
Figure 1. Comparison of actin nucleation and elongation mechanisms in the dendritic spine and the lamellipodium.
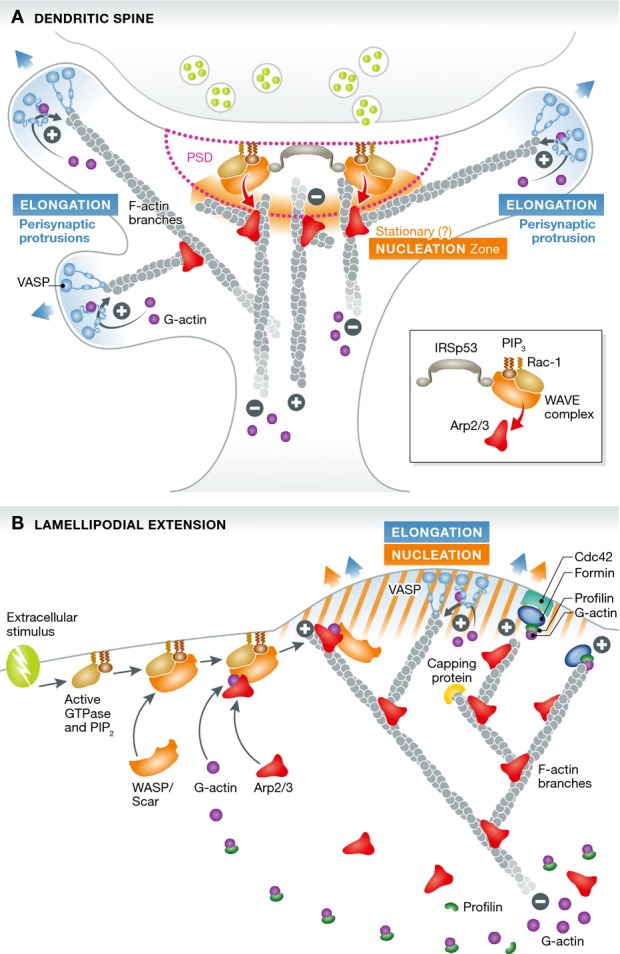
(A) In spines, the post-synaptic density functions as a hub for actin nucleation, concentrating signaling and regulatory factors (IRSp53, Rac1, WAVE complex) in order to activate the Arp2/3 complex in response to extra-synaptic signaling, recruiting it from a freely diffusible pool. Actin polymerization then proceeds in a slow and unpolarized fashion. Spine motility is ensured by protrusions stemming possibly by branched filaments on the sides of the PSD (peri-synaptic) in which formin-dependent elongation mechanisms at their tips and on the membrane generate a fast and polarized rear-flow of actin and exert the pushing force (from Chazeau et al). (B) In lamellipodia, both actin nucleation and rapid VASP- and formin-dependent elongation occur in synchrony at the membrane, where the pushing force is needed to extrude the lamellipodium, generating a fast and polarized rear-flow of actin and a complex branched actin network.