

Abstract
Invertebrate hosts often bring forward their reproductive effort in response to a parasitic infection. This is widely interpreted as a host-driven response aimed at compensating for the expected losses in future fitness as a result of parasitism. Here we report that mosquitoes bring forward their oviposition schedule when they are infected with Plasmodium, a parasite known to severely curtail mosquito fecundity. This response could aim at compensating for a negative time-dependent effect of the parasite on mosquito fitness, as infected mosquitoes seem to display a strong and progressive decrease in the quality of the eggs they lay. In addition, we show that this shift in oviposition date is dependent on mosquito strain: a comparison of several isogenic mosquitoes strains, one insecticide-susceptible and two insecticide-resistant ones, reveals that only the former shift their oviposition strategy when infected. This pattern suggests the existence of a costly host-driven response to parasitism, as insecticide-resistant mosquitoes have been shown to be in generally poorer condition.
Keywords: fecundity compensation, parasite manipulation, insecticide resistance, Plasmodium relictum, Culex pipiens, gonotrophic cycle
1. Introduction
A consistent observation from mosquito–Plasmodium studies is that the parasite has a strong detrimental effect on mosquito fecundity [1]. Although the proximate physiological mechanisms underlying this parasite-associated trait shift have been partially elucidated [1,2], the ultimate explanation for the reduction in the number of eggs laid by Plasmodium-infected females is still debated [2,3]. It is therefore not clear what, if any, are its consequences for the epidemiology and evolution of the disease. By contrast, the timing of egg-laying in mosquitoes is a key parameter for malaria transmission because it marks the end of a gonotrophic cycle (the sequence host-seeking, blood-feeding, oocyte maturation and egg-laying) and reinitiates the blood-seeking and feeding behaviour of the females. The duration of the gonotrophic cycle is therefore correlated with the host-biting frequency and, ultimately, with the number of hosts an infected mosquito will bite in its lifetime. If the effect of Plasmodium on the number of eggs laid by mosquitoes is well documented, to our knowledge no study has yet investigated whether the parasite can also have an effect on when those eggs are laid.
Hosts often bring forward their reproductive effort in response to a parasitic infection. This type of fecundity compensatory response has been observed when the parasite develops at the expense of the host's reproductive tissue [4] or, more generally, when the host's chances of future reproduction decrease with time since the onset of the infection [5]. In Plasmodium-infected mosquitoes, the degeneration of eggs that ultimately results in the fecundity reduction, appears to be a gradual phenomenon that increases with time after the infection [6]. Under these circumstances, laying eggs earlier may minimize the negative effect of the parasite on the mosquito's reproductive output.
Several pathogens including bacteria, viruses or fungi have already been shown to bring forward mosquito oviposition [7,8]. Here, we test whether Plasmodium-infected mosquitoes oviposit earlier than their non-infected counterparts. To do so, we use the Culex pipiens–Plasmodium relictum system, which combines a mosquito characterized by its strong flexibility regarding the timing of egg-laying [9,10] and a parasite that drastically reduces its fecundity [3]. Bringing forward the age of oviposition is expected to come at a cost, which only the fittest mosquitoes may be able to bear. To test this, we compare the timing of oviposition in infected and uninfected mosquitoes using three different isogenic C. pipiens mosquito strains [11]: an insecticide-susceptible strain and two insecticide-resistant strains that differ in their insecticide resistance mechanism [12,13]. In the absence of insecticides, these insecticide-resistant strains have been shown to incur significant fitness costs on several of their life-history traits [12].
The aims of this study are to answer the following three questions: (i) Do Plasmodium-infected mosquitoes oviposit earlier than their non-infected counterparts? (ii) Does the ability to advance the oviposition date depend on the insecticide resistance status of the mosquito? and (iii) Does egg number or egg quality (hatching rate) decrease with time to egg-laying in infected females?
