

Abstract
Exposures to lead (Pb) are associated with neurological problems including psychiatric disorders and impaired learning and memory. Pb can be absorbed by iron transporters, which are up-regulated in hereditary hemochromatosis, an iron overload disorder in which increased iron deposition in various parenchymal organs promote metal-induced oxidative damage. While dysfunction in HFE (High Fe) gene is the major cause of hemochromatosis, the transport and toxicity of Pb in Hfe-related hemochromatosis are largely unknown. To elucidate the relationship between HFE gene dysfunction and Pb absorption, H67D knock-in Hfe-mutant and wild-type mice were given drinking water containing Pb 1.6 mg/ml ad libitum for 6 weeks and examined for behavioral phenotypes using the nestlet-shredding and marble-burying tests. Latency to nestlet-shredding in Pb-treated wild-type mice was prolonged compared with non-exposed wild-types (p < 0.001), whereas Pb exposure did not alter shredding latency in Hfe-mutant mice. In the marble-burying test, Hfe-mutant mice showed an increased number of marbles buried compared with wild-type mice (p = 0.002), indicating more repetitive behavior upon Hfe mutation. Importantly, Pb-exposed wild-type mice buried more marbles than non-exposed wild-types, whereas the number of marbles buried by Hfe-mutant mice did not change whether or not exposed to Pb. These results suggest that Hfe mutation could normalize Pb-induced behavioral alteration. To explore the mechanism of repetitive behavior caused by Pb, western blot analysis was conducted for proteins involved in brain dopamine metabolism. The levels of tyrosine hydroxylase and dopamine transporter increased upon Pb exposure in both genotypes, whereas Hfe-mutant mice displayed down-regulation of the dopamine transporter and dopamine D1 receptor with D2 receptor elevated. Taken together, our data support the idea that both Pb exposure and Hfe mutation increase repetitive behavior in mice and further suggest that these behavioral changes could be associated with altered dopaminergic neurotransmission, providing a therapeutic basis for psychiatric disorders caused by Pb toxicity.
Keywords: HFE, Hemochromatosis, Marble-burying, Nestlet-shredding, Selenium, Autism
INTRODUCTION
Lead (Pb) is a heavy metal that is non-essential for body function but is toxic at high levels. In humans, excess Pb causes adverse effects, such as hyperactivity, cognitive deficits and other behavioral alterations (1). Even very low levels of Pb (below 5 μg/dl blood) can induce neurotoxic effects in children. Moreover, high blood levels of Pb in young children are associated with increased neurobehavioral deficits and the development of attention deficit hyperactivity disorder (ADHD) (2). In animals, hyperactivity and impaired learning are observed in the rat pups exposed to Pb during gestation (3).
Exposures to Pb occur in many different areas of people’s daily lives. Various food products, dietary supplements, and drinking water all serve as sources of Pb exposure (1,4). Significant sources of Pb are also found in the environment. Although paint containing Pb has been banned in the US after its toxicity has been observed, 38 million living units remain painted with Pb-loaded paint, and 65% of them are considered significant risk areas (1,4). Some mining sites are significant risk areas for Pb toxicity as well. For example, children around the Tar Creek in Ottawa County, Oklahoma, USA, a former major site for Pb and zinc mining, may exhibit signs of metal toxicity, including impairment of cognitive development (5).
Pb is well absorbed through ingestion, inhalation and skin penetration. An increasing amount of evidence has indicated that Pb transport is mediated by divalent metal transporter 1 (DMT1), a major transporter for iron and several other divalent metals (6). DMT1’s primary function is to absorb dietary iron for various essential functions, including hemoglobin synthesis and myelination of the nervous system. Pb absorption is enhanced in iron-deficient anemia (7), likely due to compensatory up-regulation of metal transporters, which suggests that altered iron status in the body can influence the transport and toxicity of Pb. This possibility, however, has not been tested in the iron overload disorder hemochromatosis. Hemochromatosis is characterized by elevated intestinal absorption and progressive tissue deposition of iron that leads to cardiomyopathy, liver cirrhosis, and premature death (8). Increased iron is believed to promote the formation of the free radical species, which results in organ damage. Polymorphism in the HFE (High iron or Fe) gene is the leading cause of adult onset hereditary hemochromatosis, which is one of the most common genetic diseases in the North American Caucasian population (8). C282Y and H63D are the two most prevalent HFE missense variants, accounting for 5~32% in the North American population (9) and up to 8% in the Korean population (10). Mutations in HFE have also been associated with neurodegenerative diseases including Alzheimer’s and Parkinson’s diseases (11). Notably, both HFE polymorphism and Pb exposure have been implicated in a spectrum of neurodevelopmental disorders (1,4,12). However, despite a huge prevalence in HFE missense variants (9), whether or not mutations in HFE gene alter Pb-induced neurotoxicity has yet to be examined. Therefore, in the current study we investigated the role of HFE mutation on repetitive behavior and dopamine metabolism by using Hfe-mutant and wild-type mice that were exposed to Pbcontaining water.
MATERIALS AND METHODS
Animals and Pb exposure. Mice that carry a knock-in mutation in H67D Hfe gene, which is homologous to human H63D polymorphism (13,14), and their control wild-type mice were kindly provided by Dr. James Connor (Pennsylvania State University College of Medicine, Hershey, PA, USA). We chose this model for two reasons: first, polymorphism in H63D (10~32%) is more frequently found than that in C282Y (5~17%) (9). Second, H63D has been shown to be linked to neurodegenerative diseases (13,14) and neurodevelopmental disorders (12). Only female mice were used because it has been recognized that females and children are more vulnerable to metal toxicity (15). Weanling female mice were fed facility chow (Prolab Isopro RMH 3000, LabDiet; 380 mg iron per kg diet) and exposed ad libitum to facility water or facility water containing Pb (1.6 mg as Pb; 2.5 mg as Pb acetate trihydrate per ml water; Sigma-Aldrich, St. Louis, MO, USA). This concentration was chosen because previous studies have demonstrated that animals exposed to 2.8 mg/ml Pb exhibited several physiological alterations (16). Pb was not detected in facility water. At the age of 7 wks (i.e., 4 wks since the start of Pb exposure), mice were examined for behavior tests described below in the span of 2 wks. All experimental procedures were previously approved by Northeastern University Institutional Animal Care and Use Committee.
