

Abstract
Background. The regulation of lipid biosynthesis is essential in photosynthetic eukaryotic cells. Thus far, no regulatory genes have been reported in the lipid metabolism pathway. Plant CONSTANS (CO) gene regulates blooming by participating in photoperiod and biological clock. Apart from regulating photoperiod, the Chlamydomonas CO gene also regulates starch content. Results. In this study, the results showed that, under HSM-S condition, cells accumulated more lipids at short-day conditions than at long-day conditions. The silencing of the CrCO gene via RNA interference resulted in an increase in lipid content and an increase in triacylglyceride (TAG) level by 24.5%. CrCO RNAi strains accumulated more lipids at short-day conditions than at long-day conditions. The decrease in CrCO expression resulted in the increased expression of TAG biosynthesis-related genes, such as DGAT2, PAP2, and PDAT3, whereas CIS and FBP1 genes showed a decrease in their mRNA when the CrCO expression was suppressed. On the other hand, the overexpression of CrCO resulted in the decrease in lipid content and TAG level. Conclusions. The results of this study revealed a relationship between CrCO gene and lipid metabolism in Chlamydomonas, suggesting that increasing oil by suppressing CrCO expression in microalgae is feasible.
1. Introduction
Given the risk of depleting traditional fossil fuels such as oil and coal, people have now realized the urgent need to develop renewable energy sources. Thus, the use of biodiesel from microalgae as an important source of renewable energy and as a vital alternative energy source of future fossil fuel has attracted increasing attention from scholars and enterprises. Autotrophic microalgae convert solar energy into biomass energy, which fix a large amount of CO2 and store the biomass energy as lipids in cells in specific strains. For a long time, studies on the lipid metabolism pathway of microalgae lagged behind studies on most crops such as rice, wheat, and corn. More studies have concentrated on the mechanism of lipid metabolism and high-density culture, which are significant to the genetic improvement of high quality strains and the industrialization of aquaculture.
Photoperiodism, a ubiquitous feature of plants, is the response of plants to the relative length of day and night. Photoperiodism is a key factor that affects plant conversion from vegetative growth to reproductive growth as well as the plant's flowering time, which is regulated by a large and complex genetic network [1]. CONSTANS (CO) is an important gene that regulates both plant photoperiod and flowering time [2, 3]. CO homologous genes were identified in a few plants, such as Hd1 of rice [4], TaHd1 of Tritivum aestivum [5], StCO of Solanum tuberosum [6, 7], and CrCO of Chlamydomonas reinhardtii [8], by screening the library and by homology-based cloning with the CO gene sequence of Arabidopsis. Meanwhile, CO homologous genes have been cloned in different species, including rape plants, eastern cottonwood [9], barley [10], tomato [11], pharbitisnil [12], spider flower [13], radish [14], pea [15], and perennial ryegrass [16].
CO genes were found in multiple copies in most plant genomes that have been studied. A total of 17 homologous genes were found in Arabidopsis [17], 16 in rice [18], three in Solanum lycopersicum [19], four in Brassica napus [20], and two in Picea abies [21]. CO is a zinc finger transcription factor that contains B-box and CCT conservative domains and a variable region in the middle [22–25]. The N-terminal of CO has two sequential B-box domains designated as double B-box (DBB). The B-box domain of CO interacts with proteins [26], in which the cysteine and histidine that bind to zinc ion are highly conserved. Conservative amino acid mutations resulted in a delay in flower production [27]. Ben-Naim et al. used the B-box of TCOL1 as bait to analyze its interaction with immunophilin and/or other proteins that contain B-box and determined that B-box mainly functions with proteins [19]. The C-terminal of the CO protein is composed of 70 to 80 amino acids, in which approximately 40 are highly conserved in the CO family [27, 28]. Moreover, the CCT domain of CO contains a nuclear localization signal that interacts with COP1 of the ubiquitin ligase [29]. In the present study, the mechanism of combining CO with DNA is unclear, although speculation indicated that CO binds to DNA via its CCT domain to form a complex compound.
Aside from the DBB zinc finger and the CCT domain, the B-box family of Arabidopsis also contains DBB homologous CO subfamilies, which have two DBB domains in the N-terminal that are separated by 8 to 15 amino acids, whereas the C-terminal does not contain any CCT domains. DBB homologous subfamilies are encoded by eight genes, namely, DBB1a (At2g21320), DBB1b (At4g38960), DBB2 (At4g39070), DBB3 (At1g78600), DBB4 (At4g10240), STO (At1g06040), STH (At2g31380), and STH2 (At1g75540) [30]. To date, four of the eight genes have been found to be involved in light-mediated plant growth and development. STO has a function in plant salt tolerance and negatively regulates phytochrome and blue light signal transduction pathway [31]. STH has a similar function to STO, and they both interact with ubiquitin ligase COP1 [32]. Both DBB3 and STH2 participate in regulating plant hypocotyl elongation, early chloroplast development, and anthocyanin accumulation and in the positive regulation of deetiolation in Arabidopsis [32–34].
Functions of CO in microalgae have also been reported. Serrano et al. determined that CrCO regulates light cycle in Chlamydomonas, and both knockdown and overexpression of CrCO changed the diurnal cycle of the cells. Thus, related gene expression and physiological functions are regulated [8]. Ral et al. determined that CrCO participates in starch synthesis by regulating the GBSSI gene [35]. Chlamydomonas CO gene reported by Serrano et al. was used to perform a BLASTP search in the Chlamydomonas database Phytozome, and only one homology, g6302, was found (100% identified). Other homologous proteins containing the B-box domain were not detected. Nevertheless, seven genes that encode proteins containing the CCT domain were homologous to the CrCO gene. Based on the number and conservative property of B-box, 17 genes of the CO gene family can be divided into three subgroups in Arabidopsis [27, 28]. In addition, eight genes encoded proteins that are homologous to the DBB protein subfamily, namely, DBB1 to DBB4, STO, STH, and STH2 [30]. These genes regulate multiple physiological functions in plants, such as bloom, biological clock, photoperiod, growth and development (DBB1 to DBB4), salt-tolerance (STO), light signal transduction, hypocotyl elongation (DBB3 and STH2), early formation of the chloroplast, and accumulation of anthocyanin [31–34]. Compared with the mass of genes encoding the DBB subfamily of the CO protein in Arabidopsis, only one gene encoding the CO protein was found in Chlamydomonas, which indicates that higher plants and single CO gene in Chlamydomonas had complex functions such as the regulation of flower production and multicellular development.
