

Abstract
[Purpose]
The aim of this study is to examine the association between the distribution of ACTN3 genotypes and alleles in power, speed, and strength-oriented athletics.
[Methods]
ACTN3 genotyping was carried out for a total of 975 Korean participants: top-level sprinters (n = 58), top-level strength athletes (n = 63), and healthy controls (n = 854).
[Results]
Genetic associations were evaluated by chi-squire test or Fisher’s exact test. In the power-oriented group composed of sprinters and strength athletes, the frequency of the XX genotype was significantly underrepresented (11.6%) in comparison to its representation in the control group (11.6% versus 19.1%, P < 0.05). When the power-oriented group was divided into strength-oriented and speed-oriented groups, no significant difference in the ACTN3 XX genotype was found between the strength-oriented athletes and the controls (15.9% versus 19.1%, P < 0.262). Only the speed-oriented athletes showed significant differences in the frequency distributions of the ACTN3 XX genotype (6.9% versus 19.1%, P < 0.05) from that of the controls.
[Conclusion]
The ACTN3 genotype seems to mainly affect sports performance and especially speed.
Keywords: α-actinin-3, human performance, population genetics, fast-twitch muscle fibers, contractile property
INTRODUCTION
Muscle power is defined as high-intensity muscular performance in a short term and is composed of speed and strength. Muscle power and its components are determined by environmental and genetic factors. Both familial and twin studies suggest that genetic factors in power, speed and strength account for 35%~80% of inter-individual variation [6,28-30]. During the last two decades, there has been considerable interest in finding the genetic factors responsible for human performance. It has recently been reported that some candidate genes are related to human muscle power [19,20]. Among them, ACTN3 is one of the most interesting genes in that the ACTN3 variant (ACTN3 R577X SNP) directly determines the expression of the α-actinin-3 protein that contributes to the construction of the contractile component in power-generating fast twitch fibers of the skeletal muscle [2,23]. In addition, it has been reported that the functional individual difference of α-actnin-3 protein expressed only in sarcomere of fast-twitch muscle fibers is not to be decided by multiple gene (170 counts) diversity coding α-actinin-3 but by ACTN3 R577X diversity such as dominant-recessive effect [16,18].
ACTN3, the gene for encoding α-actinin-3, is located at 11q13-q14. This gene has a 1747C>T transition within exon 16, which results in the ACTN3 R577X SNP, giving three genotypes: the RR, RX and XX genotypes. In the ACTN3 variant (R577X SNP), the XX genotype deficits α-actinin-3 due to the premature stop codon without a pathological phenotype, whereas the RR and RX genotypes express α-actinin-3. As a result, the ACTN3 variant directly determines the expression of α-actinin-3 [1,18]. An important structural component of the Z disc is α-actinin-3, where it anchors actin thin filaments, helping to maintain the myofibrillar array only in fast twitch muscle fibers that are responsible for high velocity and force for power-generating contractions [1,2,18]. It has been suggested that the ACTN3 variant for the expression or deficiency of α-actinin-3 in fast-twitch muscle fibers should influence the power-generating muscle function [2,23].
Several studies have found that the ACTN3 variant affects the top-level elite athletes, and controls speed/power performance [5,7,17,20,24,27,32]. The R allele and the RR genotype, expressing α-actinin-3, are overrepresented whereas the XX genotype for α-actinin-3 deficiencies is underrepresented in sprint events, suggesting that α-actinin-3 is required for sprint/power performance. The α-actinin-3 deficiency has been associated with poorer sprinting performance in non-athletes. Experimental studies using ACTN3 KO mice have also shown that the absence of α-actinin-3 results in fast-twitch, glycolytic fibers developing slower-twitch and poorer power generation [4,15,19], supporting the responsiveness of the α-actinin-3 expression to optimal sprint and power performance. These results show that the ACTN3 variant is somewhat responsible for speed performance and also for determining some aspects of muscle power.