Nestlet-shredding test. In order to examine repetitive behavior, each mouse was acclimated to a test cage similar to their home cage without food or water. After 15 min, a cotton nestlet (5 cm × 5 cm; Ancare, Bellmore, NY, USA) was given to each mouse in the cage for two hr. The nestlet was weighed before and after the test to calculate the percentage of nestlet shredded. The test was conducted for two consecutive days and the average of the two tests was used for analysis.
Marble-burying test. The marble-burying test was conducted as another method of detecting innate repetitive behavior of the rodents. Briefly, the animals were acclimated for 15~20 mins in the cages with 4~5 cm of bedding (the same material as their home cage bedding). After the acclimation period, the animals were taken out and the marbles were laid out in a 4 × 5 arrangement. The animals were then returned to the cage and left for 15 min, after which the marbles (both buried and remaining) were counted; marbles were considered as buried if 50% or more was buried under the bedding. The test was conducted for three consecutive days and the average of the three tests was used for analysis.
Tissue collection. After the last behavioral test, animals were euthanized by overdose of isoflurane. Brain and liver tissues were harvested, snap-frozen in liquid nitrogen and stored at −80℃ until analysis.
Metal analysis. Liver and brain samples were digested in 1 ml of 20% nitric acid (Trace grade, Fisher Scientific; Pittsburgh, PA, USA) at 125℃ for 1 hr. After a complete digestion, the resulting samples were diluted with metalfree double-distilled water up to a volume of 10 ml. Metal concentrations were determined by inductively coupled plasma mass spectrometry (ICP-MS) (Varian 810/820MS, Bruker, Billerica, MA, USA).
Non-heme iron analysis. Non-heme iron serves as a surrogate marker for body iron status and storage. Liver samples were incubated in a 15-fold volume of acid solution (10% trichloroacetic acid, 3 M HCl) in 65℃ water bath for 20 hr. Samples (0.08 ml) were first mixed with reaction buffer (10% thioglycolic acid in 1 M sodium acetate) followed by a mixture with 1% bathophenanthroline disulfonic acid for colorimetric reaction. The optical density was measured using UV/Vis spectrophotometer at 535 nm. Nonheme iron concentration was determined based on seriallydiluted iron standard solutions. Data were presented in ppm (i.e., μg metal per gram of wet tissue weight).
Western blot analysis. Snap-frozen brain tissues were homogenized in RIPA buffer (50 mM Tris, 0.1% SDS, 1% NP40, 0.5% sodium deoxycholate, pH 7.5) containing protease inhibitors (Complete Mini, Roche; Nutley, NJ, USA). Tissue homogenates were centrifuged at 16,000 g for 6 min at 4℃. Protein concentrations in brain homogenates were determined by the Bradford assay. The tissue extracts (40 μg protein) were electrophoresed on 10% gels (for all proteins except ferritin) or 12.5% (for ferritin) and transferred to nitrocellulose membranes (GE Healthcare, Pittsburgh, PA, USA). The membranes were incubated with blocking solution (5% non-fat milk in TBS) for 1 hr at room temperature, followed by overnight incubation with primary antibodies in 2% non-fat milk at 4℃. The antibodies used were rabbit antibody against dopamine D1 receptor (D1DR, 1:500, Abcam; Cambridge, MA, USA), mouse antibody against dopamine D2 receptor (D2DR, 1 : 100, Santa Cruz Biotech, Dallas, Texas), goat antibody against dopamine transporter (DAT, 1 : 200, Santa Cruz Biotech), mouse antibody against tyrosine hydroxylase (TH, 1 : 200, Santa Cruz Biotech) or rabbit antibody against ferritin (Ftn, 1 : 500, Santa Cruz Biotech). Blots were probed with mouse anti-actin (1 : 5,000, MP Biomedicals; Santa Ana, CA, USA) as a loading control. Secondary antibodies were peroxidaselabeled goat anti-rabbit antibodies (1 : 1,000, Santa Cruz Biotech), sheep anti-mouse antibodies (1 : 1,000, GE Healthcare), or donkey anti-goat antibodies (1 : 1,000, Santa Cruz Biotech). Immunoreactivity was detected using ECL West Dura substrate (Thermo Scientific; Waltham, MA, USA). Protein bands were visualized by ChemiDoc XRS (Bio- Rad, Hercules, CA, USA) and intensities of protein bands were quantified using Image Lab software (version 4.1, Bio-Rad).
Statistical analysis. Values reported were expressed as mean ± SEM. Statistical analyses were performed using SigmaPlot (version 12.3; Systat Software Inc., San Jose, CA, USA) software to determine the main individual effects as well as the result of the interaction between Hfe mutation and Pb exposure via drinking water. Posthoc comparisons were performed by the Holm-Sidak method. A significant difference was considered at p < 0.05.
RESULTS
Physiological and hematological characteristics of lead exposure in Hfe mutation. There was a significant effect of gene on body weight; Hfe-mutant mice were heavier than wild-type control mice (p = 0.005; Fig. 1A). However, Pb treatment did not alter body weight. Similarly, hematocrit values were higher in mutants than in wild-type mice (p < 0.001), but were unaffected by Pb treatment (Fig. 1B). The levels of hepatic non-heme iron, a surrogate for iron storage in the liver, were also elevated as a result of the difference in genotype (p < 0.001; Fig. 1C). This result was consistent with the increased levels of ferritin (Ftn), the major iron storage protein, in the liver upon Hfe mutation (Fig. 1D). However, Pb exposure did not change non-heme iron in the liver. These results indicate that Hfe mutation increases body iron status, which is not modified by Pb exposure.
