

Abstract
In orthopedics, tissue engineering approach using stem cells is a valid line of treatment for patients with bone defects. In this context, mesenchymal stromal cells of various origins have been extensively studied and continue to be a matter of debate. Although mesenchymal stromal cells from bone marrow are already clinically applied, recent evidence suggests that one may use mesenchymal stromal cells from extra-embryonic tissues, such as amniotic fluid, as an innovative and advantageous resource for bone regeneration. The use of cells from amniotic fluid does not raise ethical problems and provides a sufficient number of cells without invasive procedures. Furthermore, they do not develop into teratomas when transplanted, a consequence observed with pluripotent stem cells. In addition, their multipotent differentiation ability, low immunogenicity, and anti-inflammatory properties make them ideal candidates for bone regenerative medicine. We here present an overview of the features of amniotic fluid mesenchymal stromal cells and their potential in the osteogenic differentiation process. We have examined the papers actually available on this regard, with particular interest in the strategies applied to improve in vitro osteogenesis. Importantly, a detailed understanding of the behavior of amniotic fluid mesenchymal stromal cells and their osteogenic ability is desirable considering a feasible application in bone regenerative medicine.
Keywords: Mesenchymal stromal cells, Amniotic fluid, Amniotic fluid mesenchymal stromal cells, Amniotic fluid stem cells, Osteogenesis, Bone regeneration
Core tip: Several papers regarding the osteogenic differentiation potential of cells isolated from amniotic fluid have been published so far with particular attention to various feasible approaches to improving differentiation both in vitro and in vivo. Hence, an overview is necessary on the data reported up to now in order to understand the potential of amniotic fluid-derived cells in bone regenerative medicine. This review takes a general look at the current state-of-the-art of the osteogenic ability of amniotic fluid-derived cells and the different strategies available to improve bone regeneration.
INTRODUCTION
In the last few decades degenerative lesions of the musculoskeletal system have become increasingly common, leading to social and economic problems, and intensifying the demand on medical services[1]. Common examples are temporary and permanent disability due to osteoarthritis or fractures caused by loss of bone mass such as osteoporosis or osteopenia, particularly in the elderly. In the younger population widespread active lifestyles including sport activities have increased the possibility of bone and cartilage damage. Moreover, bone defects mostly emerge from trauma, tumor resection and congenital malformation. So far, the most effective clinical method for the reconstruction of large bone defects is the use of autogenous bone grafts, commonly harvested from the posterior iliac crest and transplanted into a local bone defect[2]. The weakness of this method is the surgical stress the patient undergoes in extracting bone with possible subsequent inflammation and donor site morbidity. Furthermore, the quantity of extractable bone is limited.
Recently, advances in the field of stem cells have come to represent a valid alternative to this method[3]. Nowadays there is growing interest in therapies based on mesenchymal stromal cells (MSCs) as a potential effective treatment for bone defects. MSCs are multipotent cells with the potential to engender a range of specialized cell types, such as osteoblasts, chondrocytes and adipocytes[4]. The mesenchymal progenitor cells in the bone marrow are able to differentiate into osteoblasts following the influence of multiple osteogenic signals[5,6]. In particular, after fracture MSCs of bone marrow are transferred to the site of bone injury through peripheral blood, thus enhancing the healing potential of local MSCs[7]. In this condition, bone morphogenic proteins (BMPs) play an important role. Indeed, osteoblastic differentiation begins when the BMPs bind their receptors activating the transcription factors runt-related transcription factor 2 (Runx2) and Osterix, and subsequent downstream osteoblast specific genes, through activation of a Wnt/LRP5 cascade, which is crucial in bone mass modeling[5,8,9].
Regarding the principal signaling pathways involved in MSC differentiation, various evidences have suggested that during cultivation in vitro, an inverse relationship exists between the commitment of MSCs toward osteogenic and adipogenic lineages[10]. Several cell signaling cascades are involved in this cell fate decision between osteo- and adipogenesis[11]. The master regulator of osteogenesis is Runx2, the gene target of many signaling pathways, including but not limited to transforming growth factor-beta 1 (TGF-β1), BMP[12], Wingless type (Wnt)[13], Hedgehog (HH)[14], and (NEL)-like protein type 1 (NELL-1)[15].
Thus, considering the crucial role of MSCs in bone healing, the strategy of using the osteogenic potential of such cells transplanted into the bone defect seems promising[7,16].
Mesenchymal stromal cells derived from bone marrow (BM-MSCs) have been used in clinical trials for the treatment of bone defects[17]. However, bone marrow aspiration is a difficult, invasive and painful procedure for the donor. The amount of BM-MSCs is usually between 0.001% and 0.01% of the total population and, in addition, the BM-MSC number and differentiation potential may be affected by increasing donor age. This implies an extensive in vitro expansion of such cells before transplantation, thus increasing the risk of possible differentiation induction and epigenetic modifications[18]. On the other hand, the use of allogeneic BM-MSCs for bone repair is unsuitable due to immune rejection[19].
As an alternative, mesenchymal stromal cells can be found in various adult organs and tissues[20], including blood[21], adipose tissue[22] and dermis[23]. Although sharing similar properties, these MSCs from various sources have different gene expression profile and differentiation ability[24].