To date, CrCO have not been proven to be involved in regulating lipid metabolism. In the field of microalgae lipid metabolism mechanism research, studies have concentrated on genes involved in the lipid synthesis pathway and photosynthetic carbon metabolism pathway, not on regulating genes. The present study discussed the function of CrCO gene in lipid accumulation in microalgae cells via the knockdown and overexpression of CrCO in Chlamydomonas. Furthermore, the relationship between CrCO gene and lipid metabolism was revealed by analyzing lipid accumulation at adverse -P and -S conditions and at long-day and short-day conditions.
2. Results and Analysis
2.1. Chlamydomonas Accumulated More Lipids in Short Day (SD) than in Long Day (LD) under -S Conditions
C. reinhardtii CC425 was inoculated in a 50 mL Erlenmeyer flask containing an HSM medium and grown until the mid-log phase (2 × 10e 6). The cells were collected after centrifugation and then equally divided into three parts. Each triplet was inoculated in 30 mL medium of HSM, HSM-S, and HSM-P and grown at LD condition (16 h light and 8 h dark). Identical triplets were grown in SD conditions (8 h light and 16 h dark). The cells grew slightly slower in SD conditions than in LD conditions in HSM. Moreover, cell proliferation was greatly reduced in HSM-S compared with that in HSM in LD or SD conditions, whereas the length of daylight did not affect cell growth in HSM-P (Figures 1 and 2). The lipid contents in HSM in SD conditions were no significant difference compared to the lipid contents in HSM in LD conditions, similarly, the lipid contents in HSM-P in SD conditions were no significant difference compared to the lipid contents in HSM-P in LD conditions (Figures 1 and 2), whereas the lipid content significantly increased in SD conditions compared with that in LD conditions in HSM-S.
Figure 1.

Growth curve and lipid contents of C. reinhardtii CC425 cultivated in LD or SD conditions. (a) Growth curve of C. reinhardtii CC425 cultivated in LD conditions and at -S or- P limitation conditions. (b) Growth curve of C. reinhardtii CC425 cultivated in SD conditions and at -S or -P limitation conditions. (c) Lipid contents of C. reinhardtii CC425 cultivated in LD conditions and at -S or -P limitation conditions. (d) Lipid contents of C. reinhardtii CC425 cultivated in SD conditions and at -S or -P limitation conditions. HSM, cells cultivated in the HSM medium; HSM-S, cells cultivated in the S-free HSM medium; and HSM-P, cells grown in the P-free HSM medium.
Figure 2.

Microscopic observations of C. reinhardtii under LD or SD conditions after four days of cultivation. Algal cell was observed using a Zeiss fluorescence microscope (10 × 25) after staining with Nile red dye. Orange fluorescence indicates that the oil droplet is mainly composed of TAG. LD conditions (16 h day time and 8 h dark time); SD conditions (8 h day time and 16 h dark time). HSM, cells cultivated in the HSM medium; HSM-S, cells cultivated in the S-free HSM medium; and HSM-P, cells grown in the P-free HSM medium. The scale bar indicated in the figure is 3 μm.
2.2. Analysis of the Relationship between CrCO mRNA Level and Lipid Accumulation at -S and -P Conditions
The CrCO mRNA level remarkably decreased in HSM-P compared with that in HSM. Moreover, the CrCO mRNA level decreased in HSM-S compared with that in the control sample (Figure 3). The lipid content remarkably increased in HSM-S compared with that in the control sample (Figure 1). The results illustrate that the CrCO mRNA level was negatively correlated with lipid accumulation at -S conditions.
Figure 3.
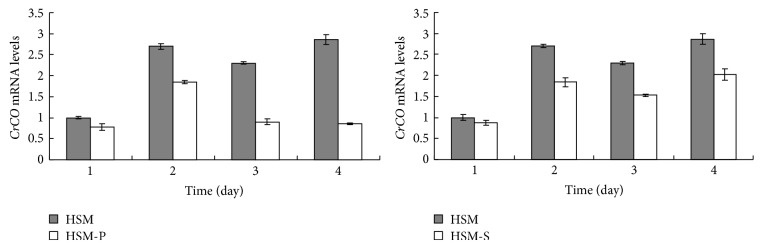
Abundance of mRNA of CrCO in HSM, HSM-N, and HSM-P media. mRNA levels of C. reinhardtii CC425 samples grown in the indicated medium for 1, 2, 3, or 4 d in full daylight were analyzed via RT-PCR. HSM, cells cultivated in the HSM medium; HSM-S, cells cultivated in the S-free HSM medium; and HSM-P, cells grown in the P-free HSM medium.
