
Figure 9.
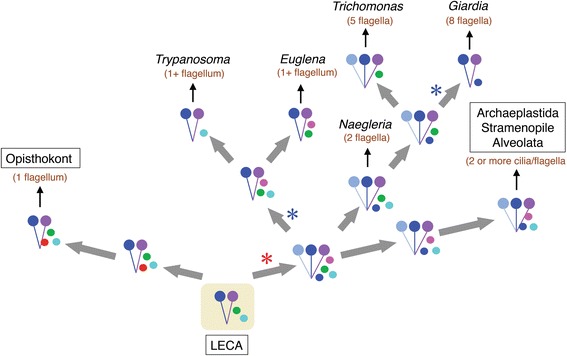
A possible model for the evolution of, and diversification in, the structures of outer arm dynein and corresponding Ca 2+ sensors during eukaryotic evolution. The model is based on analyses of the structures of outer arm dynein (two-headed, three-headed) and the types of Ca2+-sensor in each group of eukaryotes. It is assumed that the heavy chains and Ca2+-sensors of outer arm dynein of the last eukaryotic common ancestor (LECA) preceded duplication, and that duplication and divergence of Ca2+-sensors occurred at an early stage of eukaryotic diversification. The model is arranged so that the positions of eukaryotic groups match with widely accepted phylogenetic relationships [128,158]. The number of cilia/flagella per cell is also indicated in parenthesis (brown letters). Note that the numbers of cilia/flagella in Euglena and Trypanosoma are indicated as ‘1+,’ since these organisms are considered to have been biflagellates but lost or largely degenerated one of the two flagella during evolution. In this model, duplication of dynein heavy chain occurred at the root of the bikont lineage. Duplication and divergence of Ca2+-sensors would have already occurred in the ancestral organisms that contained three-headed dynein. An ancestral organism containing three-headed dynein might have recruited LC4-like sensors or CNB/NCS-like sensors and then branched into the Metamonadan (Trichomonas + Giardia) and Discoban lineages. Loss of dynein heavy chains would have occurred in Giardia and the Euglenozoa. Red or blue asterisks represent duplication or loss of a dynein heavy chain, respectively. Colored dots next to the two- or three-headed dyneins represent Ca2+-sensors (red, calaxin; blue, LC4; magenta, DC3; green, NCS; cyan, CNB). In the lineage of opisthokonts or Archaeplastida/Stramenopile/Alveolata, calaxin, LC4 or DC3 is demonstrated to be bound to the dynein heavy chain, although it is not known whether Ca2+-sensors in Excavates or any of the hypothetical ancestors could bind to the dynein or not.