2. Material and methods
Complete Material and methods is provided in the electronic supplementary material. Three isogenic C. pipiens strains were used: SLAB, susceptible to insecticides, SA4B4, resistant to insecticides through the overproduction of detoxifying esterases, and SR, resistant to insecticides through the modification of the actetylcholinesterase enzyme [12,13]. Seventy adult females from each strain were placed together in a cage to feed on an experimental canary. Ten such cages were set up, half of which were provided with a naive bird, the other half with a Plasmodium-infected bird. Engorged females were placed in tubes and provided 5 ml water for oviposition. Ten females from each infected cage were sampled and dissected on day 7 to estimate their parasitaemia (number of oocysts in the midgut) [3]. Tubes were checked daily for oviposition. Egg rafts were photographed for later counting [3]. Hatching rate was quantified as the proportion of eggs in a raft that produced a larva. Analyses were carried out using the R software. The different statistical models used are listed in the electronic supplementary material, table S2 and henceforth referred to by their number (e.g. m1, m2). Hazard ratios (HR) were estimated from Cox mixed effect models as the difference between the instantaneous risk of egg-laying between two given factor levels.
3. Results
Almost 90% of mosquitoes fed on an infected bird contained at least one oocyst (mean ± s.e.: 148 ± 40, no strain effect on oocyst prevalence m1: 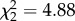 , p = 0.09, or intensity m2:
, p = 0.09, or intensity m2: 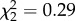 , p = 0.87). Henceforth, females exposed to an infected bird are therefore considered as being infected. Uninfected females laid on average 55.1 ± 3.9 more eggs than their Plasmodium-infected counterparts (99.5 ± 2.3 eggs), irrespective of their insecticide-resistance status. Female egg-laying date was best explained by the interaction between mosquito strain and female infection status (m3: strain × infection:
, p = 0.87). Henceforth, females exposed to an infected bird are therefore considered as being infected. Uninfected females laid on average 55.1 ± 3.9 more eggs than their Plasmodium-infected counterparts (99.5 ± 2.3 eggs), irrespective of their insecticide-resistance status. Female egg-laying date was best explained by the interaction between mosquito strain and female infection status (m3: strain × infection: 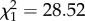 , p < 0.0001). Infected SLAB mosquitoes laid their eggs on average 1 day earlier than their uninfected counterparts (uninfected/infected: HR ± s.e = 0.52 ± 0.14; figure 1). This infection effect was, however, not observed for the insecticide-resistant strains (SA4B4: HR ± s.e = 1.12 ± 0.15; SR: HR ± s.e = 0.89 ± 0.15; figure 1). The number of eggs per raft was not dependent on the date at which they were laid (uninfected females, m4:
, p < 0.0001). Infected SLAB mosquitoes laid their eggs on average 1 day earlier than their uninfected counterparts (uninfected/infected: HR ± s.e = 0.52 ± 0.14; figure 1). This infection effect was, however, not observed for the insecticide-resistant strains (SA4B4: HR ± s.e = 1.12 ± 0.15; SR: HR ± s.e = 0.89 ± 0.15; figure 1). The number of eggs per raft was not dependent on the date at which they were laid (uninfected females, m4: 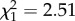 , p = 0.11; infected females, m5:
, p = 0.11; infected females, m5: 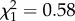 , p = 0.45). Within the rafts that produced at least one larva (92% of the total), hatching rate was found to be a decreasing function of laying date for infected females, but not for uninfected females (m6: time × infection:
, p = 0.45). Within the rafts that produced at least one larva (92% of the total), hatching rate was found to be a decreasing function of laying date for infected females, but not for uninfected females (m6: time × infection: 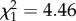 , p = 0.035; figure 2). This interaction was not observed when analysing each mosquito strain separately (SLAB, m7:
, p = 0.035; figure 2). This interaction was not observed when analysing each mosquito strain separately (SLAB, m7: 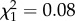 , p = 0.78; SA4B4, m8:
, p = 0.78; SA4B4, m8: 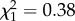 , p = 0.54; SR, m9:
, p = 0.54; SR, m9: 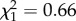 , p = 0.42; see the electronic supplementary material, figure S1).
, p = 0.42; see the electronic supplementary material, figure S1).
Figure 1.

Time to egg-laying: Kaplan–Meier curve of the time it takes the susceptible strain (a) SLAB, and the insecticide-resistant strains (b) SA4B4 and (c) SR to oviposit. Solid line: Plasmodium-infected mosquitoes, dashed line: uninfected mosquitoes. (d) Mean ± s.e. of the median time to egg-laying (i.e. the time at which 50% of the females oviposited) for each strain and treatment (empty circles, uninfected females; grey circles, infected females).