However, previous research on the ACTN3 variant focused mainly on speed performance without differentiating its influence on the power components of speed and strength. Because muscle power is the high-strength muscular performance that occurs over a short time period, it is composed of speed and strength each of which can be differentiated independently and can also be determined by environmental and genetic factors [6,28,29,30]. There is, however, a dearth of information on the association of strength, speed, or power with the ACTN3 variant. Only association of power with the ACTN3 variant and association of speed with the ACTN3 variant are reported [14,31]. Considering that the expression of sarcomeric α-actinin-3 is located at the fast-twitch fiber that generates strong, high-velocity muscle contraction, there is a strong need to understand whether α-actinin-3 is expressed or not and whether it should be associated with forceful contraction, i.e. muscle strength, for giving explosive power.
Therefore, the aim of this study is to examine the distribution of the ACTN3 R577X genotypes and alleles in poweroriented, speed-oriented and strength-oriented athletes and to search for differential associations of the power components with the ACTN3 R577X polymorphism.
METHODS
Participants
All participants in this study were recruited from August, 1996 to December, 2011. All athletes in this study were members of the Korean National team and university elite athletes and the control group consisted of non-athletes participating in national physical fitness survey and national health promotion business. Athletic participants in this study consisted of 849 elite athletes that play 11 different sports including fencing, skating, track & field, swimming, wrestling, boxing, cycling, weightlifting, gymnastics, field hockey and judo. Only weightlifters, sprinters (≤ 400 m), speed skaters (≤ 1,500 m) and swimmers (≤ 100 m) were selected for this study since these sports are mainly focused on strength, speed and power. The weightlifters are considered to be strengthoriented athletes while the speed skaters and swimmers are considered to be speed-oriented athletes [3,21,26]. All three groups (weightlifters, speed skaters and swimmers) belong to the power-oriented category [3,21,26]. As many as 121 top-level athletes participated in this study and their athletic performance records were confirmed by the Korea Sports Council.
A total of 975 Koreans aged 18-39yrs, and composed of healthy controls (n = 854), top-level strength-oriented athletes (n = 63), and top-level speed-oriented athletes (n = 58) were included in this study (see Table 1). Power-oriented top-level athletes (n = 121) were collectively assigned by merging the top-level strength-oriented athletes and the top-level speed-oriented athletes. The 854 healthy controls (435 men, 419 women) met the following inclusion criteria: (1) apparently healthy people (2) no known diseases such as neuromuscular disease, cardiovascular disease or metabolic problems (3) native Korean aged 18-39yrs and (4) no participation in athletic events. The top-level strength-oriented athletes were selected based on their having represented Korea at the international level or on their having won an international championship in weightlifting. Of the final 63 weightlifters (37 men, 26 women) selected as top level strength-oriented athletes for this study, 23 had won medals at the World Championships and Olympic Games. The top-level speedoriented athletes were selected along the same lines i.e. outstanding performance in sprint events and participation in top international competitions and/or Olympic Games as representative Korean sprinters. Of the 58 participants (37 men, 21 women) selected as top-level speed-oriented athletes, 18 were speed skaters (≤ 1,000 m), 21 were sprinters (≤ 400 m) and 19 were swimmers (≤ 100 m). Additionally 19 of them had won medals in their special area either in the Asia or World Championships or the Olympic Games. Written informed consent was obtained from all subjects under protocols approved by the Institutional Review Boards of the Eulji University School of Medicine.
Table 1.
Characteristics of the top-level elite athletes, and controls
| age | Career* | Sport Event | |
|---|---|---|---|
| Strength-oriented (n = 63) | 22.2 ± 3.6 | 3.4 ± 2.9 | Running (≤400, n = 20), |
| Speed skating (≤1,500, n = 20), | |||
| Swimming (≤200, n = 23) | |||
| Speed-oriented (n = 58) | 20.8 ± 4.6 | 2.4 ± 0.8 | Weightlifting (n = 58) |
| Power-orienteda (n =121) | 21.2 ± 4.3 | 2.9 ± 2.1 | above |
| Control (n = 854) | 32.6 ± 4.8 | - | - |
Data are mean and standard deviation (within parentheses).
