure 3). Intriguingly, several of these OTUs, including OTU43 as well as OTUs 26, 83, and 35 were also decreased in Ps patients, suggesting a possible common gut microbiota signature for Ps and PsA (Figure 2 and Supplementary Figure 3). Moreover, Ps samples showed a significantly decreased relative abundance of several other OTUs when compared to healthy subjects, including OTUs 44, 89 and 106 (Figure 2B, 2D). OTU11 was the only overrepresented OTU in the Ps group. When comparing PsA and Ps groups, patients with skin disease only had increased relative abundance of OTUs 11, 16 and 32, while patients with PsA had an overrepresentation of OTU44 only (Figure 2C, Supplementary Figure 3).
Figure 2. Decreased operational taxonomic units (OTUs) in PsA and Ps gut microbiota.
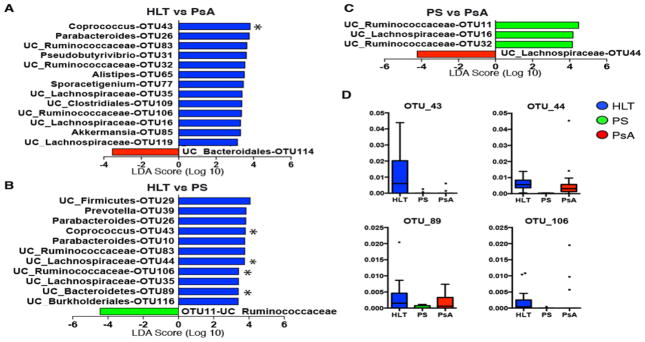
Several members of the gut microbial communities are underrepresented in PsA and Ps patients at the OTU level. (A) Bacterial OTUs significantly enriched in healthy controls (blue bars) as compared to PsA (red bars) and (B) Ps patients (green bars) were detected by LefSe. Those significant OTUs with q<0.2 after false-discovery rate (FDR) analysis are indicated by asterisks (*). (C) OTUs with distinct abundance in PsA and Ps. (D) Box-plots with relative abundance (parts per unit) of those OTUs marked with * in A–C.
Local gut immune response in PsA is characterized by an increase in fecal sIgA and a decrease in RANKL levels
To further investigate whether this alteration in intestinal microbial communities in PsA or Ps patients was associated with a differential local immune response, we measured luminal concentrations of sIgA in all groups. Given their proposed role in PsA pathogenesis (18), fecal levels of RANKL and OPG were also measured, along with S100 (a novel neutrophil-derived mucosal marker for IBD). PsA patients had an increase in sIgA relative to healthy controls (Figure 3A; P=0.06). Conversely, fecal RANKL was significantly reduced in PsA (Figure 3B; P < 0.05), with only 19% of patients having measurable RANKL compared to 30% of healthy subjects and 75% of Ps patients. Fecal OPG was significantly lower in Ps vs controls (Figure 3C; P < 0.05) but not when compared to PsA. Levels of fecal S100 were similar among all groups. To determine whether fecal quantities of all these proteins were a reflection of systemic levels (which may have translocated into the gut lumen), we performed concomitant serum measurements to calculate correlation coefficients. Although there was no significant correlation between fecal and serum levels of these proteins (Supplementary Figure 4), serum levels of S100 were significantly elevated in Ps patients compared to PsA and healthy subjects (Figure 3D; P < 0.05).
Figure 3. Gut lumen and serum sIgA, RANKL, OPG and S100 levels.
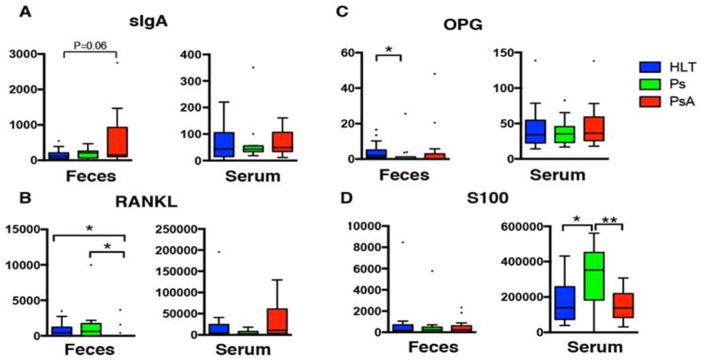
(A) Levels of gut lumen sIgA are higher in PsA patients compared to controls, while measurements in serum revealed no differences among groups. (B) Fecal RANKL were not measurable in most PsA patients, while serum levels were similar among groups. (C) Levels of OPG in fecal samples were higher in Ps vs controls, but similar to those in PsA. (D) Serum S100 levels were significantly higher in Ps compared to PsA and controls. Note: Fecal samples were not available for fecal measurements for two healthy subjects. Fecal RANKL, OPG and S100 were not available in one PsA patient.