Mesenchymal stem-like cells have also been recognized in fetal blood, liver and bone marrow[25]. In this regard, Guillot and colleagues have shown that first trimester fetal blood, liver, and bone marrow MSCs possess greater osteogenic differentiation potential than adult BM-MSCs[26]. They found through quantitative real-time RT-PCR that 16 osteogenic specific genes (OC, ON, BSP, OP, Col1, PCE, Met2A OPG, PHOS1, SORT, ALP, BMP2, CBFA1, OSX, NOG, IGFII) were more expressed in fetal MSCs under basal conditions and were up-regulated during osteogenic differentiation both in vitro and in vivo[26]. These cells are more primitive than adult MSCs[25], therefore potentially promising for therapeutic use in regenerative medicine, even if isolating them is subject to considerable public unease.
An alternative approach could result from the use of MSCs derived from extra-embryonic tissues, which possess the advantage of being isolated from tissues normally discarded after birth, hence exempt from ethical concern, such as amniotic fluid, umbilical cord and placenta[27-29].
This review will focus on the biological properties of MSCs isolated from amniotic fluid (AF-MSCs) with particular attention to their in vitro osteogenic differentiation potential with a view to possible final application in bone regenerative medicine (Figure 1). Of note, these cells possess a greater proliferative capacity, lower immunological reactivity and lower risk of graft-versus-host disease than those derived from adult bone marrow[30]. Importantly, compared with other MSC sources, such as umbilical cord blood-derived MSCs, AF-MSCs are more easily isolated and show better proliferation ability[31].
Figure 1.
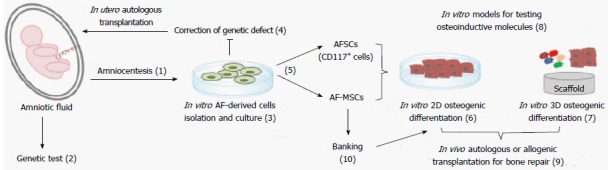
In vitro and in vivo application of amniotic fluid-derived cells. Amniotic fluid-derived cells isolated from amniotic fluid samples obtained following ultrasound-guided amniocentesis (1) for genetic test (2), could be in vitro expanded (3). These cells may be genetically manipulated and transplanted back into the same fetus (4). AF-derived cells can be used unselected (AF-MSCs) or selected for CD117 (AFSCs) (5). Both cellular populations can be in vitro differentiated in 2D (6) or 3D (7) culture conditions. They represent an ideal in vitro model for testing osteoinductive molecules (8) and for in vivo autologous or allogeneic transplantation (9). Alternatively, they could also be banked (10) and adopted post-thawing.
In particular, AF-MSCs are autologous to the foetus representing an attractive source for the treatment of perinatal disorders such as congenital malformations[32,33]. Hence, affected children could benefit from their own cells which could be banked, expanded in culture or properly engineered and implanted in the neonatal period. In addition, AF-MSCs are semi-allogeneic to each parent, therefore potentially useful for the other members of the family[34].
Moreover, the beneficial effect of AF-MSCs observed in preclinical studies, such as lung injury[35], ischemic heart[36], acute bladder injury[37], neovascularization[38], encourages their future application in regenerative medicine tissue engineering.
AMNIOTIC FLUID-DERIVED CELLS
Human amniotic fluid, contained within the amniotic cavity, begins to appear at the second week of gestation. It surrounds the growing fetus, protects from outside injury and acts as the vehicle with the mother[30]. It contains a variety of cells arising from all three germ layers (ectoderm, mesoderm and endoderm) routinely obtained for prenatal diagnosis of fetal abnormalities during second trimester pregnancy, through a minimally invasive technique. Amniotic fluid cellular composition substantially depends on the gestational age[27]. Most of the cells present in the amniotic fluid are terminally differentiated and have low proliferative capacity[32]. Three major cellular types can be classified based on morphological, biochemical and growth characteristics: epithelioid, amniotic fluid and fibroblastic cells[39].
Interestingly, a population that expresses the surface antigen c-kit (CD117), stem cell factor receptor, can be isolated from amniotic fluid. These cells, named amniotic fluid stem cells (AFSCs), represent about 1% of the total cells[40]. It should be noted that, mesenchymal stromal cells (AF-MSCs) with a multilineage differentiation potential are present in the amniotic fluid[41,42].
AF-MSCs are highly proliferative with a normal karyotype after long-term in vitro culture and do not form teratomas when transplanted in vivo[43]. They do not display any hematopoietic molecular markers (CD14, CD31 and CD45), while they express the intracellular stemness markers TERT, SOX2 and Nanog and the surface adhesion molecules CD29, CD58, CD166 and CD90[44].
