

Abstract
Mycobacterium tuberculosis (Mtb) infects about one third of the world’s population, with a majority of infected individuals exhibiting latent asymptomatic infection, while 5–10% of infected individuals progress to active pulmonary disease. Research in the past two decades has elucidated critical host immune mechanisms that mediate Mtb control. Among these, chemokines have been associated with numerous key processes that lead to Mtb containment, from recruitment of myeloid cells into the lung to activation of adaptive immunity, formation of protective granulomas and vaccine recall responses. However, imbalances in several key chemokine mediators can alter the delicate balance of cytokines and cellular responses that promote mycobacterial containment, instead precipitating terminal tissue destruction and spread of Mtb infection. In this review, we will describe recent insights in the involvement of chemokines in host responses to Mtb infection and Mtb containment (the good), chemokines contributing to inflammation during TB (the bad), and the role of chemokines in driving cavitation and lung pathology (the ugly).
Keywords: Chemokines, Mycobacterial infections, lung
1. Introduction
Tuberculosis (TB), caused by infection with Mycobacterium tuberculosis (Mtb), is estimated to affect one third of the world’s population. The majority of infected individuals develop asymptomatic latent TB, while ~5–10% of latently infected individuals will progress to active pulmonary TB (ATB), resulting in about 9 million new cases of TB and 1.4 million deaths per year [1]. The long drug treatment regimes, the relative inefficacy of the current TB vaccine, in addition to the increase in drug-resistant TB cases [1], stresses the importance of understanding host immune responses that mediate Mtb control. The past two decades have broadened our understanding of the immune mechanisms required for Mtb containment and delineated that the key processes regulating TB control or disease exacerbation involve the recruitment of host immune cell populations into the lung. This process is governed by adhesion molecules and by chemoattractant cytokines or “chemokines”, a family of small proteins, which, upon binding to membrane G protein-coupled receptors, guide the gradient-driven migration of leukocytes [2]. Chemokines are classified into the CXC-, CC-, C- and CX3C- subfamilies according to the arrangement of four conserved cysteine residues, which are important for maintenance of their tridimensional structure [2]. A recent review has described the general structure of chemokines and their overall functions in TB [3]. In this review, we have specifically focused on chemokines and their effector mechanisms that contribute to pulmonary control of Mtb infection. In addition, we will discuss the importance of chemokines in the establishment of a balance between proinflammatory and anti-inflammatory mediators during TB that may result in improved Mtb control or exacerbated disease outcomes.
2. Role of chemokines in mediating Mtb control (The good)
Over the past two decades, the availability of animal models of TB, in addition to human studies, have shed light on several key chemokine-driven immune mechanisms mediating Mtb control [4]. Mtb reaches the lower airways of the lung via inhalation of 3–5 μm droplet nuclei, generated during coughing or sneezing. Upon entry into the lung, mycobacteria are taken up by alveolar macrophages, where Mtb replicates while inhibiting macrophage killing mechanisms [5]. Despite this, infected macrophages actively secrete chemokines and cytokines, resulting in the recruitment and activation of several immune cell populations to the lung [5]. Indeed, in the mouse model of low dose aerosol infection, around day 12 post-infection there is an early influx of innate cells into the lungs, including γδ T cells, NK cells, monocyte-derived macrophages, dendritic cells and neutrophils [6]. It is possible that distinct chemokines govern the specific recruitment of these diverse immune cells to the lung. In particular, increased expression of the chemokines CXCL-3 and CXCL-5 is observed as early as day 12 after infection [6], and this correlates with the early influx of neutrophils and NK cells, which likely express the receptor CXCR2. In addition, lung epithelial cells can directly sense Mtb and produce chemokines, resulting in a potentiation of immune cell recruitment. In response to Mtb stimulation, CCL-2 and CXCL-8 are produced by a line of alveolar epithelial cells and by human bronchial epithelial cells [7, 8]. In addition, in the mouse model of Mtb infection, following TLR-2 ligation, the lung epithelium has been described to secrete CXCL-5, which, signaling through CXCR2, can increase neutrophil recruitment [9]. Despite the accumulation of these innate immune cells, Mtb continues to grow exponentially over the first 2–3 weeks following infection [6]. Thus, activation of adaptive immunity and recruitment of effector T cells into the lung is required for bacterial burden control [10]. The priming of T cells is initiated by dendritic cells (DCs), primary antigen presenting cells (APCs) that serve as a direct link between the innate branch of the immune response and the adaptive response [11].
Lung resident DCs can take up live Mtb within the lungs and transport them to the lung-draining mediastinal lymph nodes, where they were thought to serve as APCs [12]. Migration of DCs from the lungs to the mediastinal lymph nodes is governed by chemokine-receptor interactions, and occurs around day 14 post-infection in the mouse model of TB [12]. Uptake of Mtb by DCs leads to the upregulation of CCR7 expression [13], which guides the cells to the mediastinal lymph node following a gradient of the homeostatic chemokines CCL-19 and CCL-21 [12]. CCL-21 is expressed by the lymphatic endothelium, directing the initial migration of DCs, while CCL-19 and CCL-21 are expressed by lymph node resident cells. Importantly, mice lacking CCR7 have an impaired ability to migrate to the draining lymph nodes, resulting in delayed priming of Mtb-specific T cells [14]. Recently, it has come to light that the cell populations that become infected and carry antigen to the lymph node, and those that directly prime the T cells, are distinct. Indeed, infected CCR2+ inflammatory monocytes are important for antigen delivery into the lung, where they release soluble antigen that can be taken up and presented by resident lymph node DCs [15, 16]. Subsequent recognition of Mtb antigens by naïve T cells bearing specific T cell receptors, in the presence of costimulatory signals and adequate cytokines in the microenvironment leads to the activation, proliferation and differentiation of naïve T cells into effector cells [17].
While Mtb actively replicates in the lung, induction of inflammatory chemokines ultimately results in the recruitment of newly activated effector T cells from the periphery. T cells that exit the lymph node are able to enter the lung via the circulation through ligation of surface endothelial receptors that are upregulated in response to inflammation. Several chemokines and their cognate receptors have been associated with T cell migration into the lung during TB. CD4+ and CD8+ T cell activation and differentiation in the lymph node is accompanied by changes in surface chemokine receptor expression and the corresponding alteration of their migratory capacity. Upon commitment to the Th1 subset, the main CD4+ T cell subset implicated in Mtb control, effector T cells upregulate the chemokine receptors CXCR3 and CCR5 [18, 19]. It is thought that this is directly related to their recruitment into the infected lung, as the ligands for these receptors, CXCL-9, CXCL-10 and CXCL-11 for CXCR3 and CCL-3, CCL-4, CCL-5 and CCL-8 for CCR5, are upregulated in Mtb-infected mouse [6] and NHP lungs [20]. Several mechanistic studies have addressed the requirement for CXCR3 and CCR5 expression on T cells [21, 22], providing evidence that there is significant redundancy in the expression of these inflammatory chemokines and their receptors on the recruitment of Mtb-specific T cells to the lung. Human studies have shown associations between mutations in CCL-2 and CCL-5 and pulmonary TB [23–25], suggesting that despite the redundancy observed in animal models, these chemokines may have defined roles to play in human TB.