MCFAs, but not SCFAs, are decreased in both PsA and Ps fecal samples
Since microbiota-derived SCFAs and MCFAs have been implicated in intestinal health, we determined absolute amounts of these metabolites in all groups. Quantities of fecal SCFAs – including Acetate, Butyrate and Propionate – were similar in all three groups. Interestingly, however, levels of the MCFAs Hexanoate and Heptanoate were significantly reduced in both PsA and Ps compared to controls (Figure 4; P < 0.05, and P < 0.01, respectively).
Figure 4. Fecal levels of short-chain fatty acids (SCFAs) and medium-chain fatty acids (MCFAs).
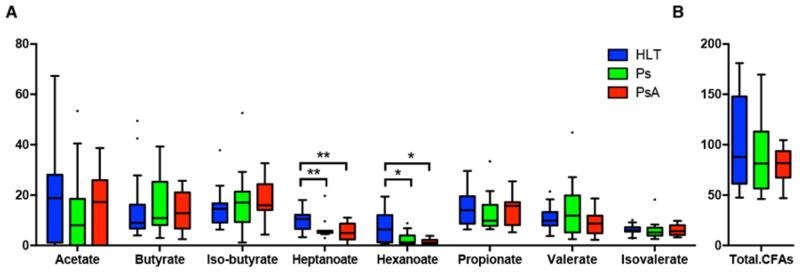
(A) The levels of SCFAs (Acetate, Butyrate, Iso-Butyrate, Propionate, Valerate, Iso-Valerate) and MCFAs (Hexanoate and Heptanoate) were quantified in fecal samples of all groups. While no difference were found in any of the SCFAs, both MCFAs levels were significantly lower in PsA and Ps samples, compared to healthy controls (P<0.01 and P<0.05, respectively). (B) Total levels of fatty acids (CFAs) were similar among groups.
Characteristic interrelations of gut microbiota and metadata in PsA
Because of univariate associations between groups, gut microbiota and metadata, we set out to describe correlations between decreased taxa in Ps and PsA and the various measured fecal and serum proteins and metabolites (Figure 5). An optimal Bayesian network, which incorporates correlations between taxa, was also performed (Supplementary Figure 5). This analysis revealed that, at the genus level, the relative abundance of intestinal Akkermansia and Ruminococcus – both low in PsA – had a significant positive correlation with Heptanoate and Hexanoate (Figure 5A). Akkermansia (as well as OTU85) was inversely correlated with fecal levels of sIgA and the SCFAs Acetate and Butyrate. Interestingly, Coprobacillus, a genus decreased in Ps, negatively correlated with S100 levels in serum. At the OTU level (Figure 5B), OTU43 (Coprococcus), diminished in both Ps and PsA, was also positively correlated with quantities of fecal Heptanoate and Hexanoate. This OTU correlated with OTU31, which in turn positively correlated with OTU109 (Supplementary Figure 5), all of which were also decreased in both groups of patients. OTU16 (UC_Lachnospiraceae), low in PsA, was the only OTU positively correlated with fecal levels of RANKL (also decreased in PsA), and co-occured with two other OTUs that were relatively decreased in PsA: OTU32 and OTU119 (Supplementary Figure 5). Taken together, these interactions describe a potential distinctive pattern representative of the PsA gut microbiota and its metabolites, characterized by lower relative abundance of several taxa and decreased levels of fecal RANKL and MCFAs.
Figure 5. Correlations between gut microbiota and metadata.