The absence of HLA-DR and presence of HLA-ABC suggest that these cells may be applicable in immune-mediated disorders as well as in the treatment of graft-versus-host disease[43]. It is also important that AF-MSCs display a broad differentiation potential toward multiple lineages (i.e., adipogenic, chondrogenic, myogenic and osteogenic)[34]. They possess an intermediate differentiation potential between embryonic (pluripotent) and adult cells with advantages over both. Compared to adult cells, AF-MSCs possess greater differentiation potential and more primitive properties with fewer accumulated mutations[45]. Respect to embryonic stem cells (ESCs), AF-MSCs possess the advantage to do not form teratomas when transplanted in vivo. Indeed, although ESCs are pluripotent cells which maintain high plasticity and extensive self-renewal capacity, possible host immune rejection after allotransplantation and the formation of tumors when injected undifferentiated or partially differentiated in vivo, raise safety concerns[46-48]. Furthermore, their clinical use is limited by ethical issues due to the need to isolate them from the inner cell mass of a blastocyst[49].
In the last few years methods able to generate patient-specific pluripotent stem cells (iPSCs) from adult cells have been developed to overcome the limitations associated with ESCs[50,51]. Such iPSCs, obtained through the ectopic expression of defined transcription factors (Oct4, Sox2, Klf-4, c-Myc), are molecularly and functionally similar to ESCs. They show similar morphology and growth properties, express pluripotency markers, are able to generate germline-competent chimeras and form tumors when injected into immune-compromised mice[51]. Although iPSCs allow one to overcome both the ethical and the tissue compatibility problems of ESCs, current studies are still investigating the safety profile of these cells for therapeutic application[52]. The main limitation related to iPSC generation is the use of retroviruses or lentiviruses, which could cause mutagenesis leading to a risk for teratogenesis.
Therefore, given the easier accessibility and the faster availability of a great number of AF-MSCs in culture in comparison to iPSCs, AF-MSCs may hold great promise in regenerative medicine.
However, since iPSCs seem one of the most promising future sources of stem cells for tissue regeneration, research is going ahead to find the best cell source to reprogram and develop alternative methods for generating pluripotent cells using non integrating systems. In this context, recent evidences has suggested that cells derived from amniotic fluid are more easily and rapidly reprogrammed than adult cells[53-55]. More recently, iPSCs have been developed from amniotic fluid cells without ectopic factors by culture in an appropriate medium[56], thus reinforcing their potential application in cellular replacement therapies.
These important features, together with the high proliferation rate, ease of retrieval and more stable profile, provide a convincing proof-of-principle for potential autologous application of AF-MSCs for bone regeneration in perinatal applications[57]. Furthermore, a bank of such cells is achievable and may in the future provide a plentiful source for autologous therapy in adulthood as well as for transplantation into HLA-matched recipients.
IN VITRO OSTEOGENIC DIFFERENTIATION
The osteogenic differentiation capacity of AF-derived cells obtained from various sources (human, sheep, mouse and rat) has been broadly documented[39,40,58,59]. Some studies have reported the great potential of c-kit selected amniotic fluid cells prior to osteogenic differentiation[40], while others have demonstrated that unselected cells are also able to properly differentiate into osteo-specific cells[13,41,44,60]. In this review we mainly discuss the in vitro osteogenic differentiation of AF-MSCs with occasional mention of papers in which authors use c-kit selected cells, named AFSCs (Table 1). It should be pointed out that, compared to BM-MSCs, AF-MSCs have higher self-renewal capacity and are more potent for lineage-specific differentiation[40]. Therefore, considering also their more easily isolation and the great number of available cells, AF-MSCs may represent excellent candidates for cell replacement therapies[61]. Of note, even if there are other MSC sources easily available, like adipose-derived MSCs, the osteogenic capacity of such cells compared to BM-MSCs is now still debated[62].
Table 1.
Studies describing in vitro modification of standard culture condition to induce osteogenic differentiation of AF-derived cells
| Cell source | Methods to induce osteogenic differentiation | Ref. |
| hAF-MSCs | Culture in standard osteogenic medium on sandblasted and acid etching titanium (SLA titanium) | [60] |
| hAF-MSCs | Transfection with a defective adenoviral vector expressing human LMP3 | [63] |
| hAFSCs | Standard osteogenic medium plus curculigoside | [65] |
| hAFSCs | Standard osteogenic medium plus naringin | [66] |
| hAFSCs | 100 µmol/L 2P-ascorbic acid, 100 nmol/L dexamethasone, supplemented with different concentrations of Ferutinin | [67] |
| hAF-MSCs | Induction of miR-21 | [71] |
| hAFSCs | Medium containing 50 nmol/L rhBMP-7, 50 mg/mL ascorbic acid, and 10 mmol/L b-glycerophosphate on nanofibrous or solid walled scaffolds | [86] |
| hAFSCs | 10 nmol/L dexamethasone, 6 mmol/L β-glycerol phosphate, 50 mg/mL ascorbic acid 2-phosphate, 50 ng/mL L-thyroxine on electrospun nanofiber meshes | [88] |
| hAFSCs | Standard osteogenic medium on: (1) Fibroin scaffold; (2) Collagen scaffold; and (3) Poly-D,L-lactic acid scaffold; | [89] |
| hAFSCs | Standard osteogenic medium on microfibrous starch and poly(ε-caprolactone) scaffold | [90] |
| hAFSCs | Medical-grade poly-ε-caprolactone scaffold | [91] |
| hAFSCs | Standard osteogenic medium on collagen matrix derived from porcine bladder submucosa matrix and poly (lactide-co-glycolide) | [92] |
| hAFSCs | Standard osteogenic medium on construct composed of collagen type I | [93] |
| hAFSCs | α-MEM plus 17% FBS, 1 μmol/L dexamethasone, 6 mmol/L of β-glycerol phosphate, 50 μg/mL ascorbic acid 2-phosphate, and 50 ng/mL thyroxine on biomaterial named Skelite | [38] |
MSCs: Mesenchymal stromal cells; AF-MSCs: MSCs isolated from amniotic fluid; AFSCs: Amniotic fluid stem cells; LMP3: Lim mineralization protein 3.