Fig. 1. Physiological parameters of mice exposed to Pb 1.6mg/ml by drinking water. Whole blood and liver were collected from mice exposed to 1.6mg/ml of Pb or facility water. The body weights (A) and hematocrit values (B) were determined at the time of euthanasia (n = 6~10 per group). The concentrations of liver non-heme iron (C) were determined by spectrophotometric analysis using bathophenathroline disulfonate (n = 5~9 per group). The levels of ferritin (Ftn) in the liver (D) were determined by Western blot analysis (n = 4~8 per group). Open and closed bars represent water and Pb treatment, respectively. All data were presented as mean ± SEM. All the P values were calculated by two-way ANOVA, followed by post-hoc comparisons. WT, wild-type; MUT, Hfe-mutant.

Metal concentrations in liver and brain of Pb-exposed Hfe-mutant mice. Liver tissues collected from the mice exposed to Pb by drinking water were analyzed to determine the levels of associated metals by inductively-coupled plasma mass spectrometry. There was a significant increase in iron levels (both heme and non-heme) in the liver from Hfe-mutants compared with wild-type mice (p < 0.001; Fig. 2A). Similar to non-heme iron, Pb exposure did not alter the status of total iron in the liver. While liver Pb in water-treated groups was undetected, mice exposed to Pb increased Pb stores in the liver, which was altered by Hfe mutation (Fig. 2B). We examined selenium status in the liver since Pb can interfere with metabolism of selenium, an element Fig. 4. Effect of Pb exposure via drinking water on nestletshredding behavior of Hfe-mutant mice. Nestlet-shredding behavior (A) over a period of 2 hr was measured by the percentage of nestlet shredded (n = 6~10 per group) and latency to the beginning of shredding (B). Open and closed bars represent water and Pb treatment, respectively. All data were presented as mean ± SEM. All the P values were calculated by twoway ANOVA, followed by post-hoc comparisons. that supports anti-oxidant activity by multiple selenocysteine- requiring anti-oxidant enzymes and transporters; however, we found no change in selenium levels among the four groups (Fig. 2C).