P-Career as the international athlete, a: the power-oriented athletes are composed of strength-oriented athletes and speed-oriented athletes.
Genotype analysis
Genomic DNA was isolated from peripheral blood cells using a QIAamp blood kit (Qiagen, Valencia, CA, UAS) according to the manufacturer’s protocols. The SNP of the ACTN3 R577X (rs1815739) was analyzed with custom-designed primers and probes (Assay ID: C_590093_1_) for the MGB TaqMan® SNP genotyping assay (Applied Biosystems, Foster City, CA, USA). The primers are Forward: 5'-ACGATCAGTTCAAGGCAACACT-3' and Reverse: 5'-ACCCTGGAT GCCCATGATG-3'. Allele-specific probes were labeled with the fluorescent dyes VIC (5'-TCGCTCTCGGTCAGC- 3') and FAM (5'-CGCTCTCAGTCAGC-3'). RCR was carried out in a total reaction volume of 20㎕ containing 200ng of template, 10㎕ TaqMan Universal PCR Master Mix No AmpErase UNG (2x) and 0.5㎕ 40xSNP Genotyping Assay with the following amplification protocol: denaturation at 95℃ for 10 minutes, followed by 40 cycles of denaturation at 92℃ for 15 seconds and finally annealing and extension at 60℃ for 1 minute. Post PCR, the genotype of each sample was attributed automatically by measuring the allele-specific fluorescence on the ABI PRISM® 7900 Sequence Detection System using SDS 2.3 software for allelic discrimination (Applied Biosystems, Foster City, CA, USA). Duplicate samples and negative controls were included to ensure accuracy of the genotyping.
Statistical analysis
PAWE (Power for Association with Error) software program was used to test sample size validity and the sample size validity was confirmed by having type 1 error set at 0.05 and type II error as 80% power level. The ACTN3 allele and genotype frequencies were obtained by direct count. The Hardy-Weinberg equilibrium (HWE) for genotype distribution of each group was then estimated by the chi-square test. Differences in the allele frequencies and genotypes between the controls and the power-oriented athletes, between the controls and the top-level strength athletes and between the controls and the top-level sprinters were compared using the chi-square test or Fisher’s exact test based on the presence of less than five frequencies in one cell. Trend analysis for the genotype distribution between the controls and the power-oriented, top-level strength and speed groups was conducted by the linear-by-linear association analysis. All analyses were conducted using SPSS statistical software (Version 13.0; SPSS Inc., Chicago, IL). A two-tailed significance at .05 level was chosen to test for type I errors.
RESULTS
A combination of control individuals and top-level power-oriented athletes, divided into top-level weightlifters (strength-oriented) and sprinters (speed-oriented), was analyzed in this study. Table 1 shows the ACTN3 allele and genotype frequen cies from the control and power-oriented groups. The ACTN3 genotype distributions in the control group and the poweroriented group were in Hardy-Weingberg equilibrium (χ2[df=1] = 0.934 for the control, χ2[df=1] = 0.258 for the power-oriented group; p > 0.10). The ACTN3 genotype and allele distribution in the power-oriented athletes showed significant differences when compared with the controls (χ2[df=2] = 7.347, P < 0.05 for genotype; χ2[df=1] = 7.105, P < 0.01 for allele). The poweroriented group had an underrepresented frequency of the XX genotype (11.6% vs 19.1%) and the X allele (35.5% vs 44.6%) in comparison to the control group.
