

Abstract
Changes in activated areas of the brain during ankle active dorsiflexion and ankle active plantar flexion were observed in six healthy subjects using functional magnetic resonance imaging. Excited areas of ankle active dorsiflexion involved the bilateral primary motor area and the primary somatosensory area, as well as the bilateral supplementary sensory area, the primary visual area, the right second visual area, and the vermis of cerebellum. Excited areas of ankle active plantar flexion included the ipsilateral supplementary motor area, the limbic system, and the contralateral corpus striatum. Fine movements of the cerebral cortex control the function of the ankle dorsiflexion to a larger extent than ankle plate flexion, and the function of ankle plate flexion is more controlled by the subcortical area.
Keywords: ankle dorsiflexion, ankle plantar flexion, functional magnetic resonance imaging, brain function, activated areas
Abbreviations:
fMRI, functional magnetic resonance imaging; BA, Brodmann area
INTRODUCTION
Ankle joint function plays an essential role in hemiplegic gait. The function of ankle dorsiflexion in many hemiplegic patients slowly recovers, even leading to plantar flexion spasm[1,2,3]. The present study assumed that those symptoms may be associated with different neural mechanisms for controlling the two kinds of movements. Previous studies have demonstrated that cerebral cortex and basal ganglia circuits are important in regulating movements[4,5], but the precise mechanism for this regulation remains poorly understood. Functional magnetic resonance imaging (fMRI) is non-invasive, rapid and convenient for studying human brain function. Studies have shown that ankle dorsiflexion and ankle plantar flexion involve some of the same activated areas in the brain, but also some different activated areas[6,7]. However, it remains unknown whether these differences contribute to variances in recovery of ankle dorsiflexion and ankle plantar flexion following hemiplegia. The present study used fMRI to scan the activated brain areas of healthy patients during ankle dorsiflexion and ankle plantar flexion. This imaging technique was used to analyze neural mechanisms for controlling the studied movements to explain the mechanism of ankle dorsiflexion dysfunction in hemiplegic patients.
RESULTS
Quantitative analysis of participants
A total of six healthy participants were included in the final analysis.
Activated brain areas during ankle dorsiflexion
fMRI showed that during right ankle dorsiflexion, the excited areas included the bilateral paracentral lobule and precuneus, which are mainly located in the left hemisphere, Brodmann area (BA) 1, 2, 4 and 7; bilateral lingual gyrus, which is mainly located in the left hemisphere; BA 17 and 18; right cuneus, BA 18 and 19; right inferior occipital gyrus adjacent to lateral occipitotemporal gyrus and lingual gyrus, BA 18 and 19; and the vermis (Table 1, Figures 1 and 2).
Table 1.
Excited brain areas during right ankle dorsiflexion and plantar flexion

Figure 1.
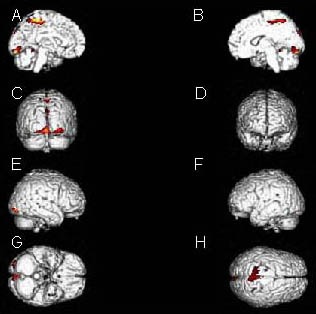
Stereo display of activated brain areas during ankle active dorsiflexion: bilateral paracentral lobule and precuneus, bilateral lingual gyrus, right cuneus, right inferior occipital gyrus adjacent to lateral occipitotemporal gyrus and lingual gyrus, as well as the vermis.
(A) Internal plane of left hemisphere; (B) internal plane of right hemisphere; (C) posterior plane; (D) anterior plane; (E) external place of right hemisphere; (F) external place of left hemisphere; (G) inferior plane; (H) superior plane.
Figure 2.
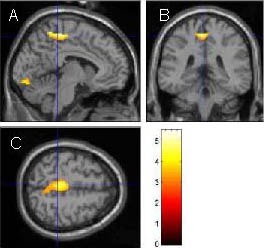
Three-dimensional cross section of activated brain areas during ankle active dorsiflexion.
Sagittal plane (A), coronal plane (B) and transverse plane (C) of paracentral lobule of the left hemisphere. The bright spot shows positive areas. As shown in scale, the light color represents a high possibility of activation.
Activated brain areas during ankle plantar flexion
fMRI showed that during right ankle plantar flexion, the excited areas included the left lenticular nucleus, caudate nucleus; right amygdaloid body and parahippocampal gyrus; right superior frontal gyrus and medial frontal gyrus; and BA 6 region (Table 1, Figures 3 and 4).