Fig. 2. Metal status in the liver of Pb-exposed mice measured by ICP-MS. Liver tissues collected from the mice exposed to Pb at 1.6mg/ml by drinking water were used to determine the levels of Fe (A), Pb (B), and Se (C) levels by inductively-coupled plasma mass spectrometry (n = 6 per group). Open and closed bars represent water and Pb treatment, respectively. All data were presented as mean ± SEM. The P values for Fe and Se levels were calculated by two-way ANOVA, followed by post-hoc comparisons, whereas those for Pb levels were calculated by the Student’s t-test because Pb was not detected in watertreated groups. ND, not detected.
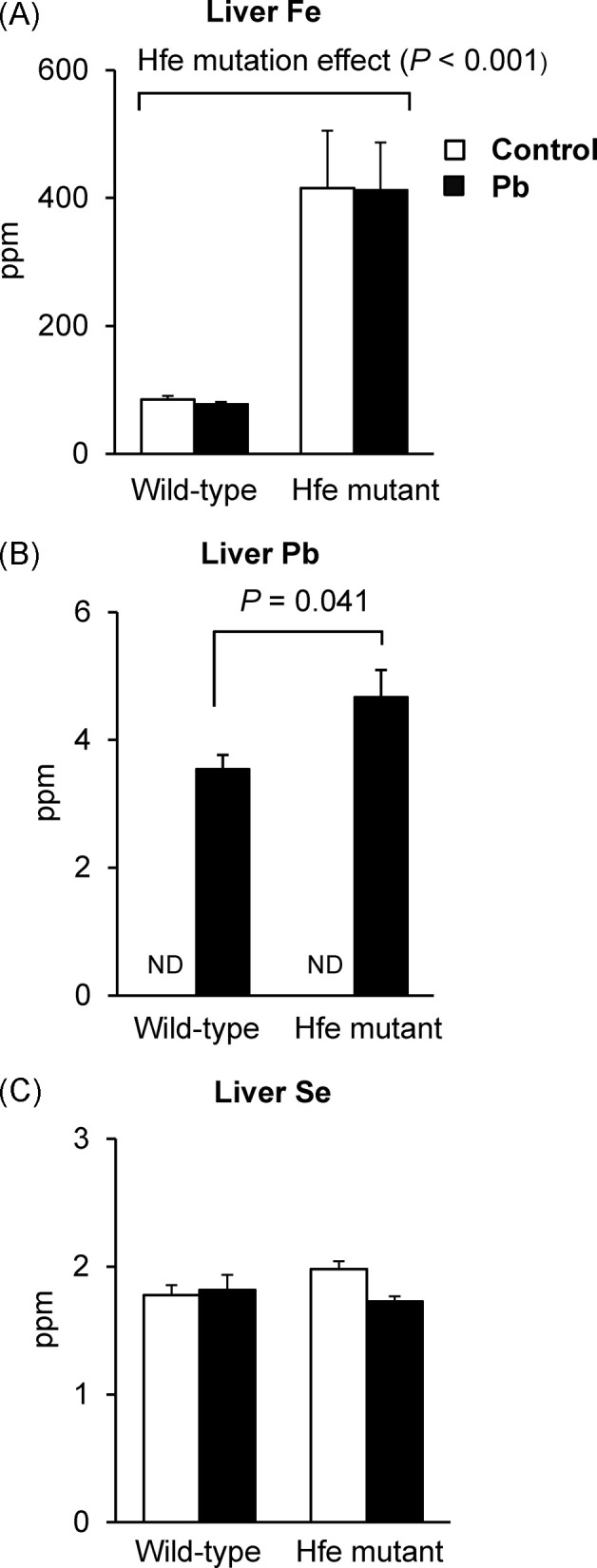
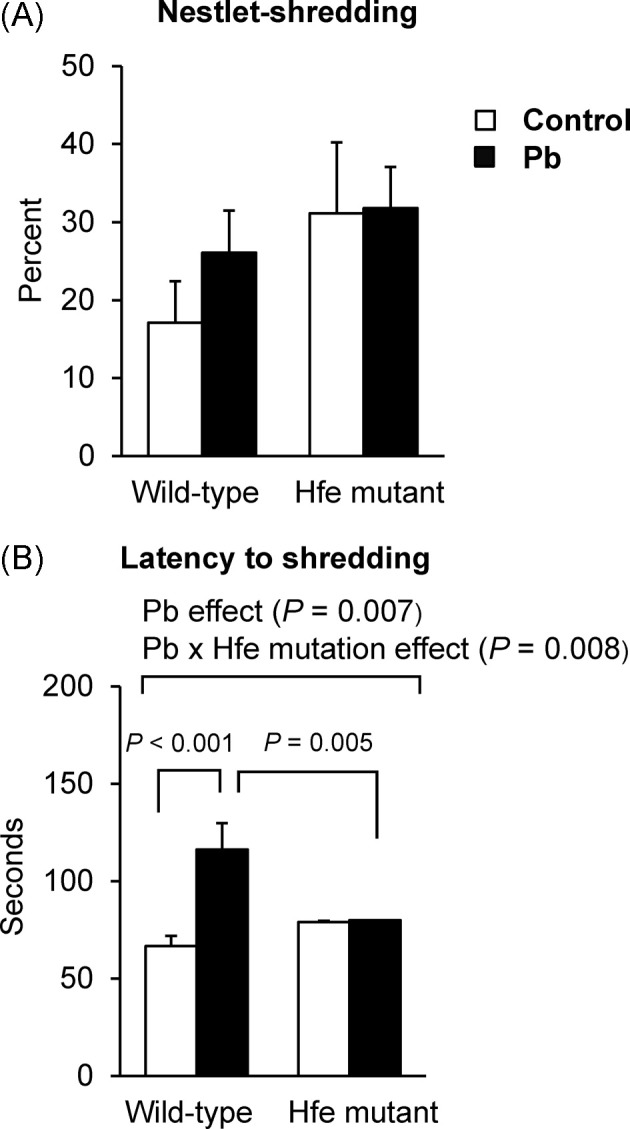
Fig. 4. Effect of Pb exposure via drinking water on nestletshredding behavior of Hfe-mutant mice. Nestlet-shredding behavior (A) over a period of 2 hr was measured by the percentage of nestlet shredded (n = 6~10 per group) and latency to the beginning of shredding (B). Open and closed bars represent water and Pb treatment, respectively. All data were presented as mean ± SEM. All the P values were calculated by twoway ANOVA, followed by post-hoc comparisons.
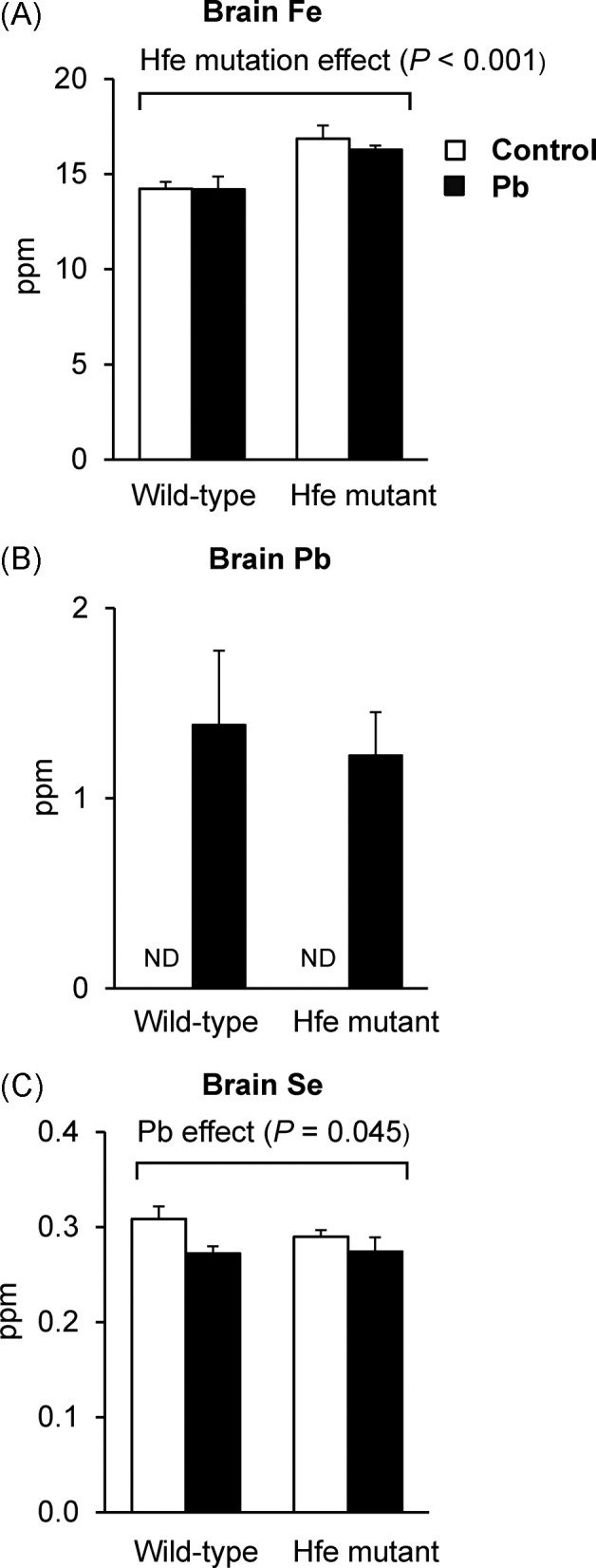
Similarly, brain iron levels were elevated upon Hfe mutation (Fig. 3A), while no difference was observed after Pb treatment. Pb levels were dramatically increased in the Pb-exposed group, regardless of Hfe mutation (Fig. 3B). Interestingly, selenium levels were decreased in Pb-exposed mice (p = 0.045), while Hfe mutation did not change selenium status (Fig. 3C). Combined, these results indicate that increased Pb levels by drinking water could deplete Se-mediated anti-oxidant capacity in the brain, but not in the liver, while Hfe mutation does not alter Pb status in the brain.
