
Figure 1.
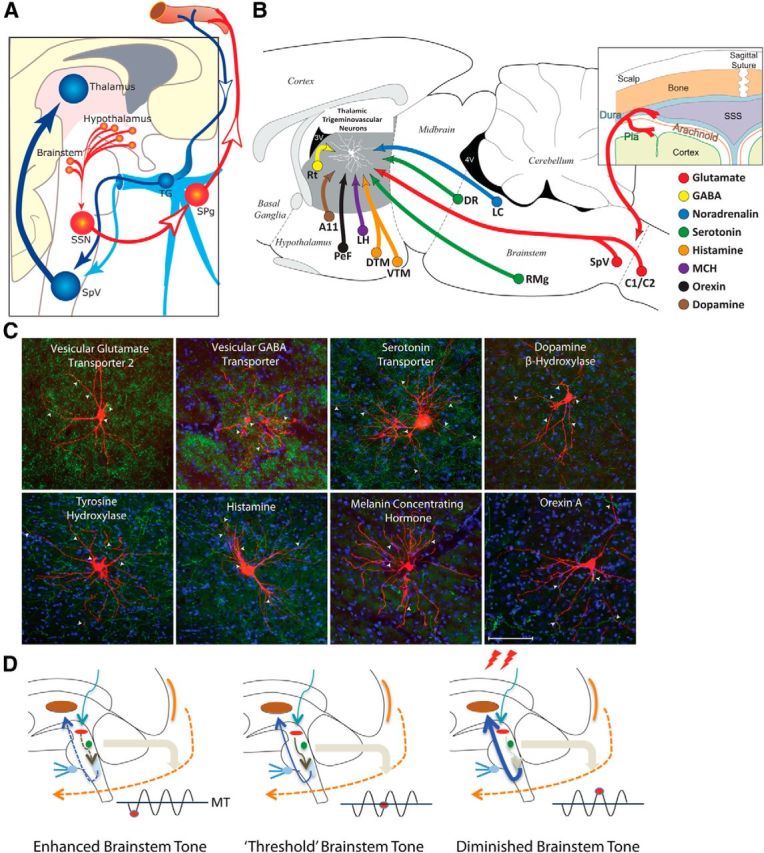
From prodromes to headache: proposed hypothesis for the initiation of headache by the hypothalamus and brainstem. A, Hypothalamic–parasympathetic pathway for the activation of meningeal nociceptors by neurons that regulate homeostasis, circadian rhythms, and autonomic functions (adapted from Burstein and Jakubowski, 2005). Hypothalamically mediated activation of preganglionic parasympathetic neurons in the SSN can trigger the release of acetylcholine, vasoactive intestinal peptide, and nitric oxide from meningeal terminals of postganglionic parasympathetic neurons in the SPG. B, C, Neurochemical pathways capable of modulating the excitability of relay thalamocortical neurons in response to deviation from physiological (food intake, sleep) and emotional (stress, anxiety) homeostasis. The illustration (top right) shows the hypothalamic and brainstem origin of each of the pathways found to converge on thalamic trigeminovascular neurons (adapted from Kagan et al., 2013; Noseda et al., 2014). The photomicrographs show the extent of innervation by vesicular glutamate transporter, vesicular GABA transporter, serotonin transporter, dopamine beta hydroxylase, tyrosine hydroxylase, histamine, melanin-concentrating hormone, and orexin A. D, Conceptual illustration of how brainstem tone (allostatic load) may allow the headache to develop incosistently in response to identical changes in external and internal conditions. Brainstem “state of tone” can limit afferent nociceptive drive in migraine-susceptible individuals (adapted from Borsook and Burstein, 2012). Fluctuation of activity in brainstem neurons is thought to drive adaptive behavior. In the context of migraine, this can apply to the modulation of nociceptive signals from the meninges. The gating of these signals depends on the threshold of the neural networks that modify these afferent signals. Thus, the robustness of the “gate” that allows nociceptive signals to drive central trigeminovascular neurons (and thus headache) is dictated by brainstem tone. When the brainstem tone is high [red dot below line of migraine threshold (MT)], nociceptive signals are inhibited; and when the brainstem tone is low (red dot above MT), afferent signals are not effectively blocked. The model illustrates the following three functional brainstem states: (1) normal state, when cyclical brainstem activity is high, the potency of pain facilitation (enhanced synaptic strength in the dorsal horn) is too high to allow nociceptive signals from the periphery to drive the central neurons into the active state (left); (2) threshold state, at threshold, the system has reached a primed state that could tip into a functional state that would allow nociceptive drive from the dura to activate the central trigeminovascular neurons (middle); and (3) migraine state, when cyclical brainstem activity is low (more sensitive to stimuli), nociceptive signals from the periphery can drive the central neurons into the active state (right). A11, Hypothalamic dopaminergic nucleus; C1/C2, cervical spinal cord segments; DR, dorsal raphe nucleus; DTM, dorsal tuberomammary hypothalamic nucleus; LC, locus ceruleus; LH, lateral hypothalamus, PeF, perifornical area; RMg, nucleus raphe magnus; TG, trigeminal ganglion.