Figure 3.
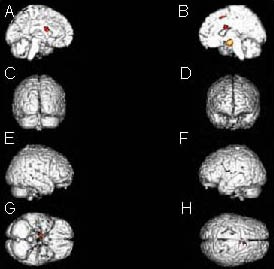
Stereo display of activated brain areas during ankle active plantar flexion: left lenticular nucleus and caudate nucleus; right amygdaloid body and parahippocampal gyrus; right superior frontal gyrus and medial frontal gyrus.
(A) Internal plane of left hemisphere; (B) internal plane of right hemisphere; (C) posterior plane; (D) anterior plane; (E) external place of right hemisphere; (F) external place of left hemisphere; (G) inferior plane; (H) superior plane.
Figure 4.
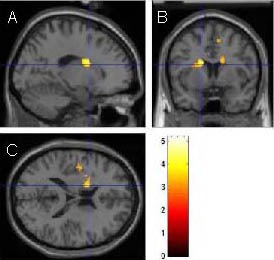
Three-dimensional cross section of activated brain areas during ankle plantar flexion.
Sagittal plane (A), coronal plane (B) and transverse plane (C) of basal ganglia of the left hemisphere. The bright spot shows positive areas. As shown in scale, the light color represents a high possibility of activation.
DISCUSSION
The present study utilized MRI echo planar imaging[8,9] to investigate activated brain areas during ankle dorsiflexion and plantar flexion movements, analyze differences in brain functional activities between the two kinds of movements and explore the mechanism of ankle dorsiflexion dysfunction in hemiplegic patients. Results showed that ankle dorsiflexion more activated the cortex, while ankle plantar flexion more excited the subcortical area.
The central nervous system controls movements through a serial of activities in cortical and subcortical structures. During this process, a large amount of information feedback among structures is required to accurately control movements[10,11,12,13]. In the present study, ankle active dorsiflexion excited areas of the bilateral primary motor area, primary somatosensory area (paracentral lobule) and supplementary sensory area (precuneus), which are mainly located in the left hemisphere, in addition to the bilateral primary visual area (lingual gyrus) and right second visual area (cuneus, right inferior occipital gyrus adjacent to lateral occipitotemporal gyrus and lingual gyrus). The paracentral lobule is the junction of the precentral gyrus and postcentral gyrus on the medial surface of the cerebral cortex. The precentral and postcentral gyri are the primary motor and somatosensory areas, and are in charge of high-level fine motor movements. Studies have shown that 30–40% of fibers in the pyramidal tract originate from the primary motor area, and that all thick fibers (3%) in the pyramidal tract are derived from the primary motor area[11,13,14]. Damage to the primary motor area can affect fine motor movements of the distal limbs[12,13,15]. The primary somatosensory areas are the final sensory areas of fine touch and proprioceptive sense, as well as sensory areas for the source, degree and property of pain, as well as temperature sensation. Damage to the primary somatosensory areas can result in the deletion in fine touch and proprioceptive senses, and but these areas are able to retain general pain, temperature and light touch sensitivity[11]. The supplementary sensory area receives extensive afferent nerve fibers, plays an important role in interaction between individuals and their surroundings[16] and controls movements by transmitting information to parietal and frontal lobe network[17]. Ankle dorsiflexion also activates the primary and secondary visual areas, indicating that ankle dorsiflexion may involve the integration of various stimulations of senses, including vision, and may be controlled through the parietal and frontal lobe network. The noted excitement of various cortexes indicates that a large number of cortexes participate in ankle active dorsiflexion, in particular the primary motor areas.
Results from the present study showed that ankle plantar flexion excited fewer cortical areas compared with ankle dorsiflexion, more in the subcortical area. Ankle active plantar flexion induced excitements in the ipsilateral supplementary motor area, mainly located anterior to the medial of the primary motor area. Stimulation of supplementary motor area can result in complex movements, and plays a role in preparation for specific posture. Only 5% of fibers in the pyramidal tract originate from the supplementary motor area, and a large number of fibers enter the supplementary motor area from basal ganglia[11]. The supplementary motor area is associated with some simple tasks and can check whether the input signals and command correspond to internal demands. Damage to the supplementary motor area does not affect motor function for an extended period of time[11]. In addition, the present study found that ankle active plantar flexion excited contralateral caudate nucleus and lenticular nucleus. The basal ganglia nucleus groups receive projection fibers from the cortex, thalamus and brain stem. Therefore, damage to the basal ganglia can reduce motor coordination ability. Ankle active plantar flexion also induced limbic system excitement, which highly correlates with autonomic nervous system, emotion and memory functions. We thus suggest that the function of ankle plantar flexion is more controlled by senior subcortical areas associated with motor coordination.