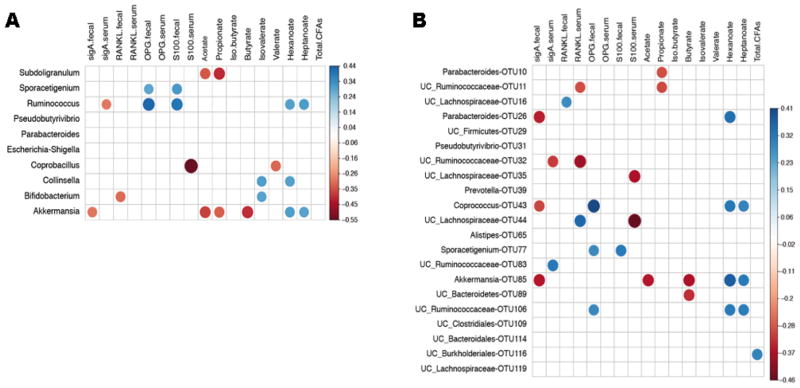
Relative abundance of gut microbiota was correlated with levels of various proteins and fatty acids in serum and/or fecal samples. Heatmaps showing correlations between patient metadata and intestinal microbiota at (A) genus level or (B) OTU level. Circle sizes and color intensity represent magnitude of correlation. Blue circles = positive correlations; red circles = negative correlations.
Finally, the Bayesian network analysis showed that OTUs 10 (Parabacteroides), 44 and 35 (both UC_Lachnospiraceae) also clustered together (Supplementary Figure 5), with the latter taxa revealing the highest inverse correlation with serum levels of S100 (Figure 5B). These interactions differentiate the gut microbiota of the Ps cohort and its associations with systemic inflammatory markers.
DISCUSSION
An expanding body of literature has linked the intestinal microbiota, gut inflammation (both clinical and subclinical), and the different phenotypic expressions of spondyloarthritis.
Two decades ago, seminal work determined the role of gut microbiota in the development of arthritis and colitis in HLA-B27 transgenic rats (7). Since then, many studies have contributed to strengthen this hypothesis, namely that in genetically susceptible subjects a state of gut microbial dysbiosis (alteration in the homeostasis of bacterial composition) promotes an exaggerated immune response in the host’s intestinal lamina propria, activating systemic inflammation and ultimately leading to joint disease. The role of HLA-B27 and related genes was recently validated in AS, IBD and PsA (19). Subclinical histological and molecular markers of gut inflammation are also found in patients with AS (20,21) and with PsA (10). While these studies addressed the activation of host mucosal immunity, they did not directly examine the role of the intestinal microbiome. The link between intestinal bacteria and SpA has been investigated separately. Prior studies, however, utilized indirect serologic methods, classic culture approaches and/or limited, low-throughput PCR/DGGE-based techniques (22), assessing for prevalence of only a handful of taxa.
Utilizing high-throughput, culture-independent, 16S rRNA gene pyrosequencing technology we have shown, for the first time, that patients with PsA and Ps have a decreased diversity in their gut microbiota, mainly due to lower relative abundance of several taxa. Some of this taxa reduction is shared between both conditions [i.e. OTU 43 (Coprococcus)], suggesting a distinctive intestinal gut microbiota that is common in psoriasis of the skin but independent of arthritis. Interestingly, however, other genera such as Ruminoccocus and Akkermansia are uniquely decreased in PsA.
This is intriguing for several reasons. First, similar results have been consistently reported (and replicated) in microbiome studies of patients with IBD, and particularly in those with Crohn’s disease (23). As discussed, SpA is a spectrum of diseases sharing genetic predisposition and phenotypic qualities. It is conceivable therefore that an altered intestinal microbiota constitutes yet another shared feature between IBD, AS and PsA patients. Whether this state of dysbiosis is causative or a consequence of systemic inflammation remains a matter of intense research (and debate).
Second – and perhaps more remarkable – is our finding that this relatively lower diversity in PsA and IBD microbiota is mainly driven by a decrease in phylogenetically similar microbiome members. Several reports showed that Akkermansia and Ruminococcus species, as well as Alistipes genus, are also diminished in IBD patients. Akkermansia is detectable in the majority of healthy subjects (24) and is an important component in two of three recently described human gut enterotypes (25). Using publicly available genetic alignment tools (BLAST), we found that OTU85 has 100% sequence identity to Akkermansia muciniphila and is virtually absent from our PsA cohort. Functionally, A. muciniphila is predominantly a mucus-degrading gut symbiont that converts mucin into short-chain fatty acids (SCFAs) Acetate and Propionate, activating host epithelial cells and stimulating an adequate immune response. Notably, A. muciniphila was the most abundantly identified mucolytic mucosa-associated bacterium in healthy controls vs IBD patients (26–28), suggesting a protective role for this taxon. In two recent studies, Ruminococcaceae were also underrepresented in gut microbiota from IBD patients, particularly in ileal Crohn’s disease (29,30). Many Ruminococcus species, except R. gnavus, are decreased in Crohn’s disease (31). Interestingly, several Ruminococcus species are also mucin-degrading bacteria and important in maintaining gut homeostasis, particularly via the production of SCFAs. SCFAs in turn promote intestinal health, creating favorable conditions for resistance to pathogenic bacteria and protection against colitis (32). None of the SCFAs analyzed in our cohort, however, were significantly lower in PsA patients.