The conventional protocol applied to induce osteogenic differentiation of AF-MSCs consists of basal medium, such as Dulbecco’s modified Eagle’s medium, supplemented with 10% Fetal Bovine Serum, β-glycerolphosphate, ascorbic acid and dexamethasone (standard osteogenic medium). For the purpose of assessing in vitro differentiation, protein and gene expression of specific osteogenic markers [e.g., alkaline phosphates (ALP), Collagen type I, bone sialoprotein, osteocalcin, RUNX2] are usually evaluated in addition to specific colorimetric assays, such as ALP, Alizarin Red S and Von Kossa staining.
However, various approaches have been applied to improve and accelerate differentiation of amniotic fluid-derived cells into osteogenic cells. Attempts have been made to produce osteoblastic cells from amniotic fluid samples in a very short time, using a single step culture procedure, which allows a 20-d reduction in culture time[60].
One notes that several studies have demonstrated how such osteoinductive molecules are able to improve the osteogenic differentiation process[44,63-68].
In detail, then, AF-MSCs transfected with a defective adenoviral vector expressing human lim mineralization protein 3, an intracellular positive regulator of osteoblast differentiation, showed downregulation of stemness kruppel-like factor-4 and then increased osteogenic differentiation[63].
Again, Simvastatin, a 3-hydroxy-3-methylglutaryl-coenzyme A reductase inhibitor able to act on cholesterol endogenous synthesis, has been shown to be efficient in stimulating new bone formation. When added to osteogenic differentiation medium, Simvastatin was able to induce massive osteogenic differentiation of AF-MSCs, as observed by Alizarin Red S staining, and increased expression of typical osteogenic genes, such as osteopontin (OPN) and osteocalcin[64].
Herbal medicines have also been used to improve the osteogenic differentiation of AF-MSCs. Naringin, the main active compound of Chinese herbal medicine (Rhizoma drynariae), proved able to enhance osteogenic differentiation, increasing ALP activity and the expression of the osteogenic specific markers OPN, Collagen I, bone morphogenetic protein 4 (BMP4), RUNX2, β-catenin and cyclin D1[66].
Another traditional Chinese herbal medicine commonly used in treating orthopedic disorders, Curculigoside, was found to have a positive effect on the osteogenic differentiation of human AFSCs[65].
Moreover, during osteogenic differentiation of both amniotic fluid and dental pulp stem cells, the in vitro addition of Ferutinin, a phytoestrogen able to prevent osteoporosis caused by ovariectomy-induced estrogen deficiency, was efficacious in enhancing the production of a calcified matrix[67].
It has been noted that, in co-culture with osteoblasts obtained from the differentiation of dental pulp stem cells AF-MSCs prove able to generate osteoblasts. The same effect was observed when AF-MSCs were cultured in the conditioned medium of osteoblasts-differentiated dental pulp stem cells[69].
Moreover, some findings show that microRNAs (miRNAs) are involved in determining the fate of stem cells[70]. For example, a recent paper has shown the role of miR-21 in accelerating osteogenesis of AF-MSCs[71]. In that work two cellular populations were isolated from amniotic fluid: spindle-shaped (SS) and round-shaped (RS). Interestingly, induction of miR-21 was found to accelerate osteogenesis more in the SS population than in RS cells[71].
Finally, human AF-MSCs analyzed by an atomic force microscope during osteogenic differentiation showed a decrease in cell elasticity, which is typical of mature osteoblasts; thus the mechanical properties of AF-MSCs again add to the interest in applying them in bone regenerative medicine[72].
Up to now, little is known about the cues regulating the AF-MSCs’ ability to differentiate to osteoblasts. In depth study of the mechanism involved in the osteogenesis of AF-MSCs may hence help to develop standard protocols for clinical application in bone regenerative medicine. In this context, there is some evidence to suggest a role by the canonical Wnt signal pathway in bone formation as activation of this pathway stimulates osteoprogenitor proliferation and osteogenesis of human MSCs[73,74]. Wnt signaling is also involved in AF-MSC commitment toward osteogenesis[13].
Recently, we identified for the first time, the presence of Calcium Sensing Receptor (CaSR) in ovine and human AF-MSCs[44,68]. CaSR, originally cloned from parathyroid glands, acts by controlling the secretion of parathyroid hormone in response to changes in extracellular calcium levels[75]. However, it is well known that CaSR plays an important role in controlling osteoblasts as well as in osteoclast recruitment, differentiation and survival via multiple intracellular signals[76]. Interestingly, we observed that CaSR expression in both ovine and human AF-MSCs increased at the membrane when cells were treated with calcimimetic R-568, a molecule able to modulate bone cell metabolism via CaSR[77]. This effect was abolished by CaSR allosteric inhibitor Calhex-231 and by selective inhibitor NPS-2143. Importantly, downregulation of CaSR by a gene-silencing approach confirmed the crucial role of CaSR in supporting osteogenic differentiation[44].