Upon entry into the lung parenchyma, however, proper Mtb containment is dependent on the correct localization of effector T cells in apposition to Mtb-infected macrophages. In recent years, several reports have demonstrated the expression of homeostatic chemokines, which are commonly expressed in secondary lymphoid organs (SLOs), in Mtb-infected lungs [6, 26]. Such chemokines, including CCL-19, CCL-21, CXCL-12 and CXCL-13, drive the organization of lymphoid follicles in SLOs and in the periphery [26]. These organized lymphoid and stromal aggregates, known as ectopic lymphoid follicles, have been reported in conditions of chronic infection and inflammation [27]. Interestingly, during Mtb infection in mice, non-human primates and humans, CXCR5-expressing CD4+ T cells also accumulate in the lungs, within ectopic lymphoid follicles [28]. Importantly, these CXCR5+ CD4+ T cells produce high levels of proinflammatory cytokines and upon accumulation in the lung, respond to CXCL-13 likely produced by stromal cells early during infection, and localize near Mtb-infected macrophages to mediate Mtb control [28]. Accordingly, both CXCR5 and CXCL13-deficient mice lacked the formation of ectopic lymphoid follicles and exhibited decreased control of Mtb, thus projecting the non-redundant role for CXCR5-CXCL-13 axis in TB. CXCR5 deficiency resulted in localization of CD4+ T cells around blood vessels in the Mtb-infected lungs, forming perivascular cuffs indicative of their inability to localize in apposition to infected macrophages [28]. Therefore, not only is the timely induction of chemokine-mediated recruitment of T cells to the lung critical for Mtb control, but emerging evidence suggests that chemokines also play a critical role in the precise positioning of Mtb-specific T cells within the lung parenchyma for maximal Mtb control. Indeed, early vaccine-induced production of CXCL-9, CXCL-10 and associated recruitment of CXCR3-expressing T cells is beneficial in vaccine-induced protection against Mtb challenge [29]. In addition, vaccine strategies that induce early CXCL-13 production to enhance and improve early T cell localization near Mtb-infected macrophages can be harnessed for vaccine design against TB [30].
Together, there is accumulating evidence that chemokines induced in response to Mtb infection effectively mediate DC trafficking to the LNs, recruitment of activated T cells to the lung and correct localization of T cells within the lung parenchyma to mediate optimal Mtb control. However, although these chemokine-dependent processes mediate control of Mtb growth, they often do not completely eliminate the bacteria (Figure 1). Further understanding of the mechanisms that lead to Mtb containment will not only allow the better development of novel therapies against TB, but will be of particular relevance for vaccine and adjuvant design.
Figure 1. “The good”: mechanisms that mediate lung Mtb containment.
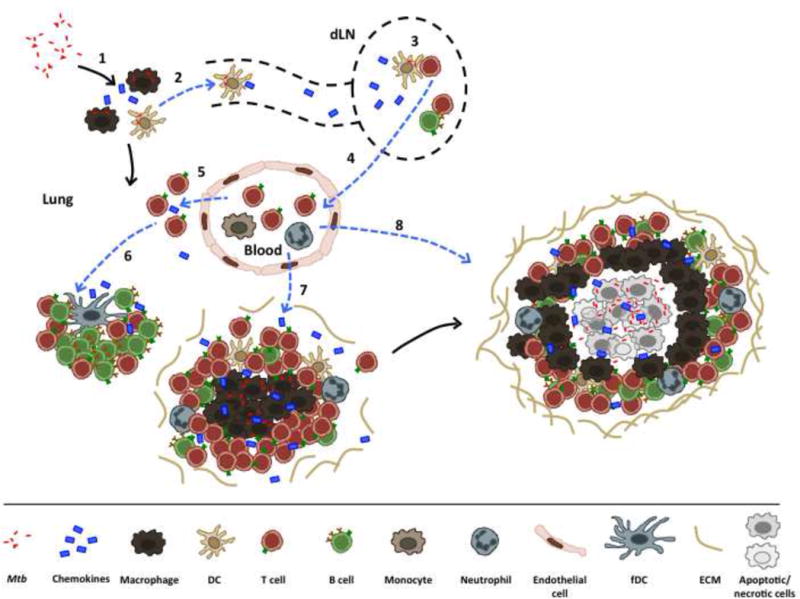
Upon Mtb entry into the lungs, alveolar macrophages become infected, leading to secretion of cytokines and chemokines, which drive additional innate cell recruitment (1). Infected DCs migrate into the lung draining lymph nodes (dLN) (2), carrying antigen that can be subsequently taken up by other APCs to activate naïve T cells (3). After activation, T cells (along with B cells) regulate their chemokine receptor expression, which guide their exit from the lymph node (4), homing to the infected lung (5), and subsequent migration. These responses are mediated by differential expression of chemokines, a process that enables ectopic lymphoid follicle formation (6). Additional innate cells, such as monocytes and neutrophils are also recruited into the lung (7). Together, interactions between innate and adaptive cells lead to granuloma formation and Mtb containment (8). Dashed blue lines represent chemokine-driven mechanisms.
3. Chemokines mediate inflammation during TB (The bad)
The aforementioned mechanisms of TB containment rely on a precise site and time-specific upregulation of chemokines and their receptors. However, numerous factors can shift the balance to limited containment or pathology. Indeed, the nonresolving immune activation that occurs in chronic diseases such as TB can lead to tissue damage and pathology. Given that maintenance of lung architecture is essential for adequate organ function, unrestricted inflammation at this site is associated with respiratory failure and increased mortality in TB patients [31]. Identification of the factors leading to exacerbated inflammation within TB lungs will enable the development of new therapies for TB.