2.3. Silencing of CrCO Gene Increases TAG Content in C. reinhardtii
Approximately, 1230 bp full-length CDS of CrCO DNA fragment was amplified via PCR and cloned into pMD18T and sequenced thereafter. This fragment exhibited 100% homology with the Chlamydomonas CO gene (g6302.t1). To determine the relationship between CrCO expression and lipid accumulation, the effects of the artificial silencing of CrCO gene on the lipid content of C. reinhardtii were examined. Based on the CrCO (g6302.t1) sequences of the gene retrieved from the Phytozome C. reinhardtii database (http://www.phytozome.net/), primers used to amplify the fragment of the coding region of CrCO, were designed. The DNA fragments were subcloned and then used to generate CrCO RNAi constructs pMaa7IR/CrCO IR. More than 100 positive transformants were obtained after transforming the silencing construct into C. reinhardtii CC425. Three transgenic algae were selected to measure the lipid content and mRNA levels of the target gene. Strains transformed with the vector pMaa7IR/XIR were used as control samples. In cells harboring the CrCO construct, analysis results of the transgenic lines via the Nile red fluorescence method indicate an increase in the lipid content by 13.5% to 35.2% (Figure 4(b)) after 10 days of cultivation. The TAG level of the transgenic strain CrCO RNAi18 increased by 24.5% compared with that in the control sample (Figure 4(c)). To evaluate the effectiveness of the RNAi construct, the abundance of the target gene-specific mRNA in transgenic algae was analyzed via real-time PCR. The CrCO mRNA abundance decreased by 90.4% to 95.2% (Figure 4(d)), which indicates the high-efficiency silencing by these constructs.
Figure 4.

The biomass, lipid content detected by Nile red fluorescence method and TAG level, and the mRNA abundance of CrCO in CrCO RNAi transgenic C. reinhardtii. Cr.CC425, C. reinhardtii CC425; Cr.Maa7, pMaa7IR/XIR transgenic algae strain; CrCO RNAi18 (31, 87), and pMaa7IR/CrCOIR transgenic algae strains. Statistical analysis was performed using SPSS statistical software. Significance is indicated as * P < 0.05, ** P < 0.01.
Subsequently, the mRNA levels of phospholipid:diacylglycerol acyltransferase PDAT3, acyl-CoA:diacylglycerol acyltransferase (DGAT2), phosphofructokinase (PFK2), fructose-1,6-bisphosphatase (FBP1), citrate synthase (CIS), and phosphatidate phosphatase (PAP2) genes were measured in transgenic strain CrCO RNAi18. Genes such as PDAT3, DGAT2, and PAP2 are directly related to lipid synthesis, which increase the mRNA levels in transgenic strain compared with those in nontransgenic C. reinhardtii CC425 and Maa7IR/XIR transgenic algae. On the other hand, the CIS gene, a key enzyme in tricarboxylic acid cycle, and the FBP1 gene, which is found in gluconeogenesis, showed a decrease in their mRNA in the transgenic strain (Figure 5). The results indicate the regulation of CIS and FBP1 genes by CrCO, which exhibits negative effects on the regulation of the expression of lipid biosynthesis genes, such as PDAT3, DGAT2, and PAP2. The FBP1 and CIS genes exhibited decreased mRNA in the transgenic strain, CrCO RNAi18, and a glycolysis enzyme, PFK2, exhibited increased mRNA in the transgenic strain, which indicated that more carbon is introduced to the fatty acid and lipid synthesis (Figure 5).
Figure 5.

mRNA abundance of PDAT3, DGAT2, PFK1, FBP1, CIS, and PAP2 in CrCO RNAi transgenic algae strain, CrCO RNAi18. Cr CC425, C. reinhardtii CC425; Cr.Maa7, pMaa7IR/XIR transgenic algae strain; CrCO RNAi18, and pMaa7IR/CrCOIR transgenic algae strain number 18.
Results similar to the above discussion were obtained via Nile red staining. More oil droplets were found in CrCO RNAi18 transgenic algae compared with those in pMaa7IR/XIR transgenic algae, as determined via microscopic analysis (Figure 6). This result indicates an increase in cell lipid content via the regulation of CrCO gene expression.
Figure 6.

Microscopic observations of CrCO transgenic C. reinhardtii. After six days of cultivation in full daylight and HSM medium, more oil droplets of CrCO RNAi transgenic algae were found. Cr.Maa7, pMaa7IR/XIR transgenic algae strain. CrCO RNAi18, and pMaa7IR/CrCOIR transgenic algae strain number 18. The scale bar indicated in the figure is 2 μm.
Considering that CrCO gene regulates photoperiod, would the lipid content of cells at SD or LD conditions change via RNAi-initiated CrCO knockdown? The results indicate that transgenic strains exhibited higher lipid content at SD condition than at LD condition (Figures 7(a) and 7(b)). The lipid content was measured at both -S and -P conditions in transgenic strains, and the results indicate that more lipids were accumulated at SD condition than at LD condition (Figures 7(c)–7(f)). Thus, more lipids are produced in RNAi transgenic strains compared with those in the control sample regardless of the cultivation conditions.
Figure 7.

Lipid contents of CrCO transgenic strains cultivated at LD or SD conditions. (a) Lipid contents of CrCO transgenic strains cultivated under SD conditions in the HSM medium. (b) Lipid contents of CrCO transgenic strains cultivated under LD conditions in the HSM medium. (c) Lipid contents of CrCO transgenic strains cultivated under SD conditions in the HSM-P medium. (d) Lipid contents of CrCO transgenic strains cultivated under LD conditions in the HSM-P medium. (e) Lipid contents of CrCO transgenic strains cultivated under SD conditions in the HSM-S medium. (f) Lipid contents of CrCO transgenic strains cultivated under LD conditions in the HSM-S medium. Cr.Maa7, pMaa7IR/XIR transgenic algae strain; CrCO RNAi18 (31, 87), and pMaa7IR/CrCOIR transgenic algae strains.