To test the association of power types (strength and speed performance) with the ACTN3 genotype, the power-oriented group was divided into the strength-oriented group (the toplevel weightlifters) and the speed-oriented group (the top-level sprinters). Both of them were in HWE (χ2[df=1] = 1.758 for speed-oriented athletes, χ2[df=1] = 0.210 for strength-oriented athletes; P > 0.10). As shown in Table 1, no statistical differences in the ACTN3 genotype and allele distributions were found between the strength-oriented athletes and the controls (χ2[df=2] = 2.682, P < 0.262 for genotype; χ2[df=1] = 2.022, P = 0.155 for allele). Only the speed-oriented athletes showed significant differences in the frequency distributions of the ACTN3 genotype (Fisher’s Exact Test = 7.252, P < 0.05) and the allele (χ2[df=1] = 6.200, P < 0.05) from that of the controls. In Fig. 1, the frequency of the α-actinin-3 non-expression genotype (XX) was significantly reduced while the frequency of the α-actinin-3 expression genotype (RR + RX) was significantly increased in the speed-oriented athletes compared with the controls (Fisher’s Exact Test = 5.395, P < 0.05; Fig. 1). No significant difference in the α -actinin-3 expression genotype distribution was found in the strength-oriented athletes compared with the controls (χ2[df=1] = 0.396, P < 0.529).
Fig. 1.
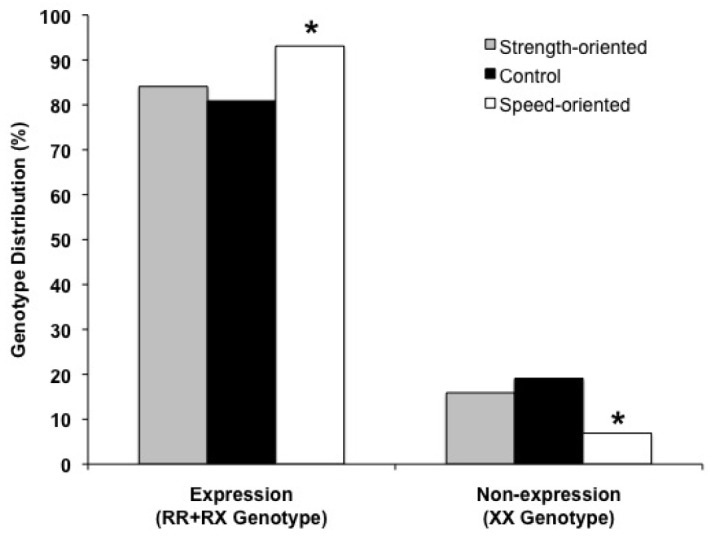
Distribution of α-actinin-3 expression and non-expression genotype in the top-level strength-oriented athletes, speed-oriented athletes, and controls
* means significantly different in genotype distribution from the control at P < 0.05
DISCUSSION
The aim of this study is to explore an association of the ACTN3 genotype with speed and/or strength performance. The ACTN3 genotype distribution of Korean top-level sprinters for speed, weightlifters for strength and mixed power-oriented athletes for power were compared to the ACTN3 genotype distribution of Korean adults. There was a significant association of the ACTN3 genotype with power performance as was expected. When dividing power performance into speed and strength performances, the ACTN3 genotype was only associated with speed performance but a relationship with strength performance was not identified. The ACTN3 XX genotype in deficit of the α-actinin-3 protein in fast-twitch muscle fibers had a significantly low distribution in the speed performance group, while the distribution of the ACTN3 XX genotype showed a somewhat low ratio compared to the control group in the strength performance group with no statistical significance. Therefore, speed performance among the power factors showed preference for the ACTN3 RR + RX genotype but not for the ACTN3 XX genotype.
The association of the ACTN3 genotype with speed performance in this study is similar to previous study results. Yang et al. [32] reported reduced distribution of the ACTN3 XX genotype in sprint/power groups, composed of 7 Australian Judo elite athletes and 42 sprinters. Niemi and Majamaa [17] and Papadimitrou et al. [20] reported remarkably low levels of the ACTN3 XX genotype in Greek sprinters. In these studies, the distributions of the ACTN3 XX genotype in the control groups were about 18% similar to the 19% distribution fur the control group in our study. Also, the authors reported low XX genotype distributions for sprinters (6%, 0%, and 8.8%) compared to the control groups which agreed with our results of 6.9%. The ACTN3 genotype is favorable to sprint performance while the ACTN3 XX genotype lacking α-actinin-3 is unfavorable to sprint performance.