A neutrophil-associated human blood transcriptional signature was seen in patients with ATB [32], and neutrophils were identified as the predominant cell infected with replicating Mtb in ATB patients [33]. In mice, different inbred strains vary in their susceptibility to Mtb. In particular, resistant strains such as C57BL/6 form smaller lung lesions mostly comprised of lymphocytes and macrophages [34]. Susceptible strains, including C3H and I/St mice, however, form more diffuse and less organized lesions, with increased neutrophil infiltration [35–37]. Emerging evidence suggests that the accumulation of neutrophils during TB is mediated by interaction between the chemokine CXCL-5 and its receptor CXCR2, specifically expressed on neutrophils. Dorhoi et al [38] recently reported that the increased susceptibility to Mtb challenge observed in miR-223-deficient mice, could be reversed by CXCL-2 blockade, CCL-3 and IL-6 neutralization, or by neutrophil depletion [38], underscoring the importance of tightly regulated inflammation for host survival. Importantly, in a high dose infection model, mice deficient in CXCR2 or CXCL-5, had a prolonged survival and decreased lung pathology in comparison to their wild type counterparts [9]. That the CXCL-5 upregulation was mediated by TLR-2 signaling [9], suggests that Mtb may have evolved ways to harness neutrophil recruitment to enhance inflammation and disease severity in the host. In unraveling the mechanisms by which neutrophils mediate lung pathology in TB, we recently showed that in NHPs and patients with ATB, increased pulmonary pathology correlated with increased neutrophil accumulation and expression of neutrophil-associated products such as S100A8/A9 proteins [39]. The effect of S100A8/A9 proteins in promoting neutrophil and monocyte migration occurs through the induction of proinflammatory chemokines such as CXCL-1, and the upregulation of integrins such as CD11b on neutrophils. Thus, in ATB patients and diversity outbred (DO) mice, which constitute a new model of the genetic variability, lung damage score correlated well with levels of S100A8/A9 and CXCL-1 protein levels [39].
Consistent with these findings, several recent papers have linked increased levels of specific neutrophil chemokines with ATB in patients, suggesting their potential as immunological markers for TB and for treatment responsiveness monitoring [40, 41]. Indeed, CCL-3, CXCL-8 and CCL-2 expression was upregulated in neutrophils isolated from patients with pulmonary TB in comparison to healthy controls, and following in vitro Mtb infection [42]. In addition, the CCL-2 2518G allele, which results in exacerbated CCL-2 secretion [43], and combinations of single-nucleotide polymorphisms in the CCL-5 promoter [24] have been associated with TB in human populations. Other chemokines and cytokines have also been shown to be elevated during pulmonary ATB. In particular, Yu et al demonstrated elevated IL-2, CXCL-10, CXCL-11 and CXCL-12 in patients with ATB and increased levels of CCL-1, CCL-21 and IL-6 in patients with tuberculous pleuritis [44]. Further, chemokine determination in the sera of ATB patients before and after treatment could serve as a correlate of treatment efficacy, as a decrease in the serum levels of CXCL-8, CXCL-9 and CXCL-10 was observed in patients with ATB following antibiotic treatment completion [45]. In addition, genetic studies have revealed an association of the 135G/A polymorphism at the level of the CXCL-10 allele and TB in a Chinese population [46]. Taken together, these findings demonstrate an emerging role for chemokines in mediating neutrophil accumulation and perpetuating inflammation during TB (Figure 2), which could be harnessed as novel therapeutic regimes and utilized as novel biomarkers.
Figure 2. “The bad”: transition to a dysregulated proinflammatory granuloma.
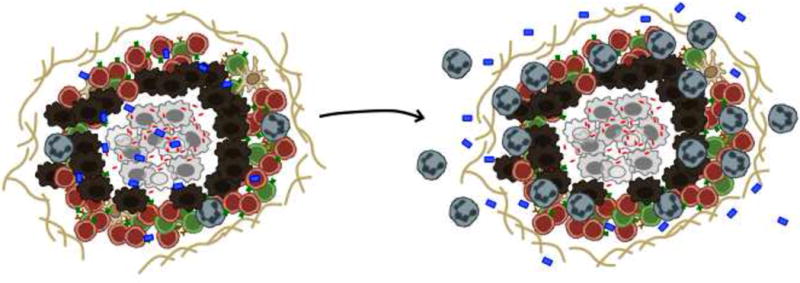
An imbalance between anti-inflammatory and pro-inflammatory factors can lead to dysregulated inflammation in TB, a feature of which is accumulation of large numbers of neutrophils in the lungs. Through the secretion of numerous chemokines and molecules, such as S100A8/A9 proteins, neutrophils can perpetuate inflammation during TB.
4. Role of chemokines in cavitary TB (The ugly)
A hallmark of active TB is the development of pulmonary cavities, which are thought to harbor high levels of replicating bacteria and constitute a contributing source of disease transmission [47]. In humans, cavitation usually occurs in the lung apices and requires dissemination of bacteria from the lung bases, where the infection typically originates [48]. The current paradigm is that cavitary TB is a product of ineffective granuloma formation, with necrosis development, liquefaction of the necrotic areas and subsequent connection of the mycobacteria-rich granulomatous content and the airways [47]. Caseation is an active process led forward by host and bacterial factors, and potentiated by unrestricted inflammation. Indeed, immune mediators such as reactive oxygen species and reactive nitrogen intermediates [49, 50], cytokines and pro-apoptotic receptors [51, 52] can induce macrophage necrosis and/or apoptosis, releasing the cell contents that form the center of the caseous granuloma. Given that lung structure is maintained by tightly bound collagen fibrils, active processes involving hydrolase activation are required for destruction of the lung parenchyma and subsequent cavitation. Amongst the hydrolases that participate in tissue destruction, matrix metalloproteinases (MMPs) play a central role in cavitation [48].
MMPs are a family of zinc-containing proteases with a range of substrate specificities and sources that play a key role in extracellular matrix degradation. In addition, MMPs can modulate cytokine and chemokine activity by cleavage, either inactivating them or potentiating their biological activity. Importantly, MMP activity is implicated in TB progression both in animal models and in humans with ATB. Indeed, in a zebrafish model of M. marinum infection, epithelial cells upregulated MMP-9 expression in response to Mtb-derived ESAT-6, and enhanced macrophage recruitment, increasing granuloma growth and mycobacterial proliferation [53]. In addition, mice overexpressing the human MMP-1 gene under the control of the macrophage-specific promoter for scavenger receptor A had increased lung matrix destruction during Mtb infection [54]. Further, use of the broad spectrum MMP inhibitor BB-94 decreased Mtb burden, reduced granuloma size and leukocyte recruitment [55, 56]. MMP-9-deficient mice had lower lung macrophage recruitment with less well-formed granulomas and reduced bacteria [57]. In animal models where cavitation occurs in response to TB, such as the guinea pig [58], and rabbit [59], activation of proteolytic enzymes is associated with lung damage and cavitation [60, 61]. Recently, elevated sputum levels of several MMPs were reported in patients with TB, and MMP-1 and -3 concentrations positively correlated with TB severity [62]. Interestingly, MMP levels at the time of diagnosis negatively correlated with responsiveness to treatment [62].