Alistipes, another frequent microorganism of the healthy gut, is significantly reduced in abundance in patients with ileal Crohn’s disease (29), making it the third taxon to be concomitantly underrepresented in both IBD and PsA.
Other gut bacterial genera were relatively decreased in our PsA cohort, including OTU43 (Coprococcus). Although not described as underrepresented in IBD, our network algorithm did reveal that its reduced presence in PsA is directly correlated with significantly lower levels of Hexanoate and Heptanoate (same effect observed for OTU85). Although insufficiently studied, two reports have shown a significant decrease of these MCFAs in IBD patients (33,34). Interestingly, MCFAs display antibacterial effects and were shown to activate peroxisome proliferator activated receptor (PPAR)-γ (35), which in turn is known to ameliorate colitis in animal models (36) and human Crohn’s (37). The relevance of MCFAs-induced PPAR-γ activation in PsA pathogenesis requires further studies.
It is plausible, therefore, that a simultaneous reduction in all these taxa translates into functional consequences on the capacity of predisposed subjects to regulate intestinal immune responses. An inability to contain this altered antigenic load may lead to broader inflammation, either in the gut (as in the case of Crohn’s disease) or in distal compartments, such as entheses or joints (as in PsA and other types of SpA) (3,5). In our studies, increased immune activation in the intestine that accompanied the dysbiosis was reflected by a higher concentration of sIgA in the gut lumen of PsA patients. It is conceivable that this indicates a local mucosal reactive pattern marked by an effort of the host to confine the immune response to the gut. Conversely, this could also represent the initial evidence of a gut barrier breach only seen in individuals with a specific intestinal dysbiosis, as in the case of recent-onset PsA (i.e., decreased diversity; absence of protective taxa). The exact nature of this response, its antigenic target and its cause-effect directionality merit further study.
Intriguingly, the majority of patients with PsA had selectively low levels of gut lumen RANKL compared to Ps subjects and controls. This observation is relevant for several reasons. First, levels of RANKL have been found to be higher in sera of PsA patients (38,39), while RANKL expression was also up-regulated in PsA synovium (40) and in the epidermis of psoriasis patients (41). Second, gut stromal-derived RANKL is the critical factor controlling the differentiation of microfold (M) cells in the intestinal lamina propria, which are crucial for sampling of antigens from the lumen (42). Lastly, in colonic explant cultures from patients with IBD there was an increase in OPG secretion with normal levels of RANKL (43). It is conceivable that this decrease in fecal RANKL levels in PsA patients may actually represent a response to a specific composition of gut bacterial community. Alternatively, the RANKL/OPG system may be a true modulator of gut microbiota and an imbalance in detriment of RANKL in the lumen of PsA patients may facilitate the dissemination of inflammation to distal (joint) sites.
We have previously utilized this approach to examine the intestinal microbiome in treatment-naïve, new-onset RA (NORA) patients (12), and found that expansion of Prevotella copri, was associated with enhanced susceptibility to yet-untreated human RA. This is in contrast with our present findings in PsA patients and suggests a distinctive pattern for both conditions. The significance of this divergence is unclear. Prevotella may represent an environmental trigger for RA in at-risk individuals (e.g., those with circulating autoantibodies) as it has been found to exacerbate induced colitis in mice by our group (12) and others (44). A second possibility is that Prevotella expands preferentially under specific local and systemic inflammatory conditions, representing rather a downstream consequence of RA pathogenesis. These marked differences will require further validation and mechanistic investigations as they may significantly contribute to the understanding of RA and Ps-PsA pathogenesis.