Figure 2.

Linear regression of hatching rate against egg-laying date. Symbol size is proportional to the number of observations used to calculate the mean hatching rate for uninfected mosquitoes (empty circles) or infected females (filled circles). Bars represent the s.e. of the mean. Dashed line: uninfected females, solid line: infected females. A logistic transformation was applied to hatching rate before plotting.
4. Discussion
We show that (i) mosquitoes lay their eggs up to 2 days earlier when they are infected with malaria parasites, (ii) this effect was only found in insecticide-susceptible mosquitoes, and (iii) late oviposition seems to have a negative effect on egg hatching rate in infected females but not in uninfected ones (see figure 2; electronic supplementary material, figure S1).
There are three potential explanations for these results. First, early oviposition in infected females may simply be a by-product of their lower fecundity [3,7]. Several lines of evidence make this option unlikely. Oocyte development is synchronous in the hundreds of ovarioles that constitute the females' ovaries [14] so producing a smaller egg batch might not necessarily be faster. In addition, under this scenario we would have expected to find a positive correlation between egg-laying date and egg number, and no differences in egg-laying timing between insecticide-resistant and -susceptible mosquitoes since they both suffer a similar reduction in fecundity when infected [3]. Neither of these conditions was, however, met. Second, early oviposition could be a parasite-driven strategy. Plasmodium has indeed already been shown to be capable of manipulating other aspects of mosquito behaviour to its own advantage [15]. Shifting the oviposition schedule to an earlier date could benefit the parasite by speeding up the return to the host-seeking and blood-feeding behaviour of its vector, thereby bringing forward the next transmission event. Against this argument is the fact that the oviposition shift is observed early in the infection, when the mosquitoes still carry non-transmissible (oocyst) stages of the parasite. In addition, oocyst-infected mosquitoes have been shown to display a downregulation in their blood-feeding behaviour as a result of which they are less likely to host-seek during this phase of the infection [16,17]. On the other hand, a second, uninfected, blood meal may enhance the developmental potential of the parasite within mosquitoes, increasing the number of sporozoites produced [18]. For the parasite, the benefits of producing more sporozoites may, however, be offset by the mortality risks for their vectors of feeding on a host when the parasite is not readily transmissible.
The final scenario is that the observed shift in egg-laying date is yet another example of a host-driven compensatory response to the infection [5]. An increase in early reproductive investment is expected if parasitism reduces survival and, by extension, the host's future reproductive chances [19]. However, in contrast to other systems [20], P. relictum increases C. pipiens longevity [3]. Alternatively, such compensatory responses may be expected if the parasite reduces host fecundity [21]. Indeed, Plasmodium significantly reduces mosquito fecundity in all Plasmodium–vector associations tested to date [3]. Here, we show that egg quality, measured as the proportion of eggs that successfully hatch as larvae, decreases significantly with oviposition time such that egg rafts laid by an infected mosquito on day 12 contain on average 36% fewer viable larvae than eggs laid on day 1 (figure 2). There may thus be a straightforward advantage to laying their eggs earlier for infected mosquitoes. This gradual egg degeneration may be made possible by the fact that, contrary to what happens in Anopheles, Culex eggs remain permeable to nutrients for weeks, long after chorionation is over [9]. The results of this reduction in egg hatching rate with time should, however, be cautiously interpreted as they were not maintained when the strains were analysed separately (electronic supplementary material, figure S1, m7–m9). Further work is therefore needed before it can be conclusively established whether Plasmodium infection reduces egg quality and whether this reduction is progressive with time.