These findings support the role of calcimimetics in the osteogenic differentiation of AF-MSCs, and suggest a strategy to develop therapy against bone injury.
Although the aforementioned studies have shown that AF-MSCs can be made to differentiate into osteoblasts in vitro[40,41] and some pathways have been investigated, more studies and clinical trials are needed before AF-MSCs can be applied clinically for in vivo bone regeneration.
TISSUE ENGINEERING APPROACHES FOR IN VIVO BONE REGENERATION
For a long period time autogenous bone grafting was considered the gold standard for in vivo bone regeneration[78]. Because of its limited availability and potential donor site morbidity, several bone substitutes have been successfully tried as an alternative, combining principles of biology and engineering. In vivo bone regeneration based on tissue engineering using scaffolds offers a plausible way of creating a favorable microenvironment for cells[79].
The choice of an appropriate scaffold along with selection of the best suited cell source is currently being widely studied.
As discussed above, the advantages of using amniotic fluid-derived cells are well established. Their high proliferation and osteogenic differentiation ability, together with the possibility of autologous or non-immunogenic transplantation and the absence of teratoma formation, makes possible a three-dimensional application of amniotic fluid cells in bone regeneration.
In general, a scaffold is a three-dimensional construct able to support adhesion, proliferation, and function of appropriate cells[78]. Stimuli mimicking the in vivo bone environment are needed for tissue-engineered constructs. It is well known that the bone regeneration ability of MSCs is mainly due to their paracrine effects[80]. They secrete bioactive substances that are able to enhance migration, proliferation, and differentiation of the neighboring resident cells[81,82]. Thus, what is required is the right combination of cells and the most appropriate support and factors.
Various synthetic and non-synthetic scaffolds have been employed to support osteogenic differentiation of AF-MSCs. One commonly used non-synthetic scaffold is collagens, which are present in the bone tissue, where they stimulate MSCs to differentiate into osteoblasts, initiating new bone formation[78]. This natural scaffolds are frequently used for their high availability, biological plasticity, biocompatibility, biodegradability and non-toxicity[83,84].
It has been found that osteoblasts obtained from AF-MSCs were able to adhere and grow well on SLA (Sandblasted and Acid Etching) titanium surfaces, materials commonly utilized in dental implantology, as revealed by electron microscopy observation[60].
Berardinelli et al[85] have demonstrated that a biomimetic commercial scaffold (magnesium-enriched hydroxyapatite (MgHA)/collagen-based scaffold) engineered with ovine AF-MSCs improves bone regeneration in a sheep model of sinus augmentation. Of note, the surface of this commercial scaffold was able to entrap a very high concentration of cells (1 × 107 cells/cm2) under dynamic cultural conditions. The osteoinductive properties of this scaffold, together with the potential of ovine AF-MSCs clearly accelerated the formation of new bone.
Human AFSCs seeded on nanofibrous (NF) or solid walled (SW) scaffolds were induced in rhBMP-7-containing medium for 7 d and implanted into male outbred thymic nude mice (nu/nu). Six weeks after implantation, bone formation was found on scaffolds as noted by von Kossa staining with greater mineralization on NF than SW scaffolds[86].
Synthetic NF scaffolds were developed with a morphology similar to that of natural collagen fibers, the aim being to mimic the morphological function of collagen fibers[87].
Human AFSCs were differentiated on electrospun nanofiber meshes and compared to BM-MSCs. In these experimental conditions, the cells displayed a delay in alkaline phosphatase activity, but elevated mineral deposition after 4 wk in culture, compared to BM-MSCs[88]. The nanofiber mesh scaffold possesses high porosity, large surface-area-to-volume ratios and size scale similar to extracellular matrix components. It allows the attachment of the cells and act as an efficient vehicle to deliver them to a defective site. A study by Maraldi et al[89] demonstrated that mineralization of hAFSCs tended to be enhanced on fibroin scaffolds, better than collagen and poly-D,L-lactic acid scaffolds. The cells were pre-differentiated on scaffolds for one week and then bone formation in vivo was determined after subcutaneous implantation into immune-suppressed rats[89].
Again, human AFSCs were able to proliferate and differentiate into the osteogenic phenotype, producing mineralized extracellular matrix similar to BM-MSCs in 2D culture conditions[90]. Moreover, when seeded on microfibrous starch and a poly(ε-caprolactone) scaffold, AFSCs successfully differentiated into osteogenic cells. The high porosity of the scaffold influenced the sequential development of osteoblastic cells and improved the osteogenic differentiation process. Some differences from BM-MSCs were observed in the expression of RunX-2, collagen I and ALP activity, denoting that cells from different origins may express different osteogenic markers at different times[90].