We acknowledge several limitations of our studies. These include the low number of participants and the obvious need for validation cohorts. In the future, it will also be necessary to address differences in gut bacterial communities between patients with axial and peripheral arthritis phenotypes (we could not find such correlations within our cohort). This may be of importance to better understand the potential common pathogenic pathways underlying the axial arthropathies with IBD.
Another key question left unanswered by our study is whether patients with current psoriasis of the skin alone (Ps) will lose certain potentially protective taxa -such as Akkermansia and Ruminoccocus- at the time of (or prior to) transition into PsA. This is crucial because – although it is established that 25–30% of Ps patients will develop arthritis over time – there is currently no possible way to predict progression. A prospective “natural history” cohort of individuals with Ps is ongoing in our center and should help to answer some of these questions.
In summary, our studies constitute a novel and comprehensive approach to investigate the symbiotic relationship between gut microbiota and PsA. We have identified several organisms that are practically absent from PsA patients (i.e., Akkermansia, Ruminococcus), resembling a state of gut dysbiosis previously described in IBD subjects. These taxa and the associated changes in immune response merit further study as potential modulators of autoimmunity in both conditions. Ps gut microbiota profile appears to be intermediate between PsA and healthy subjects, suggesting a possible continuum in disappearing intestinal taxa through the natural history of disease.
Insights into pathogenesis and possible causation, however, will require well-defined human studies using high-throughput sequencing technology, coupled with whole-genome shotgun analyses and in vivo studies in animal models. These investigations may ultimately lead to novel diagnostic tests and interventions, in the form of probiotics, prebiotics, specific microbiome-derived metabolites or molecular targets and even bacterial transplant techniques.
Supplementary Material
Acknowledgments
Supported by: Grant No. RC2 AR058986 to Drs. Abramson and Littman from the National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIAMS) through the American Recovery and Reinvestment Act (ARRA) of 2009; Grant No. K23AR064318 from NIAMS to Dr. Scher; a grant from the American Skin Association to Dr. Marmon; and grants SAF2011-29458 from the Spanish MICINN and the Marie-Curie Career Integration Grant PCIG09-GA-2011-293894 to Dr. Ubeda. The Lucille Castori Center for Microbes, Inflammation, and Cancer, at Memorial Sloan-Kettering Cancer Center, is supported by the Tow Foundation.
The authors wish to thank Drs. Pamela Rosenthal, Peter Izmirly, Jonathan Samuels, Jerome Shupak, Gary Zagon, and Gary Solomon for patient recruitment and performance of clinical assessments; Agnes Viale and Lauren Lipuma (Memorial Sloan-Kettering Cancer Center) and Núria Bonet (University Pompeu Fabra) for sequencing; Drs. Stephen Brown and Chunhai Ruan (Metabolomics Core, BCRF, University of Michigan) for measurement of short- and medium-chain fatty acids; Tim Rostron and Vincenzo Cerundolo (Weatherall Institute of Molecular Medicine, University of Oxford, UK) for HLA sequencing; Ms Rhina Medina for assistance in patient screening, enrollment, scheduling, and data collection; Ms Jyoti Patel for processing, storage, and distribution of samples; and Ms Ann Rupel for assistance in preparation of the manuscript and figures.
Footnotes
AUTHOR CONTRIBUTIONS
All authors were involved in drafting the article or revising it critically for important intellectual content, and all authors approved the final version to be published. Drs. Scher and Ubeda had full access to all of the data in the study and take responsibility for the integrity of the data and the accuracy of the data analysis.
Study conception and design. Scher, Ubeda, Littman, Pamer, Abramson.
Acquisition of data. Scher, Ubeda, Attur, Reddy, Marmon, Neimann, Brusca, Manasson, Patel, Pamer.
Analysis and interpretation of data. Scher, Ubeda, Artacho, Isaac, Attur, Brusca, Manasson, Patel, Littman, Pamer, Abramson.