Fig. 3. Metal status in the brain of Pb-exposed mice measured by ICP-MS. Whole brain tissues collected from the mice exposed to Pb at 1.6mg/ml by drinking water were used to determine the levels of Fe (A), Pb (B), and Se (C) by inductively-coupled plasma mass spectrometry (n = 4~5 per group). Open and closed bars represent water and Pb treatment, respectively. All data were presented as mean ± SEM. All the P values were calculated by two-way ANOVA, followed by post-hoc comparisons, whereas those for Pb levels were calculated by the Student’s t-test because Pb was not detected in water-treated groups. ND, not detected.
Lead exposure increases latency of nestlet-shredding in wild-type but not in Hfe-mutant mice. The nestletshredding test was conducted in order to determine whether Pb exposure would increase repetitive behavior and whether Hfe mutation could alter this response. The amount of nestlet shredded did not differ between mutants and wild-types although there was a higher trend in mutants (p = 0.083; Fig. 4A). Interestingly, the latency to the beginning of nestlet-shredding increased upon Pb exposure (p = 0.007, Pb effect; Fig. 4B). Moreover, we observed an interaction effect between Hfe mutation and Pb exposure (p = 0.008); the latency to nestlet-shredding in Pb-treated wild-types was prolonged compared with non-exposed wild-types (p < 0.001), whereas Pb exposure did not alter the shredding latency in mutant mice.
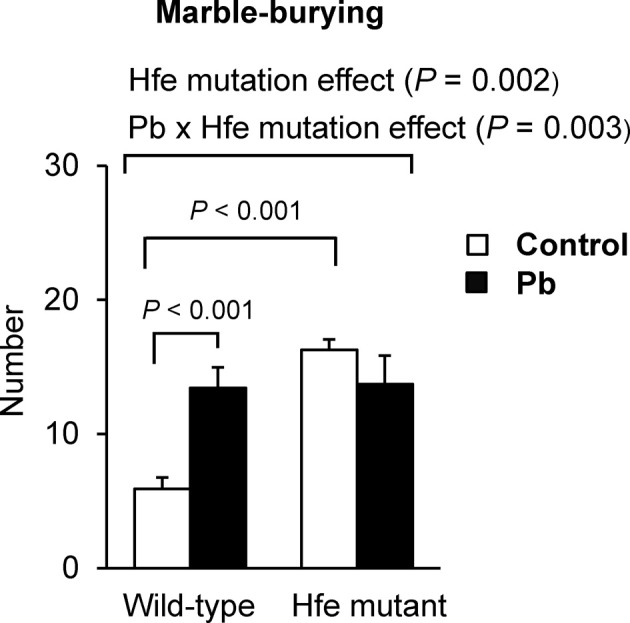
Lead exposure increases marble-burying behavior in wild-type but not in Hfe-mutant mice. The marbleburying test was conducted as another measurement of repetitive behavior. The mutant mice in general showed an increased number of marbles buried compared with wildtype mice (p = 0.002; Fig. 5), indicating that Hfe mutation is associated with elevated repetitive behavior. In addition, wild-type mice exposed to Pb buried more marbles than the mice exposed to water (p < 0.001), whereas Hfe-mutant mice did not change the number of marbles buried whether or not exposed to Pb. It is possible that Hfe-mutant mice had already achieved the maximum performance of marbleburying such that no significant effect of Pb exposure was observed in these mice. Nonetheless, these results still indicate that marble-burying behavior differs between Hfemutants and wild-type controls upon Pb exposure (interaction effect; p = 0.003).
Fig. 5. Effect of Pb exposure via drinking water on marbleburying behavior of Hfe-mutant mice. A tendency to perform repetitive behavior was measured by the marble-burying test. The numbers of marbles buried over a period of 15 min in the span of three days were averaged (n = 6~10 per group). Open and closed bars represent water and Pb treatment, respectively. All data were presented as mean ± SEM. All the P values were calculated by two-way ANOVA, followed by post-hoc comparisons.
Both Pb exposure and Hfe mutation influence dopaminergic neurotransmission. To evaluate the role of Pb exposure and Hfe mutation in dopamine signaling pathway, which has been shown to be involved in emotional behavior, we determined the levels of dopamine-related proteins. First, TH, a critical enzyme for dopamine production, was elevated in Pb-exposed mice (p = 0.003), which was not altered by Hfe mutation (Fig. 6A). Second, Hfe-mutant mice showed decreased levels of DAT, the major transporter responsible for the reuptake of synaptic dopamine, compared with wild-type mice (p = 0.017; Fig. 6B), whereas Pb exposure increased DAT expression in both mutant and wild-type mice (p = 0.006). These results indicate that increased Pb concentrations in the brain could cause an upregulation of intracellular concentrations of dopamine by increasing dopamine synthesis (TH) and reuptake (DAT). Third, Hfe mutation decreased D1DR expression (p < 0.001; Fig. 6C) and Pb showed no effect on D1DR. Finally, D2DR expression was elevated by Hfe mutation (p = 0.004; Fig. 6D) and not by Pb exposure. Taken together, these results demonstrate that both Pb exposure and Hfe mutation can affect dopaminergic neurotransmission by changing dopamine turnover and dopamine receptors.