Druzhevskaya et al. [7] and Papadimitrioum et al. [20], studied Russian and Greek populations respectively and reported that the distributions of the ACTN3 XX genotype were low in the power-oriented performance group compared to their control groups. Santiago [27] also reported a reduced distribution of the XX genotype in Spanish soccer players who needed explosive power through a combination of strength and speed. In contrasr,neither Nigerian nor Greek Track and Field athletes skilled in power-oriented performance [20,33] showed any significant differences in their ACTN3 XX genotype distribution compared to the control groups. Thus, research on the association of ACTN3 with power-oriented performance has not shown consistent results. This may come from differences in the genotype distribution between human populations [33]. It is well-known that the genetic structures of Caucasians and Africans are different, leading to some functionally different characteristics. Actually the XX genotype of the ACTN3 in Caucasian is ~19%, higher than the 10% found in Africans. In addition, the differences in the composition of speed and strength athletes skilled in power-oriented performance may be another cause of the inconsistent association of ACTN3 with power performance [16,18,33]. This study divided the ACTN3-associated power-oriented performance into speed performance and strength performance but the ACTN3 genotype distribution showed association only with speed performance and none with the strength performance. The coordination level of speed and strength affecting power-oriented performance may play an important role in determining the association between ACTN3 and power. As a result, the association of the ACTN3 R577X variant with power-oriented performance would not show consistency and a variant that distinguishes the components of power into speed and strength would be needed to find the association of the ACTN3 variant with athletic performance.
As there was no significant difference in the ACTN3 genotype distribution of Korean top-level weightlifters with that of the control group in this study, the ACTN3 variant had no significant association with strength performance. Because weightlifting requires a single maximal contraction of the skeletal muscle, it is essential for the skeletal muscle, especially for fast-twitch muscle fibers, to be hypertrophic [8,9,11,12]. It is expected that the distribution of the ACTN XX genotype was low because in the ACTN XX genotype, α-actinin-3 is structured in the Z-line of the fast-twitch muscle fiber that functions during rapid speed and strong power [13,31]. However, in contrast with this hypothesis, the ACTN3 XX genotype was not significantly reduced in weightlifters. Such a result corresponded to previous study results with isokinetic strength performance of active young men and women. There was no difference in the isokinetic torque and rate of decline of torque between ACTN3 genotypes [10]. These results including ours could support the contention that the lack of α-actinin-3 in fast-twitch muscle fibers by the XX genotype only slightly affects the muscular strength of a single maximum contraction. On the other hand, the null type of the α-actinin-3, ACTN3 XX genotype, showed a significantly low distribution in speed performance as consistently reported in previous studies and also in this study. Therefore, it is thought that the α-actinin-3 deficient genotype, the ACTN3 XX genotype, may affect speed performance that needs rapidly repeated muscular contractions. In the future, it is necessary to explore the mechanism that causes the difference in the contraction of fast-twitch muscles that is due to the lack of α-actinin-3. In the case where the lack of α-actinin-3 has no relation with the maximum muscular strength, there is also the possibility that another protein can compensate for this lack. This mechanism also requires further study.
CONCLUSIONS
The ACTN3 XX genotype that is in deficit of α-actinin-3 is confirmed to be strongly associated with speed performance that requires multiply repeated rapid muscle contractions. The ACTN3 XX genotype had no association with strength performance for single maximum muscle contractions among power components. Therefore, we found suggestive evidence that the ACTN3 R577X polymorphism plays a role in discriminating speed performance athletes from strength athletes and sedentary controls. In other words, the ACTN3 R577X variant is the main gene for speed performance but not for strength performance.
Table 2.