2.4. Overexpression of CrCO Reduced the Lipid Content of C. reinhardtii
The increase in lipid content caused by the RNAi silencing of CrCO suggests the effect of the expression of these genes on the biosynthesis of triglycerides in C. reinhardtii. Thus, the capacity of CrCO overexpression to reduce the lipid content of C. reinhardtii was determined. Vector pCAMCO, which expresses CrCO gene from the CAMV 35S promoter, was introduced into C. reinhardtii. The lipid contents and growth rate of three randomly selected transgenic algae were determined in each transgenic algae line. The overexpression of the CrCO gene increased the growth rate of the algae in the early stages from day two to day four (Figure 8(a)). Moreover, the overexpression of CrCO decreased the lipid content of the transgenic algae compared with that in the control pCAMBIA1302 transgenic algae lines. For example, six days after the growth of algae in the HSM medium in full daylight, the lipid contents of CrCO-overexpressing transgenic lines decreased by 26.2% to 36.0%, as determined via the Nile red fluorescence method (Figure 8(b)). The TAG level of the transgenic strain, pCACO64, decreased by 19.4% compared with the TAG level of the control sample (Figure 8(c)). Compared with the mRNA levels of pCAMBIA1302 transgenic strains, the mRNA levels of CrCO increased by 27 to 29 times (Figure 8(d)). In summary, the overexpression of CrCO gene decreases lipid synthesis in cells. Decreased lipid content was also observed via Nile red dye staining (Figure 9). Fewer oil droplets were found in CrCO-overexpressed transgenic algae compared with those in the control sample.
Figure 8.

Biomass and lipid content detected via the Nile red fluorescence method, TAG level, and mRNA abundance of CrCO in CrCO-overexpressed transgenic C. reinhardtii. Cr.CC425, C. reinhardtii CC425; Cr.pCAMBIA, pCAMBIA1302 transgenic algae strain; pCACO11 (64, 91), and pCAMCO transgenic algae strains. Statistical analysis was performed using SPSS. Significance is indicated as * P < 0.05, ** P < 0.01.
Figure 9.

Lipid content in transgenic algae line detected via Nile red staining. After six days of cultivation in the HSM medium, little oil droplets of CrCO transgenic algae were found. Cr.pCAMBIA, pMCAMBIA1302 transgenic algae strain; pCACO64, and pCAMCO transgenic algae strain number 64. The scale bar indicated in the figure is 2 μm.
3. Discussion
Chlamydomonas CO has a typical DBB zinc finger domain and a CCT domain in the C-terminal, which has been proven to regulate photoperiod. Moreover, CrCO has a function in controlling starch content in Chlamydomonas by regulating the effects of GBSSI gene expression. In this study, CrCO was shown to regulate lipid accumulation. The mRNA level of the CO gene of Chlamydomonas decreased at -S condition via digital gene expression (DGE) profiling. Considering the increase in the lipid content at -S condition in cells, knockdown and overexpression of the CO gene in Chlamydomonas cells were performed to determine the relationship between the CO gene and lipid accumulation. The results indicate that the CO gene was closely correlated to lipid accumulation because the silencing of the CO gene results in an increase in the lipid content and the overexpression of the CO gene results in a decrease in the lipid content. In addition, the silencing of the CO gene caused the mRNA level of genes to contribute to lipid synthesis, such as DGAT2, DGAT1, and PAP2, which were optimized at daytime. Therefore, we hypothesized that CrCO facilitates indirect lipid production by regulating gene-encoding enzymes in the lipid synthesis pathway, namely, DGAT1, DGAT2, and PAP2. The SD condition exhibited positive effects on lipid accumulation compared with the LD condition, which was more significant in the -S condition. This conclusion was consistent with the findings that lipid content changes in CO knockdown transgenic algae strains. In future studies, emphasis must be given on how the CO gene works, and the regulation path way of CO to lipid synthesis gene, such as those of DGAT1, DGAT2, and PAP2.
4. Materials and Methods
4.1. Bioinformatics, Algal Strain, Cultivation Conditions, and Biomass Assay
The information on the Chlamydomonas CO gene (g6302) was obtained from the Phytozome V9.1 Chlamydomonas database (http://www.phytozome.net/). C. reinhardtii CC425 (mt) was purchased from the Chlamydomonas Genetics Center at Duke University. The cells grown on tris-acetate-phosphate (TAP) agar plate were inoculated into 100 mL Erlenmeyer flasks containing 50 mL of HSM, P-deficient HSM (HSM-P), and S-deficient HSM (HSM-S) media [36]. The HSM medium was composed of NH4Cl (0.500 g·L−1), MgSO4 ·7H2O (0.020 g·L−1), CaCl2 ·2H2O (0.010 g·L−1), K2HPO4 (1.440 g·L−1), KH2PO4 (0.720 g·L−1), NaAc (2.000 g·L−1), H3BO3 (0.001 g·L−1), MnCl2 ·4H2O (0.005 g·L−1), ZnSO4 ·7H2O (0.022 g·L−1), FeSO4 ·7H2O (0.005 g·L−1), CoCl2 ·6H2O (0.002 g·L−1), Na2Mo7O24 ·4H2O (0.002 g·L−1), and Na2 ·EDTA (0.050 g·L−1). The HSM-P medium contained K2HPO4 and KH2PO4, which can be replaced with KCl. The HSM-S medium contained MgSO4 ·7H2O, which can be replaced with MgCl2 ·6H2O. Generally, cultures were maintained in an incubator shaker at a rate of 230 rpm at 25°C and then exposed to continuous illumination at a light intensity of 150 μmol·m−2 ·s−1. The samples tested in LD conditions were incubated in a light time of 16 h at daytime and 8 h at dark, whereas samples tested in SD conditions were incubated in a light time of 8 h at daytime and 16 h at dark.
Biomass concentration (g/L) was determined by measuring the optical density of the samples at 490 nm (OD490), as described by an earlier study [37]. To generate the standard curve, a series of C. reinhardtii CC425 samples of different biomass concentrations was collected. The OD490 value and cell dry weight were gravimetrically determined using dried cells to plot the standard curve of OD490 versus biomass concentration (g/L). The samples were diluted to appropriate ratios to ensure that the measured OD490 values ranged from 0.15 to 0.75, if applicable. The biomass concentration was then calculated using the following formula: cell dry weight (g/L) = 0.7444∗OD490 − 0.0132 (Supplementary data Figure 1 available online at http://dx.doi.org/10.1155/2014/715020).