Fig. 6. Effect of Pb exposure via drinking water on dopamine-related protein expression in the brain of Hfe-mutant mice. Western blot analysis was performed to analyze expression levels of tyrosine hydroxylase (A), dopamine transporter (B), dopamine D1 receptor (C) and dopamine D2 receptor (D) in the brain from mice exposed to Pb at 1.6mg/ml by drinking water (n = 4~8 per group). Open and closed bars represent water and Pb treatment, respectively. All data were presented as mean ± SEM. All the P values were calculated by two-way ANOVA, followed by post-hoc comparisons. WT, wild-type; MUT, Hfe-mutant.

DISCUSSION
Lead exposure has raised huge concerns in public health due to its abundance in daily life (i.e. paint, plumbing, food cans) and its adverse effect on humans. Lead neurotoxicity has been characterized by several neurotoxic effects (e.g., reduction of intellectual abilities and memory) and psychiatric disorders (anxiety, depression and phobia). Defective Hfe function is associated with increased iron uptake resulting from elevated levels of metal transporters, which can also absorb several other divalent metals (e.g., lead and cadmium). For this reason, it is important to explore if Hferelated hemochromatosis could influence Pb uptake and modulate metal-associated neurotoxicity. Moreover, because both Pb exposure and Hfe mutation are implicated in the occurrence of autism and emotional dysfunction (1,3,4,12), in the present study we investigated the potential interaction effects of these two risk factors on repetitive behavior using Hfe-mutant mice, a mouse model of Hfe-related hemochromatosis in humans (13,14). Our data support the idea that both Pb exposure and Hfe mutation increase repetitive behavior in mice. In addition, we demonstrated that Pb had a greater effect on wild-type mice, whereas Pb-exposed Hfe-mutant mice did not show significant difference in repetitive behavior from non-exposed Hfe-mutant mice. Our model suggests that individuals with hemochromatosis caused by HFE mutations may be less susceptible to emotional problems resulting from Pb exposures and further provides a therapeutic basis for Pb-related psychiatric disorders.
While Pb exposure did not alter iron levels in the liver, Hfe mutation significantly increased hepatic iron concentration. The latter is consistent with others’ findings that defective Hfe enhances intestinal iron uptake (17). Interestingly, there was a significant difference in the liver Pb accumulation between Hfe-mutant and wild-type mice after Pb exposure for 6 wks. This could be due to increased ferritin expression in response to elevated hepatic iron upon Hfe mutation, since both in vitro and in vivo studies have shown that ferritin can bind various metals including Pb (18). Alternatively, up-regulated activities of DMT1 and other metal transporters in the intestine upon hemochromatosis (17) could have increased Pb absorption. However, there was no significant difference in brain Pb content between Hfe-mutant and wild-type mice when exposed to Pb, whereas iron level was elevated in Hfe-mutant mice. This suggests the existence of different transport mechanisms for the uptake of Pb in the liver and the brain. We speculate two possibilities: a dose of 1.6 mg/ml that we used in the current study could be sufficient to saturate brain transport but may not be enough for the storage in the liver. Alternatively, it is possible that Pb clearance out of the brain is also enhanced in Hfe mutation. Future studies should differentiate these kinetic processes along with dose-response relationships.
The marble-burying and nestlet-shredding tests provide useful information about repetitive behaviors (19,20). The behavioral parameters that we obtained in the present study reveal that both Pb exposure and Hfe mutation can alter repetitive behavior and further indicate that there is an interaction effect between the two conditions. Although nestletshredding score (or percent) has been well-recognized as a measurement of repetitive behavior, its performance is also influenced by other factors, such as motivation, anxiety, and circadian rhythm (21). The latency to the beginning of shredding reflects the levels of exploratory activity of animals and their motivation to perform innate behavior (21). Therefore, these factors other than repetitive behavior might have masked a potential effect of Hfe mutation on both nestlet-shredding score and the latency to the beginning of shredding. Our results demonstrated that the latency increased with Pb exposure in wild-type mice but not in Hfe-mutant mice, indicating a different degree of motivation among these animal groups. On the other hand, marble-burying performance has been shown to be directly related to repetitive behavior (19), which was modified by Hfe mutation and Pb effect in our study. Since repetitive behavior is commonly observed in several psychological disorders, such as autism and schizophrenia (20,22), behavioral methods that are more specific to social or sensorimotor behavior and less dependent on exploration or activity (e.g., reciprocal social interactions, social preference tests, prepulse inhibition) will examine more reliably the effect of Pb and Hfe mutation on these psychiatric conditions (22).
Dopaminergic pathway plays a key role in regulating emotional behavior and it has been well-documented that Pb exposure affects dopamine and serotonin metabolism (23,24). Pb selectively decreases dopamine binding to dopamine receptors and DAT in nucleus accumbens, whereas dopamine agonist treatments restore receptor and DAT binding (24). This suggests that Pb exposure could predispose neurodegenerative diseases associated with dopaminergic dysfunction. Pb intoxication is also associated with decreased levels of molecules involved in monoamine turnover, including vesicular monoamine transporter 2, serotonin, and dopamine-β-hydroxylase, in the auditory brainstem (23). In our study, the expression of both TH and DAT increased upon Pb exposure in the brain from wild-type mice, while that of D1DR and D2DR was unchanged. Elevated DAT density could result from a compensatory mechanism in response to the increased production of dopamine, as evidenced by elevated TH level, and/or decreased dopamine binding to DAT upon Pb exposure (24). With respect to Hfe, while iron deficiency has been shown to down-regulate brain DAT (25), iron overload Hfe-mutants in our study also exhibited lower levels of DAT compared with wildtype mice. Thus, any deviation from optimal iron levels may perturb DAT homeostasis, but likely via different mechanisms. In addition, while it was shown that both dopamine D1 and D2 receptors are involved in repetitive behavior (26), our results indicate that Hfe-mutant mice had reduced D1DR density in the brain, which could contribute to decreased dopaminergic activity. Moreover, both Pb exposure and D2DR gene polymorphism are associated with neurocognitive measures (27). Hence, it is plausible that repetitive behavior observed in Hfe mutation could be attributed to decreased D1DR and increased D2DR levels and/or any coordinated actions of these two receptors. Further study on the characterization of synaptic neurotransmitters would help to illustrate the effect of Hfe mutation and Pb exposure on monoaminergic signaling pathways.