The distribution of the ACTN3 R577X genotype and the allele frequency in the top-level power-oriented athletes, and controls
| Group | Genotype |
Allele |
|||||
|---|---|---|---|---|---|---|---|
| RR | RX | XX | P* | R | X | P* | |
| Control (n = 854) | 255 (29.9) | 436 (51.1) | 163 (19.1) | 946 (55.4) | 762 (44.6) | ||
| Power-orienteda (n = 121) | 49 (40.5) | 58 (47.9) | 14 (11.6) | .025 | 156 (64.5) | 86 (35.5) | .008 |
Data are absolute and relative values (within parentheses).
P-value for the χ2 test or Fisher’s Exact Test in comparison to controls. a: the power-oriented athletes are composed of strength-oriented athletes and speed-oriented athletes.
Table 3.
The distribution of the ACTN3 R577X genotype and the allele frequency in the top-level sprinters, weightlifters, and controls
| Group | Genotype |
Allele |
|||||
|---|---|---|---|---|---|---|---|
| RR | RX | XX | P* | R | X | P* | |
| Control (n = 854) | 255 (29.9) | 436 (51.1) | 163 (19.1) | 946 (55.4) | 762 (44.6) | ||
| Strength-oriented (n = 63) | 25 (39.7) | 28 (44.4) | 10 (15.9) | .262 | 78 (61.9) | 48 (38.1) | .155 |
| Speed-oriented (n = 58) | 24 (41.4) | 30 (51.7) | 4 (6.9) | .026 | 78 (67.2) | 38 (32.8) | .013 |
Data are absolute and relative values (within parentheses).
P-value for the χ2 test or Fisher’s Exact Test in comparison to controls.
Acknowledgments
We would like to thank the Korean Olympic Committee and all the participants for their invaluable assistance.
REFERENCES
- 1.Beggs AH, Byers TJ, Knoll JH, Boyce FM, Bruns GA, Kunkel LM. Cloning and characterization of two human skeletal muscle alpha-actinin genes located on chromosomes 1 and 11. J Biol Chem. 1992;267(13):9281–9288. [PubMed] [Google Scholar]
- 2.Beggs Y, North KN. A gene for speed: The emerging role of alpha-actinin-3 in muscle metabolism. Physiology. 2010;25(4):250–259. doi: 10.1152/physiol.00008.2010. [DOI] [PubMed] [Google Scholar]
- 3.Brown J. Sports Talent: how to identify and develop outstanding athletes. Human Kinetics; Champaign, IL: 2001. [Google Scholar]
- 4.Chan S, Seto JT, Houweling PJ, Yang N, North KN, Head SI. Properties of extensor digitorum longus muscle and skinned fibers from adult and aged male and female ACTN3 knockout mice. Muscle & Nerve. 2011;43(1):37–48. doi: 10.1002/mus.21778. [DOI] [PubMed] [Google Scholar]
- 5.Chiu LL, Wu YF, Tang MT, Yu HC, Hsieh LL, Hsieh SS. ACTN3 genotype and swimming performance in Taiwan. Int J Sports Med. 2011;32(6):476–480. doi: 10.1055/s-0030-1263115. [DOI] [PubMed] [Google Scholar]
- 6.De Moor MH, Spector TD, Cherkas LF, Falchi M, Hottenga JJ, Boomsma DI, De Geus EJ. Genome-wide linkage scan for athlete status in 700 British female DZ twin pairs. Twin Res Hum Genet. 2011;10(6):812–820. doi: 10.1375/twin.10.6.812. [DOI] [PubMed] [Google Scholar]
- 7.Druzhevskaya AM, Ahmetov II, Astratenkova IV, Rogozkin VA. Association of the ACTN3 R577X polymorphism with power athlete status in Russians. Euro J App Physiol. 2011;103(6):631–634. doi: 10.1007/s00421-008-0763-1. [DOI] [PubMed] [Google Scholar]