References
- 1.Eder L, Chandran V, Pellet F, Shanmugarajah S, Rosen CF, Bull SB, et al. Human leucocyte antigen risk alleles for psoriatic arthritis among patients with psoriasis. Ann Rheum Dis. 2012;71:50–5. doi: 10.1136/ard.2011.155044. [DOI] [PubMed] [Google Scholar]
- 2.Winchester R, Minevich G, Steshenko V, Kirby B, Kane D, Greenberg DA, et al. HLA associations reveal genetic heterogeneity in psoriatic arthritis and in the psoriasis phenotype. Arthritis Rheum. 2012;64:1134–44. doi: 10.1002/art.33415. [DOI] [PubMed] [Google Scholar]
- 3.Cua DJ, Sherlock JP. Autoimmunity’s collateral damage: Gut microbiota strikes ‘back’. Nat Med. 2011;17:1055–6. doi: 10.1038/nm0911-1055. [DOI] [PubMed] [Google Scholar]
- 4.Van Praet L, Van den Bosch F, Mielants H, Elewaut D. Mucosal inflammation in spondylarthritides: past, present, and future. Curr Rheumatol Rep. 2011;13:409–15. doi: 10.1007/s11926-011-0198-2. [DOI] [PubMed] [Google Scholar]
- 5.Sherlock JP, Joyce-Shaikh B, Turner SP, Chao CC, Sathe M, Grein J, et al. IL-23 induces spondyloarthropathy by acting on ROR-gammat+ CD3+CD4-CD8- entheseal resident T cells. Nat Med. 2012;18:1069–76. doi: 10.1038/nm.2817. [DOI] [PubMed] [Google Scholar]
- 6.Van Praet L, Van den Bosch FE, Jacques P, Carron P, Jans L, Colman R, et al. Microscopic gut inflammation in axial spondyloarthritis: a multiparametric predictive model. Ann Rheum Dis. 2013;72:414–7. doi: 10.1136/annrheumdis-2012-202135. [DOI] [PubMed] [Google Scholar]
- 7.Rath HC, Herfarth HH, Ikeda JS, Grenther WB, Hamm TE, Jr, Balish E, et al. Normal luminal bacteria, especially Bacteroides species, mediate chronic colitis, gastritis, and arthritis in HLA-B27/human beta2 microglobulin transgenic rats. J Clin Invest. 1996;98:945–53. doi: 10.1172/JCI118878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ruutu M, Thomas G, Steck R, gli-Esposti MA, Zinkernagel MS, Alexander K, et al. beta-glucan triggers spondylarthritis and Crohn’s disease-like ileitis in SKG mice. Arthritis Rheum. 2012;64:2211–22. doi: 10.1002/art.34423. [DOI] [PubMed] [Google Scholar]
- 9.Orchard TR, Wordsworth BP, Jewell DP. Peripheral arthropathies in inflammatory bowel disease: their articular distribution and natural history. Gut. 1998;42:387–91. doi: 10.1136/gut.42.3.387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Scarpa R, Manguso F, D’Arienzo A, D’Armiento FP, Astarita C, Mazzacca G, et al. Microscopic inflammatory changes in colon of patients with both active psoriasis and psoriatic arthritis without bowel symptoms. J Rheumatol. 2000;27:1241–6. [PubMed] [Google Scholar]
- 11.Li WQ, Han JL, Chan AT, Qureshi AA. Psoriasis, psoriatic arthritis and increased risk of incident Crohn’s disease in US women. Ann Rheum Dis. 2013;72:1200–5. doi: 10.1136/annrheumdis-2012-202143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Scher JU, Sczesnak A, Longman RS, Segata N, Ubeda C, Bielski C, et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. Elife. 2013;2:e01202. doi: 10.7554/eLife.01202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Scher JU, Ubeda C, Equinda M, Khanin R, Buischi Y, Viale A, et al. Periodontal disease and the oral microbiota in new-onset rheumatoid arthritis. Arthritis Rheum. 2012;64:3083–94. doi: 10.1002/art.34539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sheneman L, Evans J, Foster JA. Clearcut: a fast implementation of relaxed neighbor joining. Bioinformatics. 2006;22:2823–4. doi: 10.1093/bioinformatics/btl478. [DOI] [PubMed] [Google Scholar]
- 15.Nielsen OH, Gionchetti P, Ainsworth M, Vainer B, Campieri M, Borregaard N, et al. Rectal dialysate and fecal concentrations of neutrophil gelatinase-associated lipocalin, interleukin-8, and tumor necrosis factor-alpha in ulcerative colitis. Am J Gastroenterol. 1999;94:2923–8. doi: 10.1111/j.1572-0241.1999.01439.x. [DOI] [PubMed] [Google Scholar]