To our knowledge, the current study is the first to characterize the neurotoxic effect of Pb exposure in the presence of defective Hfe, and our results also reveal that both Pb exposure and Hfe mutation modulate not only DAT, but also dopamine synthesis and receptors. Previously, a study demonstrated that exposures to Pb during pregnancy cause behavioral alterations, such as increased hyperactivity and impaired learning and memory in rats (3). Especially, the Pb level in the hair and nails of the children are found to be closely associated with the severity of autism (28). Importantly, increased Pb exposures can disrupt iron transport and metabolism (16), whereas both iron deficiency and iron overload (Hfe mutation) are associated with increased occurrences of autism (12,29). However, we observed that Pb exposure had no effect on brain iron levels, which prompts us to speculate that Pb-associated emotional changes are modified more likely by downstream effects of interaction between Pb and Hfe mutation on neurotransmission pathways rather than by direct influences of Hfe on metal transport. Nonetheless, we have not excluded the possibility that Pb exposure could alter iron levels by a region-specific manner with the whole brain iron levels unchanged. ICPMS analysis using individual brain regions will identify this effect. Alternatively, it is possible that Pb-induced neurotoxicity and altered emotional behavior could require an intrinsic function of Hfe, which is yet to be characterized, such that Hfe mutation does not exhibit repetitive behavior upon Pb exposure. Finally, these events could be mixed consequences in the downstream pathways triggered by combinatorial effects of metal exposure and adaptive changes in neurotransmission. Our finding of decreased Se levels in the brain as a result of Pb exposure in both genotypes could suggest that brain anti-oxidant system may be disrupted as a result of Pb exposure without Hfe effect. While we have characterized the dopaminergic neurotransmission pathway, a more comprehensive study is warranted to explore the exact relationship between Pb and Hfe in different brain regions along with other neurotransmitters in the context of emotional behavior.
This work was supported by the NIH R00 ES017781 (J.K.). The authors are grateful to Dr. Philip Larese-Casanova and Mr. Pooya Paydary (Northeastern University, Department of Civil & Environmental Engineering) for their help with the ICP-MS analyses. The authors also thank Dr. Timothy Maher and Dr. Alejandro Pino (MCPHS University, Boston, MA) for helpful advice on animal experiments.
References
- 1.Toscano C.D., Guilarte T.R. Lead neurotoxicity:from exposure to molecular effects. Brain Res. Rev. (2005);49:529–554. doi: 10.1016/j.brainresrev.2005.02.004. [DOI] [PubMed] [Google Scholar]
- 2.Kim S., Arora M., Fernandez C., Landero J., Caruso J., Chen A. Lead, mercury, and cadmium exposure and attention deficit hyperactivity disorder in children. Environ. Res. (2013);126:105–110. doi: 10.1016/j.envres.2013.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Moreira E.G., Vassilieff I., Vassilieff V.S. Developmental lead exposure: behavioral alterations in the short and long term. Neurotoxicol. Teratol. (2001);23:489–495. doi: 10.1016/S0892-0362(01)00159-3. [DOI] [PubMed] [Google Scholar]
- 4.Levin R., Grown M.J., Kashtock M.E., Jacobs D.E., Whelan E.A., Rodman J., Schock M.R., Padilla A., Sinks T. Lead exposures in U.S. Children, 2008: implications for prevention. Environ. Health Perspect. (2008);116:1285–1293. doi: 10.1289/ehp.11241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hu H., Shine J., Wright R.O. The challenge posed to children’s health by mixtures of toxic waste: the Tar Creek superfund site as a case-study. Pediatr. Clin. North Am. (2007);54:155–175. doi: 10.1016/j.pcl.2006.11.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Garrick M.D., Dolan K.G., Horbinski C., Ghio A.J., Higgins D., Porubcin M., Moore E.G., Hainsworth L.N., Umbreit J.N., Conrad M.E., Feng L., Lis A., Roth J.A., Singleton S., Garrick L.M. DMT1: a mammalian transporter for multiple metals. Biometals. (2003);16:41–54. doi: 10.1023/A:1020702213099. [DOI] [PubMed] [Google Scholar]
- 7.Hashmi N.S., Kachru D.N., Khandelwal S., Tandon S.K. Interrelationship between iron deficiency and lead intoxication (Part 2). Biol. Trace Elem. Res. (1989);22:299–307. doi: 10.1007/BF02916618. [DOI] [PubMed] [Google Scholar]
- 8.Pietrangelo A. Hereditary hemochromatosis—a new look at an old disease. N. Engl. J. Med. (2004);350:2383–2397. doi: 10.1056/NEJMra031573. [DOI] [PubMed] [Google Scholar]
- 9.Zhang A., Park S.K., Wright R.O., Weisskopf M.G., Mukherjee B., Nie H., Sparrow D., Hu H. HFE H63D polymorphism as a modifier of the effect of cumulative lead exposure on pulse pressure: the Normative Aging Study. Environ. Health Perspect. (2010);118:1261–1266. doi: 10.1289/ehp.1002251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lee S.H., Kim J.W., Shin S.H., Kang K.P., Choi H.C., Park K.U., Kim H.Y., Kang W., Jeong S.H. HFE gene mutations, serum ferritin level, transferrin saturation, and their clinical correlates in a Korean population. Dig. Dis. Sci. (2009);54:879–886. doi: 10.1007/s10620-008-0432-6. [DOI] [PubMed] [Google Scholar]