- 8.Fry AC, Schilling BK, Staron RS, Hagerman FC, Hikida RS, Thrush JT. Muscle fiber characteristics and performance correlates of male olympic-style weightlifters. J Strength Cond Res. 2003;17(4):746–754. doi: 10.1519/1533-4287(2003)017<0746:mfcapc>2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 9.Haff GG, Carlock JM, Hartman MJ, Kilgore JL, Kawamori N, Jackson JR, Stone MH. Force-time curve characteristics of dynamic and isometric muscle actions of elite women olympic weightlifters. J Strength Cond Res. 2005;19(4):741–748. doi: 10.1519/R-15134.1. [DOI] [PubMed] [Google Scholar]
- 10.Hanson ED, Ludlow AT, sheaf AK, Park J, Roth SM. ACTN3 Genotype does not influence muscle power. 2010;31(11):834–838. doi: 10.1055/s-0030-1263116. [DOI] [PubMed] [Google Scholar]
- 11.Ikegawa S, Funato K, Tsunoda N, Kanehisa H, Fukunaga T, Kawakami Y. Muscle force per cross-sectional area is inversely related with pennation angle in strengthtrained athletes. J Strength Cond Res. 2008;22(1):128–131. doi: 10.1519/JSC.0b013e31815f2fd3. [DOI] [PubMed] [Google Scholar]
- 12.Izquierdo M, Ibanez J, Hakkinen K, Kraemer WJ, Ruesta M, Gorostiaga EM. Maximal strength and power, muscle mass, endurance and serum hormones in weightlifters and road cyclists. Journal of Sports Sciences. 2004;22(5):465–478. doi: 10.1080/02640410410001675342. [DOI] [PubMed] [Google Scholar]
- 13.MacArthur DG, North KN. A gene for speed? the evolution and function of alpha-actinin-3. BioEssays. 2004;26(7):786–795. doi: 10.1002/bies.20061. [DOI] [PubMed] [Google Scholar]
- 14.MacArthur DG, North KN. ACTN3: A genetic influence on muscle function and athletic performance. Exer Sport Sci Rev. 2007;35(1):30–34. doi: 10.1097/JES.0b013e31802d8874. [DOI] [PubMed] [Google Scholar]
- 15.MacArthur DG, Seto JT, Chan S, Quinlan KG, Raftery JM, Turner N, North KN. An ACTN3 knockout mouse provides mechanistic insights into the association between alpha-actinin-3 deficiency and human athletic performance. Hum Mol Genet. 2008;17(8):1076–1086. doi: 10.1093/hmg/ddm380. [DOI] [PubMed] [Google Scholar]
- 16.Mills M, Yang N, Weinberger R, Vander Woude DL, Beggs AH, Easteal S, North K. Differential expression of the actin-binding proteins, alpha-actinin-2 and -3, in different species: Implications for the evolution of functional redundancy. Hum Mol Genet. 2001;10(13):1335–1346. doi: 10.1093/hmg/10.13.1335. [DOI] [PubMed] [Google Scholar]
- 17.Niemi AK, Majamaa K. Mitochondrial DNA and ACTN3 genotypes in finnish elite endurance and sprint athletes. Euro J Hum Genet. 2005;13(8):965–969. doi: 10.1038/sj.ejhg.5201438. [DOI] [PubMed] [Google Scholar]
- 18.North North, Yang N, Wattanasirichaigoon D, Mills M, Easteal S, Beggs AH. A common nonsense mutation results in alpha-actinin-3 deficiency in the general population. Nature Genet. 1999;21(4):353–354. doi: 10.1038/7675. [DOI] [PubMed] [Google Scholar]
- 19.Ogura Y, Naito H, Kakigi R, Ichinoseki-Sekine N, Kurosaka M, Katamoto S. Alpha-actinin-3 levels increase concomitantly with fast fibers in rat soleus muscle. Biochem Biophy Res Commun. 2008;372(4):584–588. doi: 10.1016/j.bbrc.2008.05.059. [DOI] [PubMed] [Google Scholar]