- 16.Segata N, Izard J, Waldron L, Gevers D, Miropolsky L, Garrett WS, et al. Metagenomic biomarker discovery and explanation. Genome Biol. 2011;12:R60. doi: 10.1186/gb-2011-12-6-r60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Scurati M. Learning Bayesian Networks with the bnlearn R Package. Journal of Statistical Software. 2010;35(3):1–22. [Google Scholar]
- 18.Mensah KA, Ritchlin CT, Schwarz EM. RANKL induces heterogeneous DC-STAMP(lo) and DC-STAMP(hi) osteoclast precursors of which the DC-STAMP(lo) precursors are the master fusogens. J Cell Physiol. 2010;223:76–83. doi: 10.1002/jcp.22012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Reveille JD. Genetics of spondyloarthritis--beyond the MHC. Nat Rev Rheumatol. 2012;8:296–304. doi: 10.1038/nrrheum.2012.41. [DOI] [PubMed] [Google Scholar]
- 20.Mielants H, Veys EM, Cuvelier C, De Vos M, Goemaere S, De Clercq L, et al. The evolution of spondyloarthropathies in relation to gut histology. II. Histological aspects. J Rheumatol. 1995;22:2273–8. [PubMed] [Google Scholar]
- 21.Ciccia F, Accardo-Palumbo A, Alessandro R, Rizzo A, Principe S, Peralta S, et al. Interleukin-22 and interleukin-22-producing NKp44+ natural killer cells in subclinical gut inflammation in ankylosing spondylitis. Arthritis Rheum. 2012;64:1869–78. doi: 10.1002/art.34355. [DOI] [PubMed] [Google Scholar]
- 22.Stebbings S, Munro K, Simon MA, Tannock G, Highton J, Harmsen H, et al. Comparison of the faecal microflora of patients with ankylosing spondylitis and controls using molecular methods of analysis. Rheumatology (Oxford) 2002;41:1395–401. doi: 10.1093/rheumatology/41.12.1395. [DOI] [PubMed] [Google Scholar]
- 23.Manichanh C, Rigottier-Gois L, Bonnaud E, Gloux K, Pelletier E, Frangeul L, et al. Reduced diversity of faecal microbiota in Crohn’s disease revealed by a metagenomic approach. Gut. 2006;55:205–11. doi: 10.1136/gut.2005.073817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Collado MC, Derrien M, Isolauri E, de Vos WM, Salminen S. Intestinal integrity and Akkermansia muciniphila, a mucin-degrading member of the intestinal microbiota present in infants, adults, and the elderly. Appl Environ Microbiol. 2007;73:7767–70. doi: 10.1128/AEM.01477-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human gut microbiome. Nature. 2011;473(7346):174–80. doi: 10.1038/nature09944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Png CW, Linden SK, Gilshenan KS, Zoetendal EG, McSweeney CS, Sly LI, et al. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am J Gastroenterol. 2010;105:2420–8. doi: 10.1038/ajg.2010.281. [DOI] [PubMed] [Google Scholar]
- 27.Vigsnaes LK, Brynskov J, Steenholdt C, Wilcks A, Licht TR. Gram-negative bacteria account for main differences between faecal microbiota from patients with ulcerative colitis and healthy controls. Benef Microbes. 2012;3:287–97. doi: 10.3920/BM2012.0018. [DOI] [PubMed] [Google Scholar]
- 28.Rajilic-Stojanovic M, Shanahan F, Guarner F, de Vos WM. Phylogenetic analysis of dysbiosis in ulcerative colitis during remission. Inflamm Bowel Dis. 2013;19:481–8. doi: 10.1097/MIB.0b013e31827fec6d. [DOI] [PubMed] [Google Scholar]
- 29.Willing BP, Dicksved J, Halfvarson J, Andersson AF, Lucio M, Zheng Z, et al. A pyrosequencing study in twins shows that gastrointestinal microbial profiles vary with inflammatory bowel disease phenotypes. Gastroenterology. 2010;139:1844–54. doi: 10.1053/j.gastro.2010.08.049. [DOI] [PubMed] [Google Scholar]
- 30.Morgan XC, Tickle TL, Sokol H, Gevers D, Devaney KL, Ward DV, et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012;13:R79. doi: 10.1186/gb-2012-13-9-r79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Joossens M, Huys G, Cnockaert M, De Preter V, Verbeke K, Rutgeerts P, et al. Dysbiosis of the faecal microbiota in patients with Crohn’s disease and their unaffected relatives. Gut. 2011;60:631–7. doi: 10.1136/gut.2010.223263. [DOI] [PubMed] [Google Scholar]