Despite having a similar parasitaemia and fecundity reduction [3,13], we found that only insecticide-susceptible mosquitoes advanced their egg-laying date. This suggests that bringing reproduction forward comes at cost, which only insecticide-susceptible mosquitoes can afford. Increased early reproductive effort is known to come with a suite of life-history costs [4,22]. We have previously shown that in Plasmodium-infected C. pipiens mosquitoes an increase in reproductive effort (fecundity) comes with substantial longevity costs. As expected, the slope of the fecundity/longevity trade-off is significantly steeper for insecticide-resistant mosquitoes [3]. Such a cost may add an unbearable constraint on the insecticide-resistant mosquitoes’ ability to adopt a plastic egg-laying response. Pending confirmation using other genetic backgrounds (i.e. other mosquito strains or field-caught insecticide-susceptible and -resistant mosquitoes), these results might have important consequences for the evolution and maintenance of insecticide resistance in the field.
In conclusion, to our knowledge this is the first time that an accelerated onset of oviposition has been reported in Plasmodium-infected mosquitoes. Although there are several potential explanations for these results, on balance the most likely scenario is the existence of a host-driven compensatory response aimed at minimizing the negative effect of Plasmodium parasitism on mosquito fecundity [3]. Pending confirmation that early oviposition maps directly onto a shorter gonotrophic cycle, these findings could have consequences for the epidemiology and evolution of malaria transmission in the field.
Supplementary Material
Supplementary Material
Supplementary Material
Acknowledgements
We thank F. Gatchitch, P. Perret, M. Weill and F. Zélé for their help.
Ethics statement
Animal experiments were carried out in strict accordance with the ‘National Charter on the Ethics of Animal Experimentation’ of the French Government and all efforts were made to minimize suffering. Experiments were approved by the Ethical Committee for Animal Experimentation established by the authors' institution (CNRS) under the auspices of the French Ministry of Education and Research (permit no. CEEA-LR-1051).
Data accessibility
Data available from the Dryad Digital Repository: http://doi.org/10.5061/dryad.q8292.
Author contributions
J.V., A.R. and S.G. conceived and designed the experiments. J.V., A.R. and A.N. performed the experiments. J.V. carried out the statistical analyses. J.V., A.R. and S.G. wrote the paper. All authors read and approved the manuscript.
Funding statement
J.V. was funded through an FCT (GABBA) grant, S.G. by ERC Starting Grant EVOLEPID243054 and A.R. by an ANRSEST grant.
References
- 1.Hurd H. 2003. Manipulation of medically important insect vectors by their parasites. Annu. Rev. Entomol. 48, 141–161. ( 10.1146/annurev.ento.48.091801.112722) [DOI] [PubMed] [Google Scholar]
- 2.Hurd H. 2009. Evolutionary drivers of parasite-induced changes in insect life-history traits: from theory to underlying mechanisms. Adv. Parasitol. 68, 85–110. ( 10.1016/s0065-308x(08)00604-0) [DOI] [PubMed] [Google Scholar]
- 3.Vézilier J, Nicot A, Gandon S, Rivero A. 2012. Plasmodium infection decreases fecundity and increases survival of mosquitoes. Proc. R. Soc. B 279, 4033–4041. ( 10.1098/rspb.2012.1394) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Minchella DJ, Loverde PT. 1981. A cost of increased early reproductive effort in the snail Biomphalaria glabrata. Am. Nat. 118, 876–881. ( 10.1086/283879) [DOI] [Google Scholar]
- 5.Agnew P, Koella JC, Michalakis Y. 2000. Host life history responses to parasitism. Microb. Infect. 2, 891–896. ( 10.1016/S1286-4579(00)00389-0) [DOI] [PubMed] [Google Scholar]
- 6.Hopwood JA, Ahmed AM, Polwart A, Williams GT, Hurd H. 2001. Malaria-induced apoptosis in mosquito ovaries: a mechanism to control vector egg production. J. Exp. Biol. 204, 2773–2780. [DOI] [PubMed] [Google Scholar]