In addition to driving direct tissue destruction, MMPs can exert their functions on chemokines, thereby altering leukocyte recruitment. Chemokine digestion by MMPs can have several outcomes, ranging from functional inactivation, to the generation of antagonistic variants and to potentiation of chemotactic activity [63]. Inactivation of several chemokines has been shown to occur through MMP-mediated cleavage. In fact, MMP-1, -2, -3, -9, -13 and -14 can cleave CXCL-12, rendering it functionally inactive [64]. Given that CXCL-12 is one of the homeostatic chemokines induced in the mouse lung in response to Mtb infection [6], MMP digestion could potentially affect ectopic lymphoid follicle formation, thereby altering Mtb control. MMP-9 can also inactivate CXCL-4 and CXCL-1 [65], potentially affecting monocyte and neutrophil recruitment. In addition, MMP-8 and MMP-9 can proteolytically inactivate CXCL-9 and CXCL-10, but not CXCL-11 [66], a process that could affect T cell recruitment into Mtb-infected lungs. However, given the redundancy between CXCL9, -10 and -11, whether this mechanism substantially affects T cell migration in TB is not known. Another potential outcome of MMP-driven chemokine proteolysis is the generation of variants that retain receptor binding ability but are unable to mediate chemotaxis. Because they can compete with active chemokines, these variants could serve as chemokine antagonists. For instance, MMP-1 and MMP-3 can cleave CCL-2, generating an antagonist molecule [67] that may affect T cell, neutrophil and NK cell recruitment via CXCR2 during Mtb infection. MMP-3, in addition, can generate a CCL-8 antagonist [67], which could impact CCR5+ cell recruitment. In addition, MMPs can potentiate chemokine activity. For instance, MMP-9 can process CXCL-8 (IL-8), leading to an increase in chemotactic activity [65]. CXCL-5, which signals through CXCR2, can also be processed by MMP-9 into a more active molecule [68]. Given that neutrophils are a main source of MMP-9, this could serve as a positive feedback loop, allowing the perpetuation of an inflammatory response in TB. In vivo, chemokines bind to glycosaminoglycans, which contribute to their stability and to the adequate formation of tridimensional gradients. MMP activity on extracellular matrix proteins can therefore also indirectly affect chemokine activity. For instance, the chemokine CXCL-1 can bind the cell surface-expressed proteoglycan syndecan-1. Interestingly, MMP-7 and MMP-9 can cleave the extracellular domain of syndecan-1, releasing the syndecan-1/CXCL-1 complex [69]. This creates a chemokine gradient that promotes neutrophil infiltration into the alveolar space and can drive further inflammation during acute lung injury [69]. Resonating with these findings, recent reports have shown increased neutrophils in bronchoalveolar lavages from patients with cavitary TB in comparison to non-cavitary TB patients, associated with decreased CXCL-10 and IL-6, which may indicate the failure of adaptive immunity at this stage of disease [70].
The events leading to terminal tissue destruction and cavitation in TB are beginning to be defined, and likely involve a complex interplay between immune activation, protease function and bacterial factors. Given the dual effect of MMPs in chemokine activity, it is likely that an imbalance between chemokine inactivation and potentiation leads to the unrestricted recruitment of immune cells to the site of infection. This, in turn, may promote overwhelming lung inflammation and destruction, contributing to the events ultimately leading to cavity formation and TB spread (Figure 3). Future studies will shed light on the relative contribution of MMP-derived chemokine variants in TB pathology. Of note, mouse models where granuloma necrosis occurs, such as the C3HeB/FeJ strain [71], will be of particular utility for the study of the mechanistic relevance of MMPs and chemokine variants to lung damage.
Figure 3. “The ugly”: TB necrosis, cavitation and spread.
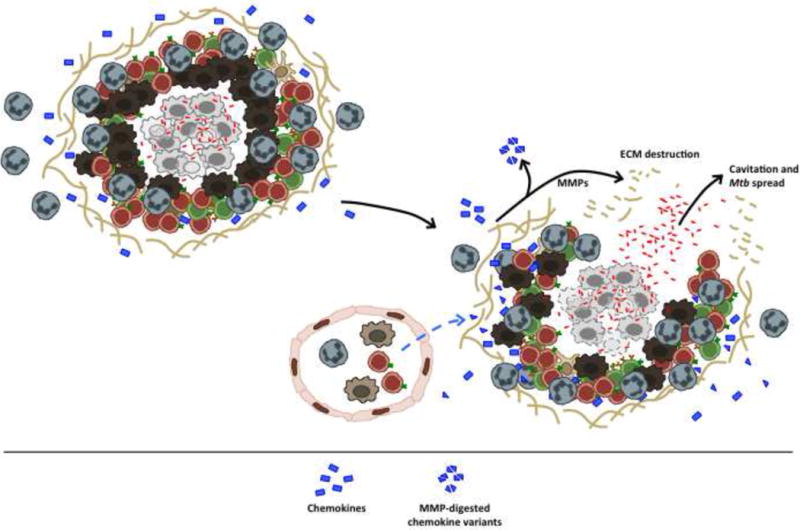
The hallmark of active TB in humans and in several animal models is cavitation, which is a product of persistent inflammation and tissue destruction. Central to this process are matrix metalloproteinases, which can degrade the extracellular matrix that maintains lung structure, and cleave chemokines. These novel chemokine variants can be more or less active than their native counterparts, and their production likely alters the balance of cells that are recruited into the lung. This overwhelming inflammation and matrix destruction communicates Mtb-rich granulomas with the airways, facilitating Mtb spread.
6. Conclusions
Chemokines play a central role in orchestrating the recruitment of cells into the Mtb-infected lung, which contributes to Mtb containment. However, under specific conditions, chemokines can also drive the disproportionate inflammation and lung damage that precipitate progression to pulmonary disease as well as cavitation. In humans, the balance between protective and damaging inflammation, as well as the levels of inflammation required for Mtb containment may differ. A number of factors, including host genotype, bacterial strain, co-morbidities and nutritional status likely shift the magnitude and nature of mechanisms that permit Mtb control. Understanding the nature and the effect of these interactions between the host, mycobacteria and the environment will be central to the design of effective therapeutic and vaccination strategies.
Table 1.
Chemokines and chemokine receptors in TB
| Receptor | Ligands | Role in TB | Reference |
|---|---|---|---|
| CCR2 | CCL2,7,8,12,13,16 | Recruitment of monocytes, DCs, T cells and NK cells. | [72–74] |
| CCR4 | CCL17,22 | Recruitment of T cells. May recruit NK cells, macrophages, DCs and basophils. | [75, 76] |
| CCR5 | CCL3,4,5,8 | Recruitment of T cells into the lung. May be involved in macrophage, DC and neutrophil migration. | [19, 75, 77] |
| CCR6 | CCL20 | iNKT and T cell recruitment. | [78, 79] |
| CCR7 | CCL19,21 | DC migration into the draining lymph nodes, recruitment of naïve and central memory cells to lymph nodes, ectopic lymphoid follicle formation. | [12–14, 26, 80] |
| CXCR1 | CXCL6,8 | Recruitment of neutrophils. | [81] |
| CXCR2 | CXCL1,2,3,5,6,7,8 | Recruitment of neutrophils and NK cells. | [73, 81, 82] |
| CXCR3 | CXCL9,10,11 | Recruitment of T cells into the lung. | [6, 18–20] |
| CXCR5 | CXCL13 | Ectopic lymphoid follicle formation. | [28] |
Highlights.