4.2. Lipid Content Analysis
Nile red fluorescence method and GC/MS were performed to determine lipid and TAG levels [38, 39]. The algal cells were directly stained with 0.1 mg/mL Nile red for 10 min, and fluorescence was then measured at excitation and emission wavelengths of 470 and 570 nm, respectively. The fluorescence value was calculated using the equation: FD (470/570) = (A2 − A1), where A2 is the fluorescence value of the algal cells after staining with Nile red and A1 is the fluorescence of algal cells before staining (Supplementary data Figure 2). Total lipid extraction was carried out according to a modified method. Logarithmic-phase algal cells were collected after centrifugation and extracted using an extraction buffer (methanol : chloroform : methanoic acid, 2 : 1 : 0.1), 1 M KCl, and 0.2 M H3PO4. The lipids were obtained after centrifugation at 13780 ×g for 3 min. For TAG separation, Si60 silica TLC plates for thin-layer chromatography (TLC) were used. The TLC plates were immersed in 0.15 M (NH4)2SO4 for 30 s and stored in an airtight container for two days. The plates were then placed in an oven at 120°C for 2.5 h and cooled at room temperature. The samples were then added with N2 flow. TAGs were observed on TLC plates via iodine staining. Lipid analysis was conducted as previously described. Fatty acid methyl esters derived from TAG were analyzed via GC/MS [40]. For microscopic assay, images were acquired using a Nikon 80i Fluorescence Microscope after the cells were stained with Nile red. Nile red signals were captured at an excitation wavelength of 480 nm, and emission was obtained between 560 and 600 nm [41–43]. A total of 30 cell lipid droplets from each algal strain were examined to determine the difference between the lipid contents.
4.3. RNA Extraction
Total RNA was prepared as described by Li et al. with modification [44]. Cells from 10 mL of cultivated algae were collected after centrifugation at 10000 ×g for 1 min. After a series of extractions using phenol-chloroform, nucleic acids were precipitated with two volumes of absolute ethanol and then washed with 75% ethanol. Finally, the air-dried pellet was dissolved in 40 μL of RNase-free water. RNA concentration was determined via spectrophotometry, and the integrity was examined via agarose gel electrophoresis.
4.4. Cloning of the CrCO Gene
The first strand of cDNA was synthesized using SuperScript III Reverse Transcriptase (Invitrogen, USA) according to the manufacturer's instructions. A fragment of the CrCO gene was amplified via polymerase chain reaction (PCR) by using primers CrCOL: 5′-ATGTCGAGTTGCGTCGTGTGCG-3′ and CrCOR: 5′-TTAGCACTCAGCGTCCAGGACCTCG-3′. PCR reactions were performed in a final volume of 25 μL containing 1× PCR reaction buffer, 2 mM MgCl2, 0.4 μmol of each primer, 0.25 mM dNTPs, 1 μL of DMSO, 0.5 M betaine, and 0.5 U Taq DNA polymerase (Promega, USA) according to the following program: 4 min at 95°C; 35 cycles of denaturation for 40 s at 95°C, annealing for 40 s at 58°C, and elongation for 20 s at 72°C; 10 min at 72°C. After purification using the EZ-10 Spin Column DNA Gel Extraction Kit (BBI, Canada), the DNA was inserted into vector pMD18-T following the manufacturer's instructions (TaKaRa, Japan). The resulting plasmid was designated as pMD18T-CrCO. The sequences of the cloned CrCO gene were verified via double-stranded sequence analysis (Shanghai Sangon Biological Engineering Technology & Services Co., Ltd).
4.5. Construction of the RNAi Vector against the CrCO Gene
A fragment of C. reinhardtii 18S gene was amplified with primers 5′-CGAACTTCTGCGAAAGCAT-3′ and 5′-TCAGCCTTGCGACCATACT-3′ and then inserted into pMD18-T to produce pMD18T-18S. The fragment of CrCO and its reverse complementary sequences were amplified via PCR by using pMD18T-CrCO as a template and the following primers: CrCORNAiL: 5′-AGCTGCTACGCACGAGACCG-3′ and CrCORNAiR: 5′-GCCCATGTCGAGCCAGTTGT-3. The PCR fragment was then digested with KpnI/BamHI and HindIII/SalI and was inserted into the corresponding cloning sites of pMD18T-18S to yield pMD18-CrCOF-18S-CrCOR, which contained an inverted repeat sequence of CrCO (CrCO IR). pMD18-CrCOF-18S-CrCOR was double-digested with KpnI and HindIII to obtain CrCO IR. Finally, the CrCO IR was inserted as a blunt-end fragment into EcoRI-digested pMaa7/XIR to yield pMaa7IR/CrCO IR.
4.6. Construction of Overexpression Vector of CrCO Gene for Chlamydomonas
To construct the overexpression vector of the CrCO gene, the coding sequence of CrCO was amplified via PCR by using pMD18T-CrCO as a template and primers 5′-AAAGATCTAATGTCGAGTTGCGTCGTGTG-3′ and 5′-AAACTAGTTTAGCACTCAGCGTCCAGGA-3′. The fragment was digested with NcoI/SpeI and inserted into similarly digested pCAMBIA1302 to produce pCAMCO, which allows the overexpression of CrCO.