- 7.Garza-Hernandez JA, Rodriguez-Perez MA, Isabel Salazar M, Russell TL, Adeleke MA, de Luna-Santillana EDJ, Reyes-Villanueva F. 2013. Vectorial capacity of Aedes aegypti for dengue virus type 2 is reduced with co-infection of Metarhizium anisopliae. PLoS Negl. Trop. Dis. 7, e2013 ( 10.1371/journal.pntd.0002013) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.de Almeida F, Moura AS, Cardoso AF, Winter CE, Tania Bijovsky A, Suesdek L. 2011. Effects of Wolbachia on fitness of Culex quinquefasciatus (Diptera; Culicidae). Infect. Genet. Evol. 11, 2138–2143. ( 10.1016/j.meegid.2011.08.022) [DOI] [PubMed] [Google Scholar]
- 9.Van Handel E. 1992. Postvitellogenic metabolism of the mosquito (Culex quinquefasciatus) ovary. J. Insect Physiol. 38, 75–79. ( 10.1016/0022-1910(92)90023-7) [DOI] [Google Scholar]
- 10.Dieter KL, Huestis DL, Lehmann T. 2012. The effects of oviposition-site deprivation on Anopheles gambiae reproduction. Parasit. Vectors 5, 235 ( 10.1186/1756-3305-5-235) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Berticat C, Boquien G, Raymond M, Chevillon C. 2002. Insecticide resistance genes induce a mating competition cost in Culex pipiens mosquitoes. Genet. Res. 79, 41–47. ( 10.1017/S001667230100547X) [DOI] [PubMed] [Google Scholar]
- 12.Vézilier J, Nicot A, De Lorgeril J, Gandon S, Rivero A. 2013. The impact of insecticide resistance on Culex pipiens immunity. Evol. Appl. 6, 497–509. ( 10.1111/eva.12037) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Vézilier J, Nicot A, Gandon S, Rivero A. 2010. Insecticide resistance and malaria transmission: infection rate and oocyst burden in Culex pipiens mosquitoes infected with Plasmodium relictum. Malar. J. 9, 379 ( 10.1186/1475-2875-9-379) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Clements AN. 2000. The biology of mosquitoes. Wallingford, UK: CABI Publishing. [Google Scholar]
- 15.Cator LJ, Lynch PA, Thomas MB, Read AF. 2014. Alterations in mosquito behaviour by malaria parasites: potential impact on force of infection. Malar. J. 13, 164 ( 10.1186/1475-2875-13-164) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cator LJ, George J, Blanford S, Murdock CC, Baker TC, Read AF, Thomas MB. 2013. ‘Manipulation’ without the parasite: altered feeding behaviour of mosquitoes is not dependent on infection with malaria parasites. Proc. R. Soc. B 280, 20130711 ( 10.1098/rspb.2013.0711) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Charlwood JD, Tomas EVE. 2011. Do developing malaria parasites manipulate their mosquito host? Evidence from infected Anopheles funestus (Giles) from Mozambique. Trans. R. Soc. Trop. Med. Hyg. 105, 352–354. ( 10.1016/j.trstmh.2011.02.006) [DOI] [PubMed] [Google Scholar]
- 18.Ponnudurai T, Lensen AHW, Vangemert GJA, Bensink MPE, Bolmer M, Meuwissen J. 1989. Sporozoite load of mosquitos infected with Plasmodium falciparum. Trans. R. Soc. Trop. Med. Hyg. 83, 67–70. ( 10.1016/0035-9203(89)90708-6) [DOI] [PubMed] [Google Scholar]
- 19.Gandon S, Agnew P, Michalakis Y. 2002. Coevolution between parasite virulence and host life-history traits. Am. Nat. 160, 374–388. ( 10.1086/341525) [DOI] [PubMed] [Google Scholar]
- 20.Ferguson HM, Read AF. 2002. Why is the effect of malaria parasites on mosquito survival still unresolved? Trends Parasitol. 18, 256–261. ( 10.1016/S1471-4922(02)02281-X) [DOI] [PubMed] [Google Scholar]
- 21.Bonds MH. 2006. Host life-history strategy explains pathogen-induced sterility. Am. Nat. 168, 281–293. ( 10.1086/506922) [DOI] [PubMed] [Google Scholar]
- 22.Rono MK, Whitten MMA, Oulad-Abdelghani M, Levashina EA, Marois E. 2010. The major yolk protein vitellogenin interferes with the anti-Plasmodium response in the malaria mosquito Anopheles gambiae. PLoS Biol. 8, e1000434 ( 10.1371/journal.pbio.1000434) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data available from the Dryad Digital Repository: http://doi.org/10.5061/dryad.q8292.