
Figure 2. The structural basis for haptoglobin-haemoglobin binding by TbHpHbR.
(A) The structure of the complex between T. brucei HpHbR (blue) bound to its ligand, HpSPHb (haptoglobin is yellow, the β-subunit of haemoglobin is red and the α-subunit of haemoglobin is orange). (B) The complex viewed from the membrane proximal end, showing the contacts made by haptoglobin and the β-subunit of haemoglobin. (C) A view of the haemoglobin-binding site showing direct contacts between the haem and the receptor. Residues from the receptor that directly contact the haemoglobin subunit are shown as sticks and are numbered.

Figure 2—figure supplement 1. Stereoview of the TbHpHbR in complex with HpHb.
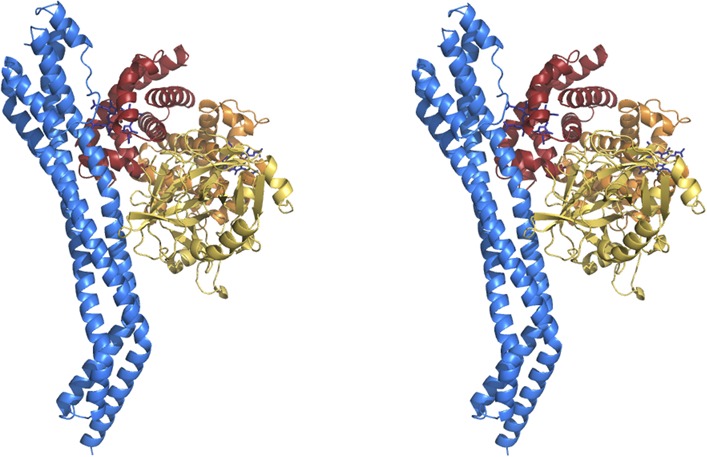
Figure 2—figure supplement 2. Small angle x-ray scattering of complexes of TcHpHbR and TbHpHbR with HpSPHb.

(A) The structure of the TbHpHbR:HpSPHb complex docked into
an ab initio molecular envelopes calculated from scattering data.
(B) The theoretical scattering calculated from ab initio
reconstructions (blue for HpSPHb, red for TbHpHbR and purple for
TbHpHbR:HpSPHb), superimposed into experimental scattering data. Guinier
plots are shown as an insert. (C) Distance distribution
functions of HpSPHb (blue), TbHpHbR (red) and TbHpHbR:HpSPHb (purple)
derived from small angle x-ray scattering. (D) A model of
the TcHpHbR:HpSPHb complex docked into an ab initio molecular envelope
calculated from scattering data. (E) The theoretical
scattering calculated from an ab initio reconstruction of the
TcHpHbR:HpSPHb complex. (F) Distance distribution function
of TcHpHbR:HpSPHb derived from small angle x-ray scattering.
Figure 2—figure supplement 3. Clashes between TbHpHbR and a haemoglobin tetramer explain why the receptor does not bind to haemoglobin.
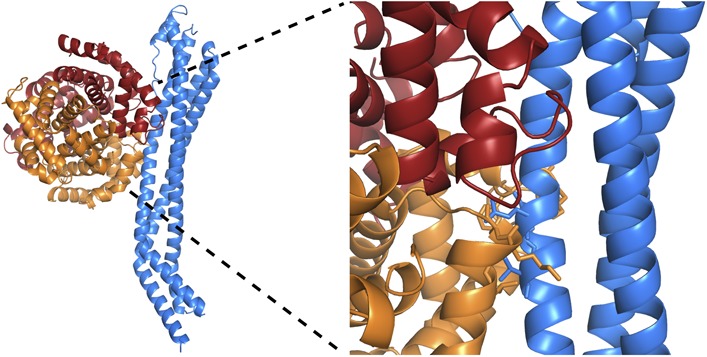
A model for a complex of TbHpHbR bound to haemoglobin. This was derived
by docking a haemoglobin tetramer onto the receptor with the
β-subunit binding to the receptor as in the TbHpHbR:HpSPHb
complex. TbHpHbR is shown in blue, the α-subunits of haemoglobin
are orange and the β-subunits are red. A close up of the model is
shown in the right hand panel with side chains involved in clashes shown
as sticks.
Figure 2—figure supplement 4. The region affected by haptoglobin cleavage is not involved in interaction with TbHpHbR.

(A) The structures of the HpSPHb region of porcine HpHb
(red) aligned to the equivalent region of human HpSPHb from the structure
of the TbHpHbR:HpSPHb complex (yellow). The structures align with a root
mean square deviation of ∼0.5 Å. The major difference is
circled and lies around the site at which haptoglobin is cleaved during a
processing event in the endoplasmic reticulum, which is disordered in the
TbHpHbR:HpSPHb complex. (B) A structural alignment of the
porcine HpSPHb structure onto the TbHpHbR:HpSPHb structure. The region
that is structurally altered by cleavage is circled and is not involved
in contacts with the receptor. This is confirmed by surface plasmon
resonance data (Figure 1—figure
supplement 1) which shows that TbHpHbR binds with similar
affinity to HpSPHb as to previously measured native, cleaved HpHb.