4.7. Transformation of Chlamydomonas
The transformation of C. reinhardtii strain CC425 was performed as described by Kindle [45]. C. reinhardtii cells were grown in a TAP medium to a cell density of (1 to 2) × 106 cells/mL. The cells were collected after centrifugation, washed twice, and resuspended in the TAP medium to a cell density of approximately 1 × 108 cells/mL. Plasmid DNA was introduced into the cells via the glass bead procedure. In each case, 2 μg of plasmid DNA was included in a mixture containing 400 μL of cells, 100 μL of 20% polyethylene glycol, and 300 mg of sterile glass beads. The reaction was mixed for 15 s on a benchtop vortex. To allow the induction of RNAi or gene expression, the cells were allowed to recover for 1 d before plating onto selective media. RNAi transformants were selected on the TAP medium containing 1.5 mM L-tryptophan, 5 μg/mL paromomycin, and 5 μM 5-FI. pCAMCO transformants were selected on the TAP medium containing 50 μg/mL hygromycin. The plates were incubated under dim light (approximately 50 μmol·m−2 ·sec−1 photosynthetically active radiation). The isolated transgenic strains were kept at a constant selective pressure.
4.8. Quantitative Real-Time PCR
The samples were subjected to real-time PCR analysis as described by Fei and Deng [46]. RNA was extracted using TRIzol reagent (Sangon Biotech, Shanghai, China). Single-strand cDNA was obtained using 100 ng of the RNA sample and a random primer at 65°C for 5 min, 25°C for 5 min, and 42°C for 50 min in a Bio-Rad iScript select cDNA synthesis kit. Real-time PCR was conducted in a BioRad iCycler iQ real-time PCR detection system by using SYBR Green as the fluorescent dye. Each reaction was conducted in a final volume of 25 µL with the following components: 0.2 pmol of each primer, 1 µL of cDNA, and 12.5 µL of SYBR Green Mix (Invitrogen SYBR Greener QPCR). Water was used to adjust the volume to 25 µL. The iCycler run protocol was performed as follows: denaturation at 95°C for 5 min, 40 cycles of denaturation at 95°C for 30 s, annealing at 54°C for 30 s, and amplification at 72°C for 15 s. The specificity of the PCR amplification was examined using a melting curve program (55°C to 100°C at a heating rate of 0.5°C/s). 18S rRNA was used as the control sample with primers 18SrRNAF (5′-TCAACTTTCGATGGTAGGATAGTG-3′) and 18SrRNAR (5′-CCGTGTCAGGATTGGGTAATTT-3′). 18S rRNA expression was measured and determined to be constant at all conditions. The gene-specific primers listed in Supplementary data Table 1 were used to evaluate the quantity of target cDNA. The amplification rate of each transcript (Ct) was calculated via the PCR baseline-subtracted method and performed in the iCycler software at a constant fluorescence level. Cts were determined over three repeats. Relative fold differences were calculated based on the relative quantification analytical method (2−ΔΔCT) by using 18S rRNA amplification as the internal standard [47].
Supplementary Material
The standard curve of the biomass concentration and Nile red fluorescence value, and the gene primers for Real Time PCR.
Acknowledgments
This study was supported by the National Natural Science Foundation of China (31160050, 31360051, and 31000117), Major Technology Project of Hainan (ZDZX2013023-1), National Nonprofit Institute Research Grants (CATAS-ITBB110507, 130305), Funds of Hainan Engineering and Technological Research (GCZX2011006, GCZX2012004, and GCZX2013004), and Natural Science Foundation of Hainan Provence (313077).
Ethical Approval
This paper does not contain any studies with human or animal subjects performed by other authors.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
Authors' Contribution
Xiaodong Deng performed the molecular biology studies, participated in data analysis, and drafted the paper. Ping Li performed the detection of the mRNA levels of the genes and documented all microscopic images. Xinzhao Fan performed the lipid and TAG content detection. Xiaowen Fei performed the statistical analysis, conceived the study, participated in designing the study, coordinated the research, and assisted in drafting the paper.
References
- 1.Russell W. K., Stuber C. Effects of photoperiod and temperatures on the duration of vegetative in maize. Agro Journal. 1983;75:795–802. [Google Scholar]
- 2.Imaizumi T., Schultz T. F., Harmon F. G., Ho L. A., Kay S. A. Plant science: FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis. Science. 2005;309(5732):293–297. doi: 10.1126/science.1110586. [DOI] [PubMed] [Google Scholar]
- 3.Kobayashi Y., Weigel D. Move on up, it's time for change—mobile signals controlling photoperiod-dependent flowering. Genes and Development. 2007;21(19):2371–2384. doi: 10.1101/gad.1589007. [DOI] [PubMed] [Google Scholar]
- 4.Yano M., Katayose Y., Ashikari M., et al. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS . Plant Cell. 2000;12(12):2473–2483. doi: 10.1105/tpc.12.12.2473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nemoto Y., Kisaka M., Fuse T., Yano M., Ogihara Y. Characterization and functional analysis of three wheat genes with homology to the CONSTANS flowering time gene in transgenic rice. Plant Journal. 2003;36(1):82–93. doi: 10.1046/j.1365-313X.2003.01859.x. [DOI] [PubMed] [Google Scholar]
- 6.Drobyazina P. E., Khavkin E. E. A structural homolog of CONSTANS in potato. Russian Journal of Plant Physiology. 2006;53(5):698–701. doi: 10.1134/S1021443706050153. [DOI] [Google Scholar]