Previous brain function studies have shown that a variety of cerebral cortexes and basal ganglia areas are involved in motor control, and that motor learning, programming, correcting and implementing are dominated by a cortex-corpus striatum network circuit[4,5]. An fMRI study showed that ankle dorsiflexion and plantar flexion involved common cortical areas (primary the motor area and supplementary motor area), and other different brain areas[6]. During repetitive ankle dorsiflexion and plantar flexion, the primary motor area, primary somatosensory area, supplementary motor area, premotor cortex area and subcortical areas (putamen) are activated, indicating that senior cortex and subcortical areas participate in controlling ankle motor functions[18,19,20,21,22,23,24,25]. The present study separately investigated ankle dorsiflexion and plantar flexion to prevent the interaction of dorsiflexion and plantar flexion, as both the motor cortex and basal ganglia areas are involved in these movements.
However, ankle dorsiflexion may involve more cortical areas than ankle plantar flexion, in particular cortical areas related to fine motor movements. Clinical studies have shown that ankle dorsiflexion is difficult to recover following central nerve injury, and internal capsule damage can induce severe hemiplegia or spasm, manifested as plantar flexion spasm in the ankle joint[3]. The frontal lobe can be generally divided into primary motor areas and non-primary motor areas. The primary motor areas are responsible for controlling simple and fine movements, and non-primary motor areas contribute to coordinated motion and complex motion.
Results from the present study suggest that ankle dorsiflexion is more controlled by the senior cortex associated with fine motor movements, while ankle plantar flexion is more controlled by subcortical areas and frontal non-primary motor areas. Brunnstrom proposed that motor functions post stroke recover in a specific order; i.e., synergic movement recovers first, followed by senior voluntary movement. This means that fine motor movements restore slowly, but rough synergic movement restore rapidly. Following stroke, ankle dorsiflexion, which is controlled by the senior cortex associated with fine motor movements, slowly recovers, but ankle plantar flexion easily recovers. Therefore, plantar flexion spasm is common in a clinical setting. In conclusion, the brain controls body movements in multiple directions and proportion. Ankle active dorsiflexion is more controlled by the senior cortex associated with fine motor movements, and ankle active plantar flexion is more controlled by subcortical areas and frontal complex movements. Brain injury thus results in different influences on these two kinds of movements.
SUBJECTS AND METHODS
Design
Block design of neuroimaging study.
Time and setting
The experiment was performed at the Magnetic Laboratory, Chinese PLA General Hospital, China, from September 2009 to March 2010.
Subjects
A total of six healthy, male volunteers were staff members of the Plumbing Class, Chinese PLA General Hospital, China, aged 20–25 years. All six volunteers were free of nerve, mental, foot or ankle joint injury. No neurological function impairment was noted by clinical physical or neurological examination; no abnormalities were found in brain structures or signals in routine MRI. Informed consent was obtained from all subjects. The study was performed in accordance with the Declaration of Helsinki.
Methods
Motor and scanning pattern
Motor pattern: the subjects were placed in a dorsal position. The subject’s head was fixed in place using a sponge mat, face mask and bandage. The right foot was raised above the bed via the placement of a soft pad under the right leg. The subject was asked to relax and close his eyes; his head was subsequently immobilized. Two types of motor functions were performed: (1) continuous ankle dorsiflexion and (2) continuous ankle plantar flexion. Each subject performed each exercise once.
Scanning pattern: Each subject was asked to rest for 28 seconds, followed by the assigned continuous ankle dorsiflexion or continuous ankle plantar flexion movement for 20 seconds (results from preliminary experiment). fMRI scanning was performed at the same time. The resting state was free of any movements. To reduce reactions following movement, image processing only utilized original data, but the data acquired during the resting period were not used. The scanning process was as follows:

fMRI scanning parameters: GE TWINSPEED 1.5 Tesla scanner (GE Healthcare, Bethesda, MD, USA) and head coil were used with an echo-planar imaging sequence: repetition time: 2 000 ms; echo time: 40 ms; flip angle: 90°; matrix: 64 × 64 × 16; slice thickness: 5 mm; slice gap: 1.5 mm.
