
Figure 7.
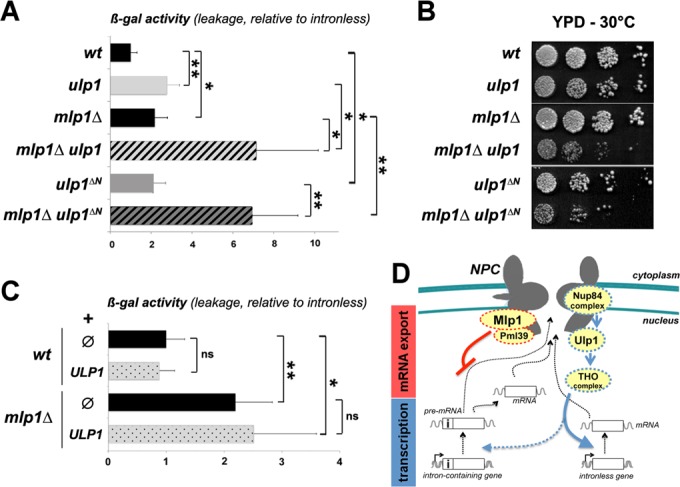
Distinct functions of nuclear pore-associated proteins in intron-containing gene expression. (A) β-gal activities (mean ± SD; n = 5) were measured in the indicated strains transformed with the pre-mRNA ‘leakage’ reporter. β-gal activities relative to the ones obtained with strains transformed with the intronless reporter are represented. Values were set to 1 for wt cells. (B) Serial dilutions of the indicated strains were grown at 30°C on YPD solid medium. (C) β-gal activities (mean ± SD; n = 5) were measured in the indicated strains transformed with the pre-mRNA ‘leakage’ reporter together with the ULP1-overexpressing construct (pRS315-NOP1-GFP-ULP1, ‘ULP1’) or an empty vector (pRS315, ‘ø’). β-gal activities relative to the ones obtained with strains transformed with the intronless reporter are represented. Values were set to 1 for wt cells without ULP1. Note that the ULP1 construct used here was previously reported to suppress the phenotypes associated with Ulp1 loss-of-function (49,50). Raw data are provided in Supplementary Table S4. (D) Mlp1/Pml39 and Nup84/Ulp1/THO impact on distinct stages of intron-containing mRNA biogenesis and export. On the one hand, the Nup84 complex contributes to the NPC localization of Ulp1, which targets the THO complex for desumoylation. Alteration in any of these components would impinge on THO complex activity, which is more dispensable for the transcription of intron-containing as compared to intronless genes. On the other hand, Mlp1/Pml39 have specific roles in nuclear retention of intron-containing pre-mRNAs (see Discussion). Note that nuclear pores also influence gene expression at the stage of transcription initiation, as shown for some inducible genes (40).