- 7.Drobyazina P. E., Khavkin E. E. The structure of two CONSTANS-LIKE1 genes in potato and its wild relatives. Gene. 2011;471(1-2):37–44. doi: 10.1016/j.gene.2010.10.005. [DOI] [PubMed] [Google Scholar]
- 8.Serrano G., Herrera-Palau R., Romero J. M., Serrano A., Coupland G., Valverde F. Chlamydomonas CONSTANS and the evolution of plant photoperiodic signaling. Current Biology. 2009;19(5):359–368. doi: 10.1016/j.cub.2009.01.044. [DOI] [PubMed] [Google Scholar]
- 9.Yuceer C., Harkess R. L., Land S. B., Jr., Luthe D. S. Structure and developmental regulation of CONSTANS-LIKE genes isolated from Populus deltoides . Plant Science. 2002;163(3):615–625. doi: 10.1016/S0168-9452(02)00169-3. [DOI] [Google Scholar]
- 10.Griffiths S., Dunford R. P., Coupland G., Laurie D. A. The evolution of CONSTANS-like gene families in barley, rice, and Arabidopsis . Plant Physiology. 2003;131(4):1855–1867. doi: 10.1104/pp.102.016188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ben-Naim O., Eshed R., Parnis A., et al. The CCAAT binding factor can mediate interactions between CONSTANS-like proteins and DNA. Plant Journal. 2006;46(3):462–476. doi: 10.1111/j.1365-313X.2006.02706.x. [DOI] [PubMed] [Google Scholar]
- 12.Liu J., Yu J., McIntosh L., Kende H., Zeevaart J. A. D. Isolation of a CONSTANS ortholog from Pharbitis nil and its role in flowering. Plant Physiology. 2001;125(4):1821–1830. doi: 10.1104/pp.125.4.1821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Schranz M. E., Mitchell-Olds T. Independent ancient polyploidy events in the sister families Brassicaceae and Cleomaceae. Plant Cell. 2006;18(5):1152–1165. doi: 10.1105/tpc.106.041111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Moon Y.-H., Chae S., Jung J.-Y., An G. Expressed sequence tags of radish flower buds and characterization of a CONSTANS LIKE 1 gene. Molecules and Cells. 1998;8(4):452–458. [PubMed] [Google Scholar]
- 15.Hecht V., Foucher F., Ferrandiz C., Macknight R. Conservation of Arabidopsis flowering genes in model legumes. Plant Physiology. 2005;137:1420–1434. doi: 10.1104/pp.104.057018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Martin J., Storgaard M., Andersen C. H., Nielsen K. K. Photoperiodic regulation of flowering in perennial ryegrass involving a CONSTANS-like homolog. Plant Molecular Biology. 2004;56(2):159–169. doi: 10.1007/s11103-004-2647-z. [DOI] [PubMed] [Google Scholar]
- 17.Robson F., Costa M. M. R., Hepworth S. R., et al. Functional importance of conserved domains in the flowering-time gene CONSTANS demonstrated by analysis of mutant alleles and transgenic plants. The Plant Journal. 2001;28(6):619–631. doi: 10.1046/j.1365-313x.2001.01163.x. [DOI] [PubMed] [Google Scholar]
- 18.Griffiths S., Dunford R. P., Coupland G., Laurie D. A. The evolution of CONSTANS-like gene families in barley, rice, and Arabidopsis. Plant Physiology. 2003;131(4):1855–1867. doi: 10.1104/pp.102.016188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ben-Naim O., Eshed R., Parnis A., et al. The CCAAT binding factor can mediate interactions between CONSTANS-like proteins and DNA. The Plant Journal. 2006;46(3):462–476. doi: 10.1111/j.1365-313X.2006.02706.x. [DOI] [PubMed] [Google Scholar]
- 20.Robert L. S., Robson F., Sharpe A., Lydiate D., Coupland G. Conserved structure and function of the Arabidopsis flowering time gene CONSTANS in Brassica napus . Plant Molecular Biology. 1998;37(5):763–772. doi: 10.1023/A:1006064514311. [DOI] [PubMed] [Google Scholar]
- 21.Holefors A., Opseth L., Ree Rosnes A. K., et al. Identification of PaCOL1 and PaCOL2, two CONSTANS-like genes showing decreased transcript levels preceding short day induced growth cessation in Norway spruce. Plant Physiology and Biochemistry. 2009;47(2):105–115. doi: 10.1016/j.plaphy.2008.11.003. [DOI] [PubMed] [Google Scholar]
- 22.Borden K. L. B. RING fingers and B-boxes: zinc-binding protein-protein interaction domains. Biochemistry and Cell Biology. 1998;76(2-3):351–358. doi: 10.1139/o98-021. [DOI] [PubMed] [Google Scholar]
- 23.Corbesier L., Coupland G. Photoperiodic flowering of Arabidopsis: integrating genetic and physiological approaches to characterization of the floral stimulus. Plant, Cell and Environment. 2005;28(1):54–66. doi: 10.1111/j.1365-3040.2005.01283.x. [DOI] [Google Scholar]
- 24.Strayer C., Oyama T., Schultz T. F., et al. Cloning of the Arabidopsis clock gene TOC1, an autoregulatory response regulator homolog. Science. 2000;289(5480):768–771. doi: 10.1126/science.289.5480.768. [DOI] [PubMed] [Google Scholar]
- 25.Torok M., Etkin L. D. Two B or not two B? Overview of the rapidly expanding B-box family of proteins. Differentiation. 2001;67(3):63–71. doi: 10.1046/j.1432-0436.2001.067003063.x. [DOI] [PubMed] [Google Scholar]
- 26.Khanna R., Kronmiller B., Maszle D. R., et al. The arabidopsis B-Box zinc finger family. Plant Cell. 2009;21(11):3416–3420. doi: 10.1105/tpc.109.069088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Robson F., Costa M. M. R., Hepworth S. R., et al. Functional importance of conserved domains in the flowering-time gene CONSTANS demonstrated by analysis of mutant alleles and transgenic plants. Plant Journal. 2001;28(6):619–631. doi: 10.1046/j.1365-313x.2001.01163.x. [DOI] [PubMed] [Google Scholar]