Chemokines govern cell influx to Mtb-infected lungs and are critical for TB control.
Productive granuloma formation is tightly regulated by lung-secreted chemokines.
Chemokine dysregulation can shift the balance from protection to inflammation.
Host, pathogen and environmental factors can impact chemokine secretion in the TB lung.
Acknowledgments
This work was supported by Washington University in St. Louis and NIH grant HL105427 to S.A.K., and a Children’s Hospital of Pittsburgh Research Advisory Committee Grant from Children’s Hospital of Pittsburgh of the UPMC Health System to L.M.
Abbreviations
- APC
Antigen presenting cell
- ATB
Active tuberculosis
- CD
Cluster of differentation
- CXCL
Chemokine (C-X-C motif) ligand
- CXCR
C-X-C chemokine receptor
- DC
Dendritic cell
- ESAT-6
Early Secretory Antigenic Target 6 kDa
- iNKT
invariant Natural Killer T cell
- IL
Interleukin
- LN
Lymph node
- miR
Micro RNA
- MMP
Matrix metalloproteinase
- Mtb
Mycobacterium tuberculosis
- NHP
Non-human primate
- NK cells
Natural Killer cells
- SLO
Secondary lymphoid organ
- TB
Tuberculosis
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Dye C, Glaziou P, Floyd K, Raviglione M. Prospects for tuberculosis elimination. Annual review of public health. 2013;34:271–286. doi: 10.1146/annurev-publhealth-031912-114431. [DOI] [PubMed] [Google Scholar]
- 2.Zlotnik A, Yoshie O. Chemokines : A New Classification System and Their Role in Immunity. Immunity. 2000;12:121–127. doi: 10.1016/s1074-7613(00)80165-x. [DOI] [PubMed] [Google Scholar]
- 3.Slight SR, Khader SA. Chemokines shape the immune responses to tuberculosis. Cytokine & growth factor reviews. 2013;24:105–113. doi: 10.1016/j.cytogfr.2012.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Flynn JL. Lessons from experimental Mycobacterium tuberculosis infections. Microbes Infect. 2006;8:1179–1188. doi: 10.1016/j.micinf.2005.10.033. [DOI] [PubMed] [Google Scholar]
- 5.Guirado E, Schlesinger LS, Kaplan G. Macrophages in tuberculosis: friend or foe. Seminars in immunopathology. 2013;35:563–583. doi: 10.1007/s00281-013-0388-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kang DD, Lin Y, Rangel-Moreno J, Randall TD, Khader SA. Profiling early lung immune responses in the mouse model of tuberculosis. PLoS One. 2011;6:e16161–e16161. doi: 10.1371/journal.pone.0016161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lin Y, Zhang M, Barnes PF. Chemokine production by a human alveolar epithelial cell line in response to Mycobacterium tuberculosis. Infection and immunity. 1998;66:1121–1126. doi: 10.1128/iai.66.3.1121-1126.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wickremasinghe MI, Thomas LH, Friedland JS. Pulmonary epithelial cells are a source of IL-8 in the response to Mycobacterium tuberculosis: essential role of IL-1 from infected monocytes in a NF-kappa B-dependent network. Journal of immunology. 1999;163:3936–3947. [PubMed] [Google Scholar]
- 9.Nouailles G, Dorhoi A, Koch M, Zerrahn J, Weiner J, 3rd, Fae KC, et al. CXCL5-secreting pulmonary epithelial cells drive destructive neutrophilic inflammation in tuberculosis. J Clin Invest. 2014;124:1268–1282. doi: 10.1172/JCI72030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cooper AM, Khader SA. The role of cytokines in the initiation, expansion, and control of cellular immunity to tuberculosis. Immunological reviews. 2008;226:191–204. doi: 10.1111/j.1600-065X.2008.00702.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tian T, Woodworth J, Sköld M, Behar M. In vivo depletion of CD11c+ cells delays the CD4+ T cell response to Mycobacterium tuberculosis and exacerbates the outcome of infection. The Journal of Immunology. 2005;175:3268–3272. doi: 10.4049/jimmunol.175.5.3268. [DOI] [PubMed] [Google Scholar]
- 12.Wolf AJ, Linas B, Trevejo-Nuñez GJ, Kincaid E, Tamura T, Takatsu K, et al. Mycobacterium tuberculosis Infects Dendritic Cells with High Frequency and Impairs Their Function In Vivo. J Exp Med. 2007;179 doi: 10.4049/jimmunol.179.4.2509. [DOI] [PubMed] [Google Scholar]
- 13.Bhatt K, Hickman SP, Salgame P. Cutting Edge: A New Approach to Modeling Early Lung Immunity in Murine Tuberculosis. The Journal of Immunology. 2004;172:2748–2751. doi: 10.4049/jimmunol.172.5.2748. [DOI] [PubMed] [Google Scholar]
- 14.Olmos S, Stukes S, Ernst JD. Ectopic Activation of Mycobacterium tuberculosis -Specific CD4+ T Cells in Lungs of CCR7−/− Mice. The Journal of Immunology. 2010;184:895–901. doi: 10.4049/jimmunol.0901230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Srivastava S, Ernst JD. Cell-to-cell transfer of M. tuberculosis antigens optimizes CD4 T cell priming. Cell Host Microbe. 2014;15:741–752. doi: 10.1016/j.chom.2014.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Samstein M, Schreiber HA, Leiner IM, Susac B, Glickman MS, Pamer EG. Essential yet limited role for CCR2+ inflammatory monocytes during Mycobacterium tuberculosis-specific T cell priming. eLife. 2013;2:e01086. doi: 10.7554/eLife.01086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Reiley WW, Calayag MD, Wittmer ST, Huntington JL, Pearl JE, Fountain JJ, et al. ESAT-6-specific CD4 T cell responses to aerosol Mycobacterium tuberculosis infection are initiated in the mediastinal lymph nodes. Proc Natl Acad Sci U S A. 2008;105:10961–10966. doi: 10.1073/pnas.0801496105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Groom JR, Luster AD. CXCR3 ligands: redundant, collaborative and antagonistic functions. Immunol Cell Biol. 2011;89:207–215. doi: 10.1038/icb.2010.158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Campbell JD, HayGlass KT. T cell chemokine receptor expression in human Th1- and Th2- associated diseases. Archivum immunologiae et therapiae experimentalis. 2000;48:451–456. [PubMed] [Google Scholar]