In addition, AFSCs were also able to produce mineralized matrix within porous medical-grade poly-e-caprolactone (mPCL) scaffold. The latter has good mechanical stability, large surface area and high porosity, allowing the attachment of AFSCs and extracellular matrix deposition. The construct, composed of an mPCL scaffold and AFSCs pre-differentiated for 28 d, was succeeded in producing seven times more mineralized matrix when implanted subcutaneously on the dorsal side of athymic female nude rats[91].
The same cells also proved able to adhere to a composite scaffold formed of a collagen matrix derived from porcine bladder submucosa matrix and poly(lactide-co-glycolide). In this condition, the osteogenic differentiation of AFSCs was improved, as revealed by increased mineralization and upregulation of osteogenic genes[92]. Indeed, the combination of natural scaffold with synthetic polymers was able to provide a microenvironment to facilitate the osteogenic differentiation.
More recently, a construct composed of collagen type I and AFSCs was used to evaluate bone regeneration in critical-sized rat calvarial defect after oral administration of phytoestrogen ferutinin[93]. Interestingly, 4 wk after implantation, the construct had reconstructed almost 70% of the critical-size defect[93]. Collagen type I, the major component of the extracellular matrix, facilitated cell adhesion and bone-forming cells in the defect site.
Another interesting study, by Mirabella et al[38] demonstrated that AFSCs loaded onto a biomaterial named Skelite and subcutaneously implanted in the back of CD-1 nu/nu mice gave rise to mineralized bone. Skelite disk is composed of silicon stabilized tricalcium phosphate biomaterial with a porosity of approximately 60% and an open structure similar to bone. However, they found that AFSCs did not have a direct role in new bone formation, but contributed to host progenitor recruitment and vessel formation in the engineered bone.
The unique features of AF-derived cells combined with the interesting results obtained in animal models of skeletal damage (Table 2) make them promising for the regeneration of bone tissue. Thanks to their immunosuppressive properties, allogeneic transplantation of AF-MSCs may be applied in various orthopedic conditions, and cell banks may be set up for regenerative medicine (Figure 1). However, more accurate understanding of the behavior and homing of AF-MSCs in vivo is necessary before they can be clinically applied. Moreover, additional studies are required to improve implanted cell survival and to ascertain the best biomaterial and the optimal combination of cytokines and growth factors.
Table 2.
Studies describing in vivo osteogenic differentiation of AF-derived cells
| Cell source | Scaffold | Animal model | Ref. |
| oAF-MSCs | Magnesium-enriched hydroxyapatite/collagen-based scaffold | Sheep model of sinus augumentation | [85] |
| hAFSCs | Nanofibrous or solid walled scaffolds | Midsagittal incision made on the dorsa and two subcutaneous pockets created using blunt dissection on male outbred thymic nude mouse model (nu/nu) | [86] |
| hAFSCs | Fibroin scaffolds compared to collagen and poly-D,L-lactic acid scaffolds | Dorsal mid-sagittal incision made on the dorsa and two subcutaneous pockets were created using blunt dissection male outbred rats | [89] |
| hAFSCs | Microfibrous starch and poly(ε-caprolactone) scaffold | Subcutaneous implantation into the dorsal side of athymic female nude rats | [91] |
| hAFSCs | Construct composed of collagen type I | Critical-sized rat calvarial defect after oral administration of phytoestrogen ferutinin | [93] |
| hAFSCs | Biomaterial Skelite | Subcutaneous implantation in the back of CD-1 nu/nu mice | [38] |
MSCs: Mesenchymal stromal cells; AF-MSCs: MSCs isolated from amniotic fluid; AFSCs: Amniotic fluid stem cells.
CONCLUSION
One purpose of bone regeneration is to find the most appropriate source of stem cells for clinical application together with the best bio-compatible support.
Bone marrow harvesting requires invasive procedures while the MSC number and differentiation potential decline with increasing age of the donor[94].
MSCs from extra-embryonic tissues are easily accessible. Moreover, their osteogenic potential and their safety in humans have been already tested. What is more, considering the high number of MSCs that can be isolated from amniotic fluid, a possibility of banking these cells adds greater promise of their clinical application. Furthermore, the osteogenic commitment of AF-MSCs could be enhanced by using appropriate osteoconductive scaffolds and osteoinductive growth factors[90].
Although these cells are very promising, therefore, in order to apply them for bone regeneration further investigations are needed to select the safest and most efficient cell-based approach. Certainly the results obtained so far are most promising: preclinical and clinical studies should be continued, thus opening new insights in the foreseeable future.