- 28.Griffiths S., Dunford R. P., Coupland G., Laurie D. A. The evolution of CONSTANS-like gene families in barley, rice, and Arabidopsis . Plant Physiology. 2003;131(4):1855–1867. doi: 10.1104/pp.102.016188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Jang S., Marchal V., Panigrahi K. C. S., et al. Arabidopsis COP1 shapes the temporal pattern of CO accumulation conferring a photoperiodic flowering response. EMBO Journal. 2008;27(8):1277–1288. doi: 10.1038/emboj.2008.68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kumagai T., Ito S., Nakamichi N., et al. The common function of a novel subfamily of B-box zinc finger proteins with reference to circadian-associated events in Arabidopsis thaliana. Bioscience, Biotechnology and Biochemistry. 2008;72(6):1539–1549. doi: 10.1271/bbb.80041. [DOI] [PubMed] [Google Scholar]
- 31.Indorf M., Cordero J., Neuhaus G., Rodríguez-Franco M. Salt tolerance (STO), a stress-related protein, has a major role in light signalling. Plant Journal. 2007;51(4):563–574. doi: 10.1111/j.1365-313X.2007.03162.x. [DOI] [PubMed] [Google Scholar]
- 32.Datta S., Hettiarachchi C., Johansson H., Holm M. Salt Tolerance Homolog2, a B-box protein in Arabidopsis that activates transcription and positively regulates light-mediated development. Plant Cell. 2007;19(10):3242–3255. doi: 10.1105/tpc.107.054791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Datta S., Johansson H., Hettiarachchi C., et al. LZF1/salt tolerance HOMOLOG3, an Arabidopsis B-box protein involved in light-dependent development and gene expression, undergoes COP1-mediated ubiquitination. Plant Cell. 2008;20(9):2324–2338. doi: 10.1105/tpc.108.061747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chang C.-S. J., Li Y.-H., Chen L.-T., et al. LZF1, a HY5-regulated transcriptional factor, functions in Arabidopsis de-etiolation. Plant Journal. 2008;54(2):205–219. doi: 10.1111/j.1365-313X.2008.03401.x. [DOI] [PubMed] [Google Scholar]
- 35.Ral J.-P., Colleoni C., Wattebled F., et al. Circadian clock regulation of starch metabolism establishes GBSSI as a major contributor to amylopectin synthesis in Chlamydomonas reinhardtii . Plant Physiology. 2006;142(1):305–317. doi: 10.1104/pp.106.081885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Harris E. H. The Chlamydomonas Source Book: A Comprehensive Guide to Biology and Laboratory Use. San Diego, Calif, USA: Academic Press; 1989. [DOI] [PubMed] [Google Scholar]
- 37.Deng X.-D., Gu B., Li Y.-J., Hu X.-W., Guo J.-C., Fei X.-W. The roles of acyl-CoA: diacylglycerol acyltransferase 2 genes in the biosynthesis of triacylglycerols by the green algae Chlamydomonas reinhardtii . Molecular Plant. 2012;5(4):945–947. doi: 10.1093/mp/sss040. [DOI] [PubMed] [Google Scholar]
- 38.Deng X., Li Y., Fei X. The mRNA abundance of pepc2 gene is negatively correlated with oil content in Chlamydomonas reinhardtii . Biomass and Bioenergy. 2011;35(5):1811–1817. doi: 10.1016/j.biombioe.2011.01.005. [DOI] [Google Scholar]
- 39.Liu B., Vieler A., Li C., Daniel Jones A., Benning C. Triacylglycerol profiling of microalgae Chlamydomonas reinhardtii and Nannochloropsis oceanica. Bioresource Technology. 2013;146:310–316. doi: 10.1016/j.biortech.2013.07.088. [DOI] [PubMed] [Google Scholar]
- 40.La Russa M., Bogen C., Uhmeyer A., et al. Functional analysis of three type-2 DGAT homologue genes for triacylglycerol production in the green microalga Chlamydomonas reinhardtii . Journal of Biotechnology. 2012;162(1):13–20. doi: 10.1016/j.jbiotec.2012.04.006. [DOI] [PubMed] [Google Scholar]
- 41.Gao C., Xiong W., Zhang Y., Yuan W., Wu Q. Rapid quantitation of lipid in microalgae by time-domain nuclear magnetic resonance. Journal of Microbiological Methods. 2008;75(3):437–440. doi: 10.1016/j.mimet.2008.07.019. [DOI] [PubMed] [Google Scholar]
- 42.Huang G. H., Chen G., Chen F. Rapid screening method for lipid production in alga based on Nile red fluorescence. Biomass and Bioenergy. 2009;33(10):1386–1392. doi: 10.1016/j.biombioe.2009.05.022. [DOI] [Google Scholar]
- 43.Chen W., Zhang C., Song L., Sommerfeld M., Hu Q. A high throughput Nile red method for quantitative measurement of neutral lipids in microalgae. Journal of Microbiological Methods. 2009;77(1):41–47. doi: 10.1016/j.mimet.2009.01.001. [DOI] [PubMed] [Google Scholar]
- 44.Li Y., Fei X., Deng X. Novel molecular insights into nitrogen starvation-induced triacylglycerols accumulation revealed by differential gene expression analysis in green algae Micractinium pusillum . Biomass and Bioenergy. 2012;42:199–211. doi: 10.1016/j.biombioe.2012.03.010. [DOI] [Google Scholar]
- 45.Kindle K. L. High-frequency nuclear transformation of Chlamydomonas reinhardtii . Proceedings of the National Academy of Sciences of the United States of America. 1990;87(3):1228–1232. doi: 10.1073/pnas.87.3.1228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Fei X. W., Deng X. D. A novel Fe deficiency-responsive element (FeRE) regulates the expression of atx1 in Chlamydomonas reinharditii . Plant and Cell Physiology. 2007;48(10):1496–1503. doi: 10.1093/pcp/pcm110. [DOI] [PubMed] [Google Scholar]
- 47.Livak K. J., Schmittgen T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods. 2001;25(4):402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The standard curve of the biomass concentration and Nile red fluorescence value, and the gene primers for Real Time PCR.