- 20.Mehra S, Pahar B, Dutta NK, Conerly CN, Philippi-Falkenstein K, Alvarez X, et al. Transcriptional reprogramming in nonhuman primate (rhesus macaque) tuberculosis granulomas. PLoS One. 2010;5:e12266. doi: 10.1371/journal.pone.0012266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Seiler P, Aichele P, Bandermann S, Hauser AE, Lu B, Gerard NP, et al. Early granuloma formation after aerosol Mycobacterium tuberculosis infection is regulated by neutrophils via CXCR3-signaling chemokines. Eur J Immunol. 2003;33:2676–2686. doi: 10.1002/eji.200323956. [DOI] [PubMed] [Google Scholar]
- 22.Chakravarty SD, Xu J, Lu B, Gerard C, Flynn J, Chan J. The chemokine receptor CXCR3 attenuates the control of chronic Mycobacterium tuberculosis infection in BALB/c mice. J Immunol. 2007;178:1723–1735. doi: 10.4049/jimmunol.178.3.1723. [DOI] [PubMed] [Google Scholar]
- 23.Feng WX, Flores-Villanueva PO, Mokrousov I, Wu XR, Xiao J, Jiao WW, et al. CCL2-2518 (A/G) polymorphisms and tuberculosis susceptibility: a meta-analysis. Int J Tuberc Lung Dis. 2012;16:150–156. doi: 10.5588/ijtld.11.0205. [DOI] [PubMed] [Google Scholar]
- 24.Chu SF, Tam CM, Wong HS, Kam KM, Lau YL, Chiang AK. Association between RANTES functional polymorphisms and tuberculosis in Hong Kong Chinese. Genes and immunity. 2007;8:475–479. doi: 10.1038/sj.gene.6364412. [DOI] [PubMed] [Google Scholar]
- 25.Sanchez-Castanon M, Baquero IC, Sanchez-Velasco P, Farinas MC, Ausin F, Leyva-Cobian F, et al. Polymorphisms in CCL5 promoter are associated with pulmonary tuberculosis in northern Spain. Int J Tuberc Lung Dis. 2009;13:480–485. [PubMed] [Google Scholar]
- 26.Khader SA, Rangel-moreno J, Jeffrey J, Martino CA, Reiley WW, John E, et al. In a Murine Tuberculosis Model, the Absence of Homeostatic Chemokines Delays Granuloma Formation and Protective Immunity. The Journal of Immunology. 2009;183 doi: 10.4049/jimmunol.0901937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pitzalis C, Jones GW, Bombardieri M, Jones SA. Ectopic lymphoid-like structures in infection, cancer and autoimmunity. Nat Rev Immunol. 2014;14:447–462. doi: 10.1038/nri3700. [DOI] [PubMed] [Google Scholar]
- 28.Slight SR, Rangel-Moreno J, Gopal R, Lin Y, Fallert Junecko BA, Mehra S, et al. CXCR5 + T helper cells mediate protective immunity against tuberculosis. J Clin Invest. 2013;123:712–726. doi: 10.1172/JCI65728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Khader SA, Bell GK, Pearl JE, Fountain JJ, Rangel-moreno J, Cilley GE, et al. IL-23 and IL-17 in the establishment of protective pulmonary CD4 + T cell responses after vaccination and during Mycobacterium tuberculosis challenge. Nat Immunol. 2007;8:369–377. doi: 10.1038/ni1449. [DOI] [PubMed] [Google Scholar]
- 30.Gopal R, Rangel-Moreno J, Slight S, Lin Y, Nawar HF, Fallert Junecko BA, et al. Interleukin-17-dependent CXCL13 mediates mucosal vaccine-induced immunity against tuberculosis. Mucosal Immunol. 2013;6:972–984. doi: 10.1038/mi.2012.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sacks LV, Pendel S. Factors related to in-hospital deaths in patients with tuberculosis. Arch Intern Med. 1998;158:1916–1922. doi: 10.1001/archinte.158.17.1916. [DOI] [PubMed] [Google Scholar]
- 32.Berry MPR, Graham CM, McNab FW, Xu Z, Bloch SAA, Oni T, et al. An interferon-inducible neutrophil-driven blood transcriptional signature in human tuberculosis. Nature. 2010;466:973–977. doi: 10.1038/nature09247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Eum S-y, Kong J-h, Hong M-s, Lee Y-j. Neutrophils Are the Predominant Infected Phagocytic Cells in the Airways of Patients With Active Pulmonary TB. Chest. 2010;137:122–128. doi: 10.1378/chest.09-0903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Aly S, Wagner K, Keller C, Malm S, Malzan A, Brandau S, et al. Oxygen status of lung granulomas in Mycobacterium tuberculosis-infected mice. J Pathol. 2006;210:298–305. doi: 10.1002/path.2055. [DOI] [PubMed] [Google Scholar]
- 35.Keller C, Hoffmann R, Lang R, Brandau S, Hermann C, Ehlers S. Genetically Determined Susceptibility to Tuberculosis in Mice Causally Involves Accelerated and Enhanced Recruitment of Granulocytes. Infect Immun. 2006;74:4295–4309. doi: 10.1128/IAI.00057-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kondratieva E, Logunova N, Majorov K, Averbakh M, Apt A. Host Genetics in Granuloma Formation : Human-Like Lung Pathology in Mice with Reciprocal Genetic Susceptibility to M. tuberculosis and M. avium. PLoS One. 2010;5 doi: 10.1371/journal.pone.0010515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Eruslanov EB, Lyadova IV, Tatiana K, Majorov KB, Scheglov IV, Orlova MO, et al. Neutrophil Responses to Mycobacterium tuberculosis Infection in Genetically Susceptible and Resistant Mice. Infect Immun. 2005;73:1744–1753. doi: 10.1128/IAI.73.3.1744-1753.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Dorhoi A, Iannaccone M, Farinacci M, Fae KC, Schreiber J, Moura-Alves P, et al. MicroRNA-223 controls susceptibility to tuberculosis by regulating lung neutrophil recruitment. J Clin Invest. 2013;123:4836–4848. doi: 10.1172/JCI67604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gopal R, Monin L, Torres D, Slight SR, Mehra S, McKenna K, et al. S100A8/A9 proteins mediate neutrophilic inflammation and lung pathology during tuberculosis. American Journal of Respiratory and Critical Care Medicine. 2013;188:1137–1146. doi: 10.1164/rccm.201304-0803OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Anbarasu D, Raja CP, Raja A. Multiplex analysis of cytokines/chemokines as biomarkers that differentiate healthy contacts from tuberculosis patients in high endemic settings. Cytokine. 2013;61:747–754. doi: 10.1016/j.cyto.2012.12.031. [DOI] [PubMed] [Google Scholar]
- 41.Mihret A, Bekele Y, Bobosha K, Kidd M, Aseffa A, Howe R, et al. Plasma cytokines and chemokines differentiate between active disease and non-active tuberculosis infection. The Journal of infection. 2013;66:357–365. doi: 10.1016/j.jinf.2012.11.005. [DOI] [PubMed] [Google Scholar]