Footnotes
Conflict-of-interest: The authors do not conflict of interests.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: December 14, 2014
First decision: December 26, 2014
Article in press: February 11, 2015
P- Reviewer: Cai KY, Kiselev SL, Song GB, Zhang Q S- Editor: Ji FF L- Editor: A E- Editor: Wu HL
References
- 1.Cauley JA. Public health impact of osteoporosis. J Gerontol A Biol Sci Med Sci. 2013;68:1243–1251. doi: 10.1093/gerona/glt093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Jimi E, Hirata S, Osawa K, Terashita M, Kitamura C, Fukushima H. The current and future therapies of bone regeneration to repair bone defects. Int J Dent. 2012;2012:148261. doi: 10.1155/2012/148261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Waese EY, Kandel RA, Stanford WL. Application of stem cells in bone repair. Skeletal Radiol. 2008;37:601–608. doi: 10.1007/s00256-007-0438-8. [DOI] [PubMed] [Google Scholar]
- 4.Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A, Prockop Dj, Horwitz E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315–317. doi: 10.1080/14653240600855905. [DOI] [PubMed] [Google Scholar]
- 5.Liu ZJ, Zhuge Y, Velazquez OC. Trafficking and differentiation of mesenchymal stem cells. J Cell Biochem. 2009;106:984–991. doi: 10.1002/jcb.22091. [DOI] [PubMed] [Google Scholar]
- 6.Harada S, Rodan GA. Control of osteoblast function and regulation of bone mass. Nature. 2003;423:349–355. doi: 10.1038/nature01660. [DOI] [PubMed] [Google Scholar]
- 7.Bielby R, Jones E, McGonagle D. The role of mesenchymal stem cells in maintenance and repair of bone. Injury. 2007;38 Suppl 1:S26–S32. doi: 10.1016/j.injury.2007.02.007. [DOI] [PubMed] [Google Scholar]
- 8.Phimphilai M, Zhao Z, Boules H, Roca H, Franceschi RT. BMP signaling is required for RUNX2-dependent induction of the osteoblast phenotype. J Bone Miner Res. 2006;21:637–646. doi: 10.1359/JBMR.060109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rawadi G, Vayssière B, Dunn F, Baron R, Roman-Roman S. BMP-2 controls alkaline phosphatase expression and osteoblast mineralization by a Wnt autocrine loop. J Bone Miner Res. 2003;18:1842–1853. doi: 10.1359/jbmr.2003.18.10.1842. [DOI] [PubMed] [Google Scholar]
- 10.Gimble JM, Zvonic S, Floyd ZE, Kassem M, Nuttall ME. Playing with bone and fat. J Cell Biochem. 2006;98:251–266. doi: 10.1002/jcb.20777. [DOI] [PubMed] [Google Scholar]
- 11.James AW. Review of Signaling Pathways Governing MSC Osteogenic and Adipogenic Differentiation. Scientifica (Cairo) 2013;2013:684736. doi: 10.1155/2013/684736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kang Q, Song WX, Luo Q, Tang N, Luo J, Luo X, Chen J, Bi Y, He BC, Park JK, et al. A comprehensive analysis of the dual roles of BMPs in regulating adipogenic and osteogenic differentiation of mesenchymal progenitor cells. Stem Cells Dev. 2009;18:545–559. doi: 10.1089/scd.2008.0130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.D’Alimonte I, Lannutti A, Pipino C, Di Tomo P, Pierdomenico L, Cianci E, Antonucci I, Marchisio M, Romano M, Stuppia L, et al. Wnt signaling behaves as a “master regulator” in the osteogenic and adipogenic commitment of human amniotic fluid mesenchymal stem cells. Stem Cell Rev. 2013;9:642–654. doi: 10.1007/s12015-013-9436-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.James AW, Leucht P, Levi B, Carre AL, Xu Y, Helms JA, Longaker MT. Sonic Hedgehog influences the balance of osteogenesis and adipogenesis in mouse adipose-derived stromal cells. Tissue Eng Part A. 2010;16:2605–2616. doi: 10.1089/ten.tea.2010.0048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.James AW, Pang S, Askarinam A, Corselli M, Zara JN, Goyal R, Chang L, Pan A, Shen J, Yuan W, et al. Additive effects of sonic hedgehog and Nell-1 signaling in osteogenic versus adipogenic differentiation of human adipose-derived stromal cells. Stem Cells Dev. 2012;21:2170–2178. doi: 10.1089/scd.2011.0461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Knight MN, Hankenson KD. Mesenchymal Stem Cells in Bone Regeneration. Adv Wound Care (New Rochelle) 2013;2:306–316. doi: 10.1089/wound.2012.0420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Caplan AI. Review: mesenchymal stem cells: cell-based reconstructive therapy in orthopedics. Tissue Eng. 2005;11:1198–1211. doi: 10.1089/ten.2005.11.1198. [DOI] [PubMed] [Google Scholar]
- 18.Wang X, Wang Y, Gou W, Lu Q, Peng J, Lu S. Role of mesenchymal stem cells in bone regeneration and fracture repair: a review. Int Orthop. 2013;37:2491–2498. doi: 10.1007/s00264-013-2059-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Weissman IL. Translating stem and progenitor cell biology to the clinic: barriers and opportunities. Science. 2000;287:1442–1446. doi: 10.1126/science.287.5457.1442. [DOI] [PubMed] [Google Scholar]