- 20.Papadimitriou ID, Papadopoulos C, Kouvatsi A, Triantaphyllidis C. The ACTN3 gene in elite Greek track and field athletes. Int J Sports Med. 2008;29(4):352–355. doi: 10.1055/s-2007-965339. [DOI] [PubMed] [Google Scholar]
- 21.Powers SK, Howley ET. Bioenergenics in Exercise Physiology. 4th ed. McGrawHill; Dubuqe IL: 2001. pp. 22–46. [Google Scholar]
- 22.Puthucheary Z, Skipworth JR, Rawal J, Loosemore M, Van Someren K, Montgomery HE. Genetic influences in sport and physical performance. Sports Med. 2011;41(10):845–859. doi: 10.2165/11593200-000000000-00000. [DOI] [PubMed] [Google Scholar]
- 23.Rankinen T, Roth SM, Bray MS, Loos R, Perusse L, Wolfarth B, Bouchard C. Advances in exercise, fitness, and performance genomics. Med Sci Sports Exer. 2010;42(5):835–846. doi: 10.1249/MSS.0b013e3181d86cec. [DOI] [PubMed] [Google Scholar]
- 24.Ruiz JR, Arteta D, Buxens A, Artieda M, Gomez-Gallego F, Santiago C, Lucia A. Can we identify a power-oriented polygenic profile? J Appl Physiol. 2010;108(3):561–566. doi: 10.1152/japplphysiol.01242.2009. [DOI] [PubMed] [Google Scholar]
- 25.Ruiz JR, Fernandez Del Valle M, Verde Z, Diez-Vega I, Santiago C, Yvert T, Lucia A. ACTN3 R577X polymorphism does not influence explosive leg muscle power in elite volleyball players. Scnd J Med Sci Sports. 2011;21(6):e34–e41. doi: 10.1111/j.1600-0838.2010.01134.x. [DOI] [PubMed] [Google Scholar]
- 26.Sander D. Sports power. Human Kinetics; Champaign, IL: 2005. [Google Scholar]
- 27.Santiago C, Gonzalez-Freire M, Serratosa L, Morate FJ, Meyer T, Gomez-Gallego F, Lucia A. ACTN3 genotype in professional soccer players. Br J Sports Med. 2008;42(1):71–73. doi: 10.1136/bjsm.2007.039172. [DOI] [PubMed] [Google Scholar]
- 28.Silventoinen K, Magnusson PK, Tynelius P, Kaprio J, Rasmussen F. Heritability of body size and muscle strength in young adulthood: A study of one million Swedish men. Genet Epidem. 2008;32(4):341–349. doi: 10.1002/gepi.20308. [DOI] [PubMed] [Google Scholar]
- 29.Simoneau JA, Bouchard C. Genetic determinism of fiber type proportion in human skeletal muscle. FASEB. 1995;9(11):1091–1095. doi: 10.1096/fasebj.9.11.7649409. [DOI] [PubMed] [Google Scholar]
- 30.Tiainen K, Sipila S, Kauppinen M, Kaprio J, Rantanen T. Genetic and environmental effects on isometric muscle strength and leg extensor power followed up for three years among older female twins. J Appl Physiol. 2009;106(5):1604–1610. doi: 10.1152/japplphysiol.91056.2008. [DOI] [PubMed] [Google Scholar]
- 31.Yang N, Garton F, North K. Alpha-actinin-3 and performance. Med Sport Sci. 2009;54:88–101. doi: 10.1159/000235698. [DOI] [PubMed] [Google Scholar]
- 32.Yang N, MacArthur DG, Gulbin JP, Hahn AG, Beggs AH, Easteal S, North K. ACTN3 genotype is associated with human elite athletic performance. Am J Hum Genet. 2003;73(3):627–631. doi: 10.1086/377590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yang N, MacArthur DG, Wolde B, Onywera VO, Boit MK, Lau SY, North K. The ACTN3 R577X polymorphism in east and West African athletes. Med Sci Sports Exer. 2007;39(11):1985–1988. doi: 10.1249/mss.0b013e31814844c9. [DOI] [PubMed] [Google Scholar]