- 11.Nandar W., Connor J.R. HFE gene variants affect iron in the brain. J. Nutr. (2011);141:729S–739S. doi: 10.3945/jn.110.130351. [DOI] [PubMed] [Google Scholar]
- 12.Gebril O.H., Meguid N.A. HFE gene polymorphisms and the risk for autism in Egyptian children and impact on the effect of oxidative stress. Dis. Markers. (2011);31:289–294. doi: 10.1155/2011/605620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Liu Y., Lee S.Y., Neely E., Nandar W., Moyo M., Simmons Z., Connor J.R. Mutant HFE H63D protein is associated with prolonged endoplasmic reticulum stress and increased neuronal vulnerability. J. Biol. Chem. (2011);286:13161–13170. doi: 10.1074/jbc.M110.170944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nandar W., Neely E.B., Unger E., Connor J.R. A mutation in the HFE gene is associated with altered brain iron profiles and increased oxidative stress in mice. Biochim. Biophys. Acta. (2013);1832:729–741. doi: 10.1016/j.bbadis.2013.02.009. [DOI] [PubMed] [Google Scholar]
- 15.Berglund M., Lindberg A.L., Rahman M., Yunus M., Grandér M., Lönnerdal B., Vahter M. Gender and age differences in mixed metal exposure and urinary excretion. Environ. Res. (2011);111:1271–1279. doi: 10.1016/j.envres.2011.09.002. [DOI] [PubMed] [Google Scholar]
- 16.Molina R.M., Phattanarudee S., Kim J., Thompson K., Wessling-Resnick M., Maher T.J., Brain J.D. Ingestion of Mn and Pb by rats during and after pregnancy alters iron metabolism and behavior in offspring. Neurotoxicology. (2011);32:413–422. doi: 10.1016/j.neuro.2011.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Morgan E.H., Oates P.S. Mechanisms and regulation of intestinal iron absorption. Blood Cells Mol. Dis. (2002);29:384–399. doi: 10.1006/bcmd.2002.0578. [DOI] [PubMed] [Google Scholar]
- 18.Joshi J.G., Zimmeman A. Ferritin: an expanded role in metabolic regulation. Toxicology. (1988);48:21–29. doi: 10.1016/0300-483X(88)90055-8. [DOI] [PubMed] [Google Scholar]
- 19.Thomas A., Burant A., Bui N., Graham D., Yuva-Paylor L.A., Paylor R. Marble burying reflects a repetitive and perseverative behavior more than novelty-induced anxiety. Psychopharmacology (Berl) (2009);204:361–373. doi: 10.1007/s00213-009-1466-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Angoa-Pérez M., Kane M.J., Briggs D.I., Francescutti D.M., Kuhn D.M. Marble burying and nestlet shredding as tests of repetitive, compulsive-like behaviors in mice. J. Visualized Exp. (2013);24:50978. doi: 10.3791/50978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jirkof P., Fleischmann T., Cesarovic N., Rettich A., Vogel J., Arras M. Assessment of postsurgical distress and pain in laboratory mice by nest complexity scoring. Lab. Anim. (2013);47:153–161. doi: 10.1177/0023677213475603. [DOI] [PubMed] [Google Scholar]
- 22.Silveman J.L., Yang M., Lord C., Crawley J.N. Behavioural phenotyping assays for mouse models of autism. Nat. Rev. Neurosci. (2010);11:490–502. doi: 10.1038/nrn2851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fortune T., Lurie D.I. Chronic low-level lead exposure affects the monoaminergic system in the mouse superior olivary complex. J. Comp. Neurol. (2009);513:542–558. doi: 10.1002/cne.21978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Pokora M.J., Richfield E.K., Cory-Slechta D.A. Preferential vulnerability of nucleus accumbens dopamine binding sites to low-level lead exposure: time course of effects and interactions with chronic dopamine agonist treatments. J. Neurochem. (1996);67:1540–1550. doi: 10.1046/j.1471-4159.1996.67041540.x. [DOI] [PubMed] [Google Scholar]
- 25.Bianco L.E., Wiesinger J., Earley C.J., Jones B.C., Beard J.L. Iron deficiency alters dopamine uptake and response to L-DOPA injection in Sprague-Dawley rats. J. Neurochem. (2008);106:205–215. doi: 10.1111/j.1471-4159.2008.05358.x. [DOI] [PubMed] [Google Scholar]
- 26.Taylor J.L., Rajbhandari A.K., Berridge K.C., Aldridge J.W. Dopamine receptor modulation of repetitive grooming actions in the rat: potential relevance for Tourette syndrome. Brain Res. (2010);1322:92–101. doi: 10.1016/j.brainres.2010.01.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kordas K., Ettinger A.S., Bellinger D.C., Schnaas L., Téllez Rojo M.M., Hernández-Avila M., Hu H., Wright R.O. A dopamine receptor (DRD2) but not dopamine transporter (DAT1) gene polymorphism is associated with neurocognitive development of Mexican preschool children with lead exposure. J. Pediatr. (2011);159:638–643. doi: 10.1016/j.jpeds.2011.03.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lakshmi Priya M.D., Geetha A. Level of trace elements (copper, zinc, magnesium and selenium) and toxic elements (lead and mercury) in the hair and nail of children with autism. Biol. Trace Elem. Res. (2011);142:148–158. doi: 10.1007/s12011-010-8766-2. [DOI] [PubMed] [Google Scholar]
- 29.Latif A., Heinz P., Cook R. Iron deficiency in autism and Asperger syndrome. Autism. (2002);6:103–114. doi: 10.1177/1362361302006001008. [DOI] [PubMed] [Google Scholar]