- 32.Smith PM, Howitt MR, Panikov N, Michaud M, Gallini CA, Bohlooly Y, et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science. 2013;341(6145):569–73. doi: 10.1126/science.1241165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Garner CE, Smith S, de Lacy CB, White P, Spencer R, Probert CS, et al. Volatile organic compounds from feces and their potential for diagnosis of gastrointestinal disease. FASEB J. 2007;21:1675–88. doi: 10.1096/fj.06-6927com. [DOI] [PubMed] [Google Scholar]
- 34.De Preter V, Machiels K, Joossens M, Arijs I, Matthys C, Vermeire S, et al. Faecal metabolite profiling identifies medium-chain fatty acids as discriminating compounds in IBD. Gut. 2014 doi: 10.1136/gutjnl-2013-306423. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 35.Liberato MV, Nascimento AS, Ayers SD, Lin JZ, Cvoro A, Silveira RL, et al. Medium chain fatty acids are selective peroxisome proliferator activated receptor (PPAR) gamma activators and pan-PPAR partial agonists. PLoS One. 2012;7(5):e36297. doi: 10.1371/journal.pone.0036297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bassaganya-Riera J, Viladomiu M, Pedragosa M, De SC, Carbo A, Shaykhutdinov R, et al. Probiotic bacteria produce conjugated linoleic acid locally in the gut that targets macrophage PPAR gamma to suppress colitis. PLoS One. 2012;7(2):e31238. doi: 10.1371/journal.pone.0031238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bassaganya-Riera J, Hontecillas R, Horne WT, Sandridge M, Herfarth HH, Bloomfeld R, et al. Conjugated linoleic acid modulates immune responses in patients with mild to moderately active Crohn’s disease. Clin Nutr. 2012;31(5):721–7. doi: 10.1016/j.clnu.2012.03.002. [DOI] [PubMed] [Google Scholar]
- 38.Chandran V, Cook RJ, Edwin J, Shen H, Pellett FJ, Shanmugarajah S, et al. Soluble biomarkers differentiate patients with psoriatic arthritis from those with psoriasis without arthritis. Rheumatology (Oxford) 2010;49:1399–405. doi: 10.1093/rheumatology/keq105. [DOI] [PubMed] [Google Scholar]
- 39.Xue Y, Jiang L, Cheng Q, Chen H, Yu Y, Lin Y, et al. Adipokines in psoriatic arthritis patients: the correlations with osteoclast precursors and bone erosions. PLoS One. 2012;7(10):e46740. doi: 10.1371/journal.pone.0046740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ritchlin CT, Haas-Smith SA, Li P, Hicks DG, Schwarz EM. Mechanisms of TNF-alpha- and RANKL-mediated osteoclastogenesis and bone resorption in psoriatic arthritis. J Clin Invest. 2003;111:821–31. doi: 10.1172/JCI16069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Toberer F, Sykora J, Gottel D, Ruland V, Hartschuh W, Enk A, et al. Tissue microarray analysis of RANKL in cutaneous lupus erythematosus and psoriasis. Exp Dermatol. 2011;20:600–2. doi: 10.1111/j.1600-0625.2011.01303.x. [DOI] [PubMed] [Google Scholar]
- 42.Knoop KA, Kumar N, Butler BR, Sakthivel SK, Taylor RT, Nochi T, et al. RANKL is necessary and sufficient to initiate development of antigen-sampling M cells in the intestinal epithelium. J Immunol. 2009;183:5738–47. doi: 10.4049/jimmunol.0901563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Moschen AR, Kaser A, Enrich B, Ludwiczek O, Gabriel M, Obrist P, et al. The RANKL/OPG system is activated in inflammatory bowel disease and relates to the state of bone loss. Gut. 2005;54:479–87. doi: 10.1136/gut.2004.044370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Elinav E, Strowig T, Kau AL, Henao-Mejia J, Thaiss CA, Booth CJ, et al. NLRP6 inflammasome regulates colonic microbial ecology and risk for colitis. Cell. 2011;145:745–57. doi: 10.1016/j.cell.2011.04.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.