- 42.Hilda JN, Narasimhan M, Das SD. Neutrophils from pulmonary tuberculosis patients show augmented levels of chemokines MIP-1alpha, IL-8 and MCP-1 which further increase upon in vitro infection with mycobacterial strains. Human immunology. 2014;75:914–922. doi: 10.1016/j.humimm.2014.06.020. [DOI] [PubMed] [Google Scholar]
- 43.Flores-Villanueva PO, Ruiz-Morales JA, Song CH, Flores LM, Jo EK, Montano M, et al. A functional promoter polymorphism in monocyte chemoattractant protein-1 is associated with increased susceptibility to pulmonary tuberculosis. The Journal of experimental medicine. 2005;202:1649–1658. doi: 10.1084/jem.20050126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Yu Y, Zhang Y, Hu S, Jin D, Chen X, Jin Q, et al. Different patterns of cytokines and chemokines combined with IFN-gamma production reflect Mycobacterium tuberculosis infection and disease. PLoS One. 2012;7:e44944. doi: 10.1371/journal.pone.0044944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Almeida Cde S, Abramo C, Alves CC, Mazzoccoli L, Ferreira AP, Teixeira HC. Antimycobacterial treatment reduces high plasma levels of CXC-chemokines detected in active tuberculosis by cytometric bead array. Mem Inst Oswaldo Cruz. 2009;104:1039–1041. doi: 10.1590/s0074-02762009000700018. [DOI] [PubMed] [Google Scholar]
- 46.Tang NL, Fan HP, Chang KC, Ching JK, Kong KP, Yew WW, et al. Genetic association between a chemokine gene CXCL-10 (IP-10, interferon gamma inducible protein 10) and susceptibility to tuberculosis. Clinica chimica acta; international journal of clinical chemistry. 2009;406:98–102. doi: 10.1016/j.cca.2009.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kaplan G, Post FA, Moreira AL, Wainwright H, Kreiswirth BN, Tanverdi M, et al. Mycobacterium tuberculosis growth at the cavity surface: a microenvironment with failed immunity. Infect Immun. 2003;71:7099–7108. doi: 10.1128/IAI.71.12.7099-7108.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ong CWM, Elkington P, Friedland JS. Tuberculosis, Pulmonary Cavitation, and Matrix Metalloproteinases. American Journal of Respiratory and Critical Care Medicine. 2014;190:9–18. doi: 10.1164/rccm.201311-2106PP. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Chan ED, Chan J, Schluger NW. What is the role of nitric oxide in murine and human host defense against tuberculosis?Current knowledge. Am J Respir Cell Mol Biol. 2001;25:606–612. doi: 10.1165/ajrcmb.25.5.4487. [DOI] [PubMed] [Google Scholar]
- 50.Florido M, Cooper AM, Appelberg R. Immunological basis of the development of necrotic lesions following Mycobacterium avium infection. Immunology. 2002;106:590–601. doi: 10.1046/j.1365-2567.2002.01459.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Oddo M, Renno T, Attinger A, Bakker T, MacDonald HR, Meylan PR. Fas ligand-induced apoptosis of infected human macrophages reduces the viability of intracellular Mycobacterium tuberculosis. J Immunol. 1998;160:5448–5454. [PubMed] [Google Scholar]
- 52.Riendeau CJ, Kornfeld H. THP-1 cell apoptosis in response to Mycobacterial infection. Infect Immun. 2003;71:254–259. doi: 10.1128/IAI.71.1.254-259.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Volkman HE, Pozos TC, Zheng J, Davis JM, Rawls JF, Ramakrishnan L. Tuberculous granuloma induction via interaction of a bacterial secreted protein with host epithelium. Science. 2010;327:466–469. doi: 10.1126/science.1179663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Elkington P, Shiomi T, Breen R, Nuttall RK, Ugarte-Gil CA, Walker NF, et al. MMP-1 drives immunopathology in human tuberculosis and transgenic mice. J Clin Invest. 2011;121:1827–1833. doi: 10.1172/JCI45666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hernandez-Pando R, Orozco H, Arriaga K, Pavon L, Rook G. Treatment with BB-94, a broad spectrum inhibitor of zinc-dependent metalloproteinases, causes deviation of the cytokine profile towards type-2 in experimental pulmonary tuberculosis in Balb/c mice. International journal of experimental pathology. 2000;81:199–209. doi: 10.1046/j.1365-2613.2000.00152.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Izzo AA, Izzo LS, Kasimos J, Majka S. A matrix metalloproteinase inhibitor promotes granuloma formation during the early phase of Mycobacterium tuberculosis pulmonary infection. Tuberculosis (Edinb) 2004;84:387–396. doi: 10.1016/j.tube.2004.07.001. [DOI] [PubMed] [Google Scholar]
- 57.Taylor JL, Hattle JM, Dreitz SA, Troudt JM, Izzo LS, Basaraba RJ, et al. Role for matrix metalloproteinase 9 in granuloma formation during pulmonary Mycobacterium tuberculosis infection. Infect Immun. 2006;74:6135–6144. doi: 10.1128/IAI.02048-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lurie MB. Experimental Epidemiology of Tuberculosis : The Route of Infection in Naturally Acquired Tuberculosis of the Guinea Pig. J Exp Med. 1930;51:769–776. doi: 10.1084/jem.51.5.769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Converse PJ, Dannenberg AM, Jr, Estep JE, Sugisaki K, Abe Y, Schofield BH, et al. Cavitary tuberculosis produced in rabbits by aerosolized virulent tubercle bacilli. Infect Immun. 1996;64:4776–4787. doi: 10.1128/iai.64.11.4776-4787.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Subbian S, Tsenova L, O’Brien P, Yang G, Koo MS, Peixoto B, et al. Phosphodiesterase-4 inhibition combined with isoniazid treatment of rabbits with pulmonary tuberculosis reduces macrophage activation and lung pathology. The American journal of pathology. 2011;179:289–301. doi: 10.1016/j.ajpath.2011.03.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Zhukova NL, Gedymin LE, Golyshevskaia VI. Comparative characteristics of the activity of proteolytic enzymes and their inhibitors in the lung tissue of guinea pigs during development of generalized and destructive tuberculosis. Voprosy meditsinskoi khimii. 1986;32:80–84. [PubMed] [Google Scholar]