Data acquisition and processing
To reduce errors induced by uneven scanning at the beginning of data-acquisition period, data in the original 8 seconds of scan time were excluded. The final included data points were (1) rest for 20 seconds and (2) movement for 20 seconds. Data were exported in DICOM format. DICOM format data were transformed into SPM99 (Wellcome Dept. of Cognitive Neurology, University College London, Queen Square, London, UK) using MRIcro software wrote by Chris Rorden[26] (Neuropsychology Lab, Columbia, SC, USA). All data were realigned, normalized and smoothed using MATLAB 5.3 platform (The MathWorks, Natick, MA, USA) with SPM99 software programmed by Dick Veltman and Chloe Hutton, followed by t-test (P = 0.001) of stimulation and rest conditions. Results were displayed using a pseudo-color image.
Acknowledgments:
We thank Lin Ma and Dejun Li, Laboratory of MRI, Chinese PLA General Hospital, for technical support. The authors also thank additional staff at the Department of Neurology and Clinical Rehabilitation for their help.
Footnotes
Funding: This study was supported by the Science and Technology Innovation Nursery Foundation of Chinese PLA General Hospital, No. 09KMM41.
Conflicts of interest: None declared.
Ethical approval: This study was approved by the Ethics Committee of Chinese PLA General Hospital, China.
(Edited by Sun XJ, Liu HJ/Su LL/Song LP)
REFERENCES
- [1].Klein CS, Brooks D, Richardson D, et al. Voluntary activation failure contributes more to plantar flexor weakness than antagonist coactivation and muscle atrophy in chronic stroke survivors. J Appl Physiol. 2010;109(5):1337–1346. doi: 10.1152/japplphysiol.00804.2009. [DOI] [PubMed] [Google Scholar]
- [2].Welmer AK, Widén Holmqvist L, Sommerfeld DK. Location and severity of spasticity in the first 1-2 weeks and at 3 and 18 months after stroke. Eur J Neurol. 2010;17(5):720–725. doi: 10.1111/j.1468-1331.2009.02915.x. [DOI] [PubMed] [Google Scholar]
- [3].Miyai I, Blau AD, Reding MJ, et al. Patients with stroke confined to basal ganglia have diminished response to rehabilitation efforts. Neurology. 1997;48(1):95–101. doi: 10.1212/wnl.48.1.95. [DOI] [PubMed] [Google Scholar]
- [4].Balleine BW, O’Doherty JP. Human and rodent homologies in action control: corticostriatal determinants of goal-directed and habitual action. Neuropsychopharmacology. 2010;35(1):48–69. doi: 10.1038/npp.2009.131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Jueptner M, Weiller C. A review of differences between basal ganglia and cerebellar control of movements as revealed by functional imaging studies. Brain. 1998;121(Pt 8):1437–1449. doi: 10.1093/brain/121.8.1437. [DOI] [PubMed] [Google Scholar]
- [6].Trinastic JP, Kautz SA, McGregor K, et al. An fMRI study of the differences in brain activity during active ankle dorsiflexion and plantarflexion. Brain Imaging Behav. 2010;4(2):121–131. doi: 10.1007/s11682-010-9091-2. [DOI] [PubMed] [Google Scholar]
- [7].Ciccarelli O, Toosy AT, Marsden JF, et al. Identifying brain regions for integrative sensorimotor processing with ankle movements. Exp Brain Res. 2005;166(1):31–42. doi: 10.1007/s00221-005-2335-5. [DOI] [PubMed] [Google Scholar]
- [8].Tsao J. Ultrafast imaging: principles, pitfalls, solutions, and applications. J Magn Reson Imaging. 2010;32(2):252–266. doi: 10.1002/jmri.22239. [DOI] [PubMed] [Google Scholar]
- [9].Charlene X. Tan: Functional MRI in Neuroscience-Part I: A Review in Principal. Chin J Med Physic. 1999;16(4):209–213. [Google Scholar]
- [10].Allan HR, Martin AS. 9th ed. New York: McGraw-Hill; 2009. Adams and Victor's principles of neurology. [Google Scholar]
- [11].Adel KA, Ronald AB. New York: McGraw-Hill; 1998. Functional Neuroanatomy Text and Atlas. [Google Scholar]
