
Fig 1. Kv channels and their mechanosensitivity.
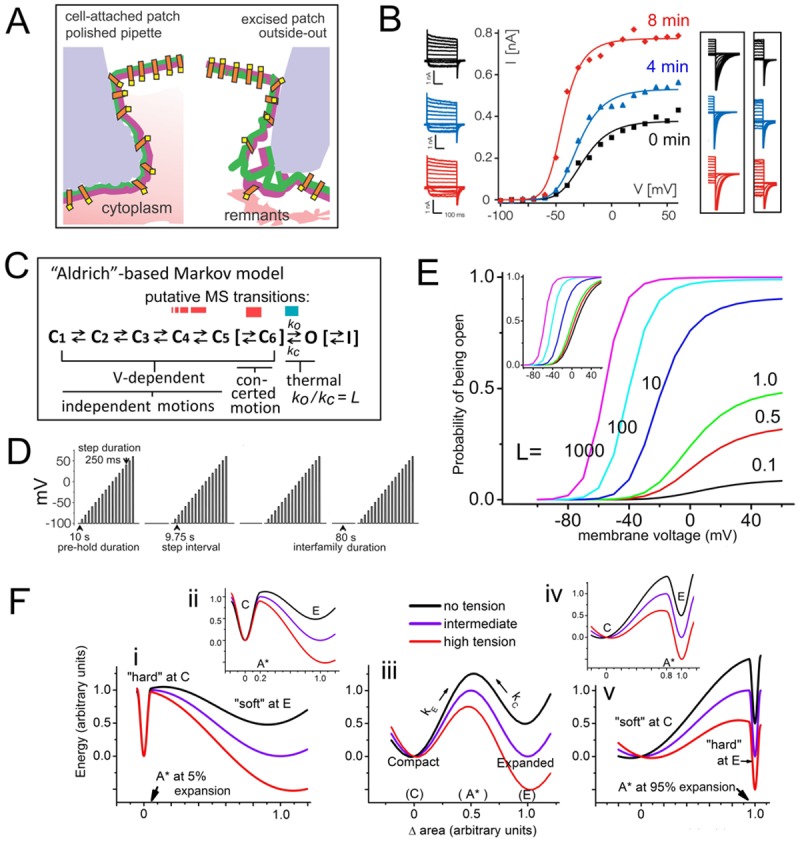
A. Cartoon of some differences at the gigaohm seal zones for (left) cell-attached versus (right) excised outside-out oocyte patches. Left, the uneven surface (pale blue) depicts pipette tip glass sputtered with molten soda glass during fire polishing [18]. Right, outside-out patches (whose structure would resemble that of pinched off blebs or multi-lipid artificial bilayers or [30,31]) reseal within the pipette’s unmodified borosilicate interior surface, then creep inward, developing progressively higher tension [19]. Membrane-glass interactions are extensive and stable at soft-glass sputtered tips (Left) and spontaneous tension build-up is less evident [17]; patches withstand multiple prolonged bouts of applied suction [32,33,34,35]. B. Exemplar data from Schmidt et al [19] (modified from their Fig. 1) showing “conversion” for Kv-Shaker, plus (boxed) tail currents for other Kvs. C. A kinetic scheme for Kv channels, extending Scheme 1 with optional addition (square brackets) of C6 (“pre-open”) and I (slow inactivated) states. Putative MS transitions color-coded red for V-dependent (variants explained in the text), blue for thermal. D. V(t) clamp protocol for acquiring g(V) data (from “tail-IK”): as per [19], except that to confirm trends we simulated acquisition of 4 g(V)s (not 3). E. Effect of varying L on P open (V) (values from Ref. [36]); inset, same curves normalized to P open-max (similar to Fig. 2 of ref [37]). F, i-v. Reaction co-ordinate (drawn by eye) depicting MS transition energetics. Some area, A*, between area, C, of the smaller compacted and area, E, of the larger expanded protein conformation will be the area of the unstable “activated” or transition state [38] (for V-dependent transitions, A*-C compares to the equivalent charge moved through the electric field to put a two-state V-sensor in its transition state; see [37]). A MS channel transition could be between two distinct closed states [38,39], or, as per Schmidt et al [19] a closed and an open state. Activation barriers at the extrema, A* = C and A* = E, would imply extreme rigidities at those locations. Protein energetics depicted here at zero, intermediate and high membrane tensions (i, iii, v) are for A* marginally >C (5% of its expansion), or A* midway between C and E (50% of its expansion), or A* marginally <E (95% of its expansion), with two cases (ii, iv) of moderate barrier asymmetry (20% and 80% area expansion) as insets. In vMS and LMS models here (as in a previous MS-Kv channel model [39]) symmetry is implied (A*midway between C and E) as it is the simplest assumption in the absence of structural data for transition states (see also [38,40,41,42]).