- 62.Ugarte-Gil CA, Elkington P, Gilman RH, Coronel J, Tezera LB, Bernabe-Ortiz A, et al. Induced sputum MMP-1, -3 & -8 concentrations during treatment of tuberculosis. PLoS One. 2013;8:e61333. doi: 10.1371/journal.pone.0061333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Van Lint P, Libert C. Chemokine and cytokine processing by matrix metalloproteinases and its effect on leukocyte migration and inflammation. J Leukoc Biol. 2007;82:1375–1381. doi: 10.1189/jlb.0607338. [DOI] [PubMed] [Google Scholar]
- 64.McQuibban GA, Butler GS, Gong JH, Bendall L, Power C, Clark-Lewis I, et al. Matrix metalloproteinase activity inactivates the CXC chemokine stromal cell-derived factor-1. J Biol Chem. 2001;276:43503–43508. doi: 10.1074/jbc.M107736200. [DOI] [PubMed] [Google Scholar]
- 65.Van den Steen PE, Proost P, Wuyts A, Van Damme J, Opdenakker G. Neutrophil gelatinase B potentiates interleukin-8 tenfold by aminoterminal processing, whereas it degrades CTAP-III, PF-4, and GRO-alpha and leaves RANTES and MCP-2 intact. Blood. 2000;96:2673–2681. [PubMed] [Google Scholar]
- 66.Van den Steen PE, Husson SJ, Proost P, Van Damme J, Opdenakker G. Carboxyterminal cleavage of the chemokines MIG and IP-10 by gelatinase B and neutrophil collagenase. Biochemical and biophysical research communications. 2003;310:889–896. doi: 10.1016/j.bbrc.2003.09.098. [DOI] [PubMed] [Google Scholar]
- 67.McQuibban GA, Gong JH, Wong JP, Wallace JL, Clark-Lewis I, Overall CM. Matrix metalloproteinase processing of monocyte chemoattractant proteins generates CC chemokine receptor antagonists with anti-inflammatory properties in vivo. Blood. 2002;100:1160–1167. [PubMed] [Google Scholar]
- 68.Van Den Steen PE, Wuyts A, Husson SJ, Proost P, Van Damme J, Opdenakker G. Gelatinase B/MMP-9 and neutrophil collagenase/MMP-8 process the chemokines human GCP-2/CXCL6, ENA-78/CXCL5 and mouse GCP-2/LIX and modulate their physiological activities. European journal of biochemistry/FEBS. 2003;270:3739–3749. doi: 10.1046/j.1432-1033.2003.03760.x. [DOI] [PubMed] [Google Scholar]
- 69.Li Q, Park PW, Wilson CL, Parks WC. Matrilysin shedding of syndecan-1 regulates chemokine mobilization and transepithelial efflux of neutrophils in acute lung injury. Cell. 2002;111:635–646. doi: 10.1016/s0092-8674(02)01079-6. [DOI] [PubMed] [Google Scholar]
- 70.Nolan A, Condos R, Huie ML, Dawson R, Dheda K, Bateman E, et al. Elevated IP-10 and IL-6 from bronchoalveolar lavage cells are biomarkers of non-cavitary tuberculosis. Int J Tuberc Lung Dis. 2013;17:922–927. doi: 10.5588/ijtld.12.0610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Pichugin AV, Yan BS, Sloutsky A, Kobzik L, Kramnik I. Dominant role of the sst1 locus in pathogenesis of necrotizing lung granulomas during chronic tuberculosis infection and reactivation in genetically resistant hosts. The American journal of pathology. 2009;174:2190–2201. doi: 10.2353/ajpath.2009.081075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Carr MW, Roth SJ, Luther E, Rose SS, Springer TA. Monocyte chemoattractant protein 1 acts as a T-lymphocyte chemoattractant. Proc Natl Acad Sci U S A. 1994;91:3652–3656. doi: 10.1073/pnas.91.9.3652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Pokkali S, Das SD. Augmented chemokine levels and chemokine receptor expression on immune cells during pulmonary tuberculosis. Human immunology. 2009;70:110–115. doi: 10.1016/j.humimm.2008.11.003. [DOI] [PubMed] [Google Scholar]
- 74.Xu LL, Warren MK, Rose WL, Gong W, Wang JM. Human recombinant monocyte chemotactic protein and other C-C chemokines bind and induce directional migration of dendritic cells in vitro. J Leukoc Biol. 1996;60:365–371. doi: 10.1002/jlb.60.3.365. [DOI] [PubMed] [Google Scholar]
- 75.Vesosky B, Rottinghaus EK, Stromberg P, Turner J, Beamer G. CCL5 participates in early protection against Mycobacterium tuberculosis. J Leukoc Biol. 2010;87:1153–1165. doi: 10.1189/jlb.1109742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Freeman CM, Stolberg VR, Chiu BC, Lukacs NW, Kunkel SL, Chensue SW. CCR4 participation in Th type 1 (mycobacterial) and Th type 2 (schistosomal) anamnestic pulmonary granulomatous responses. J Immunol. 2006;177:4149–4158. doi: 10.4049/jimmunol.177.6.4149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Telenti A. Safety concerns about CCR5 as an antiviral target. Curr Opin HIV AIDS. 2009;4:131–135. doi: 10.1097/COH.0b013e3283223d76. [DOI] [PubMed] [Google Scholar]
- 78.Stolberg VR, Chiu BC, Martin BE, Shah SA, Sandor M, Chensue SW. Cysteine-cysteinyl chemokine receptor 6 mediates invariant natural killer T cell airway recruitment and innate stage resistance during mycobacterial infection. Journal of innate immunity. 2011;3:99–108. doi: 10.1159/000321156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Acosta-Rodriguez EV, Rivino L, Geginat J, Jarrossay D, Gattorno M, Lanzavecchia A, et al. Surface phenotype and antigenic specificity of human interleukin 17-producing T helper memory cells. Nat Immunol. 2007;8:639–646. doi: 10.1038/ni1467. [DOI] [PubMed] [Google Scholar]
- 80.Cyster JG. Chemokines and cell migration in secondary lymphoid organs. Science. 1999;286:2098–2102. doi: 10.1126/science.286.5447.2098. [DOI] [PubMed] [Google Scholar]
- 81.Bizzarri C, Beccari AR, Bertini R, Cavicchia MR, Giorgini S, Allegretti M. ELR+ CXC chemokines and their receptors (CXC chemokine receptor 1 and CXC chemokine receptor 2) as new therapeutic targets. Pharmacology & therapeutics. 2006;112:139–149. doi: 10.1016/j.pharmthera.2006.04.002. [DOI] [PubMed] [Google Scholar]
- 82.Goncalves AS, Appelberg R. The involvement of the chemokine receptor CXCR2 in neutrophil recruitment in LPS-induced inflammation and in Mycobacterium avium infection. Scandinavian journal of immunology. 2002;55:585–591. doi: 10.1046/j.1365-3083.2002.01097.x. [DOI] [PubMed] [Google Scholar]