- [12].Davidoff RA. The pyramidal tract. Neurology. 1990;40(2):332–339. doi: 10.1212/wnl.40.2.332. [DOI] [PubMed] [Google Scholar]
- [13].Whishaw IQ, Gorny B, Sarna J. Paw and limb use in skilled and spontaneous reaching after pyramidal tract, red nucleus and combined lesions in the rat: behavioral and anatomical dissociations. Behav Brain Res. 1998;93(1-2):167–183. doi: 10.1016/s0166-4328(97)00152-6. [DOI] [PubMed] [Google Scholar]
- [14].Muir GD, Whishaw IQ. Complete locomotor recovery following corticospinal tract lesions: measurement of ground reaction forces during overground locomotion in rats. Behav Brain Res. 1999;103(1):45–53. doi: 10.1016/s0166-4328(99)00018-2. [DOI] [PubMed] [Google Scholar]
- [15].Kombos T, Suess O, Kern BC, et al. Comparison between monopolar and bipolar electrical stimulation of the motor cortex. Acta Neurochir (Wien) 1999;141(12):1295–1301. doi: 10.1007/s007010050433. [DOI] [PubMed] [Google Scholar]
- [16].Marigold DS, Andujar JE, Lajoie K, et al. Chapter 6&motor planning of locomotor adaptations on the basis of vision: the role of the posterior parietal cortex. Prog Brain Res. 2011;188:83–100. doi: 10.1016/B978-0-444-53825-3.00011-5. [DOI] [PubMed] [Google Scholar]
- [17].Filimon F. Human cortical control of hand movements: parietofrontal networks for reaching, grasping, and pointing. Neuroscientist. 2010;16(4):388–407. doi: 10.1177/1073858410375468. [DOI] [PubMed] [Google Scholar]
- [18].Hollnagel C, Brügger M, Vallery H, et al. Brain activity during stepping: a novel MRI-compatible device. J Neurosci Methods. 2011;201(1):124–130. doi: 10.1016/j.jneumeth.2011.07.022. [DOI] [PubMed] [Google Scholar]
- [19].Kocak M, Ulmer JL, Sahin Ugurel M, et al. Motor homunculus: passive mapping in healthy volunteers by using functional MR imaging&initial results. Radiology. 2009;251(2):485–492. doi: 10.1148/radiol.2512080231. [DOI] [PubMed] [Google Scholar]
- [20].Casellato C, Ferrante S, Gandolla M, et al. Simultaneous measurements of kinematics and fMRI: compatibility assessment and case report on recovery evaluation of one stroke patient. J Neuroeng Rehabil. 2010;7:49. doi: 10.1186/1743-0003-7-49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Christensen MS, Lundbye-Jensen J, Petersen N, et al. Watching your foot move&an fMRI study of visuomotor interactions during foot movement. Cereb Cortex. 2007;17(8):1906–1917. doi: 10.1093/cercor/bhl101. [DOI] [PubMed] [Google Scholar]
- [22].Khaslavskaia S, Sinkjaer T. Motor cortex excitability following repetitive electrical stimulation of the common peroneal nerve depends on the voluntary drive. Exp Brain Res. 2005;162(4):497–502. doi: 10.1007/s00221-004-2153-1. [DOI] [PubMed] [Google Scholar]
- [23].Knash ME, Kido A, Gorassini M, et al. Electrical stimulation of the human common peroneal nerve elicits lasting facilitation of cortical motor-evoked potentials. Exp Brain Res. 2003;153(3):366–377. doi: 10.1007/s00221-003-1628-9. [DOI] [PubMed] [Google Scholar]
- [24].Francis S, Lin X, Aboushoushah S, et al. fMRI analysis of active, passive and electrically stimulated ankle dorsiflexion. Neuroimage. 2009;44(2):469–479. doi: 10.1016/j.neuroimage.2008.09.017. [DOI] [PubMed] [Google Scholar]
- [25].Phillips JP, Sullivan KJ, Burtner PA, et al. Ankle dorsiflexion fMRI in children with cerebral palsy undergoing intensive body-weight-supported treadmill training: a pilot study. Dev Med Child Neurol. 2007;49(1):39–44. doi: 10.1017/s0012162207000102.x. [DOI] [PubMed] [Google Scholar]
- [26].Rorden C, Brett M. Stereotaxic display of brain lesions. Behav Neurol. 2000;12(4):191–200. doi: 10.1155/2000/421719. [DOI] [PubMed] [Google Scholar]