- 20.da Silva Meirelles L, Chagastelles PC, Nardi NB. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J Cell Sci. 2006;119:2204–2213. doi: 10.1242/jcs.02932. [DOI] [PubMed] [Google Scholar]
- 21.Zvaifler NJ, Marinova-Mutafchieva L, Adams G, Edwards CJ, Moss J, Burger JA, Maini RN. Mesenchymal precursor cells in the blood of normal individuals. Arthritis Res. 2000;2:477–488. doi: 10.1186/ar130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhu X, Shi W, Tai W, Liu F. The comparition of biological characteristics and multilineage differentiation of bone marrow and adipose derived Mesenchymal stem cells. Cell Tissue Res. 2012;350:277–287. doi: 10.1007/s00441-012-1453-1. [DOI] [PubMed] [Google Scholar]
- 23.Park JR, Kim E, Yang J, Lee H, Hong SH, Woo HM, Park SM, Na S, Yang SR. Isolation of human dermis derived mesenchymal stem cells using explants culture method: expansion and phenotypical characterization. Cell Tissue Bank. 2014:Aug 28; Epub ahead of print. doi: 10.1007/s10561-014-9471-8. [DOI] [PubMed] [Google Scholar]
- 24.Wagner W, Wein F, Seckinger A, Frankhauser M, Wirkner U, Krause U, Blake J, Schwager C, Eckstein V, Ansorge W, et al. Comparative characteristics of mesenchymal stem cells from human bone marrow, adipose tissue, and umbilical cord blood. Exp Hematol. 2005;33:1402–1416. doi: 10.1016/j.exphem.2005.07.003. [DOI] [PubMed] [Google Scholar]
- 25.Campagnoli C, Roberts IA, Kumar S, Bennett PR, Bellantuono I, Fisk NM. Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver, and bone marrow. Blood. 2001;98:2396–2402. doi: 10.1182/blood.v98.8.2396. [DOI] [PubMed] [Google Scholar]
- 26.Guillot PV, De Bari C, Dell’Accio F, Kurata H, Polak J, Fisk NM. Comparative osteogenic transcription profiling of various fetal and adult mesenchymal stem cell sources. Differentiation. 2008;76:946–957. doi: 10.1111/j.1432-0436.2008.00279.x. [DOI] [PubMed] [Google Scholar]
- 27.Fauza D. Amniotic fluid and placental stem cells. Best Pract Res Clin Obstet Gynaecol. 2004;18:877–891. doi: 10.1016/j.bpobgyn.2004.07.001. [DOI] [PubMed] [Google Scholar]
- 28.Pipino C, Shangaris P, Resca E, Zia S, Deprest J, Sebire NJ, David AL, Guillot PV, De Coppi P. Placenta as a reservoir of stem cells: an underutilized resource? Br Med Bull. 2013;105:43–68. doi: 10.1093/bmb/lds033. [DOI] [PubMed] [Google Scholar]
- 29.Secco M, Zucconi E, Vieira NM, Fogaça LL, Cerqueira A, Carvalho MD, Jazedje T, Okamoto OK, Muotri AR, Zatz M. Mesenchymal stem cells from umbilical cord: do not discard the cord! Neuromuscul Disord. 2008;18:17–18. doi: 10.1016/j.nmd.2007.11.003. [DOI] [PubMed] [Google Scholar]
- 30.Abdulrazzak H, Moschidou D, Jones G, Guillot PV. Biological characteristics of stem cells from foetal, cord blood and extraembryonic tissues. J R Soc Interface. 2010;7 Suppl 6:S689–S706. doi: 10.1098/rsif.2010.0347.focus. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bieback K, Kern S, Klüter H, Eichler H. Critical parameters for the isolation of mesenchymal stem cells from umbilical cord blood. Stem Cells. 2004;22:625–634. doi: 10.1634/stemcells.22-4-625. [DOI] [PubMed] [Google Scholar]
- 32.Siegel N, Rosner M, Hanneder M, Valli A, Hengstschläger M. Stem cells in amniotic fluid as new tools to study human genetic diseases. Stem Cell Rev. 2007;3:256–264. doi: 10.1007/s12015-007-9003-z. [DOI] [PubMed] [Google Scholar]
- 33.Shaw SW, Blundell MP, Pipino C, Shangaris P, Maghsoudlou P, Ramachandra DL, Georgiades F, Boyd M, Thrasher AJ, Porada CD, et al. Sheep CD34+ amniotic fluid cells have hematopoietic potential and engraft after autologous in utero transplantation. Stem Cells. 2015;33:122–132. doi: 10.1002/stem.1839. [DOI] [PubMed] [Google Scholar]
- 34.Cananzi M, Atala A, De Coppi P. Stem cells derived from amniotic fluid: new potentials in regenerative medicine. Reprod Biomed Online. 2009;18 Suppl 1:17–27. doi: 10.1016/s1472-6483(10)60111-3. [DOI] [PubMed] [Google Scholar]
- 35.Carraro G, Perin L, Sedrakyan S, Giuliani S, Tiozzo C, Lee J, Turcatel G, De Langhe SP, Driscoll B, Bellusci S, et al. Human amniotic fluid stem cells can integrate and differentiate into epithelial lung lineages. Stem Cells. 2008;26:2902–2911. doi: 10.1634/stemcells.2008-0090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bollini S, Cheung KK, Riegler J, Dong X, Smart N, Ghionzoli M, Loukogeorgakis SP, Maghsoudlou P, Dubé KN, Riley PR, et al. Amniotic fluid stem cells are cardioprotective following acute myocardial infarction. Stem Cells Dev. 2011;20:1985–1994. doi: 10.1089/scd.2010.0424. [DOI] [PubMed] [Google Scholar]
- 37.De Coppi P, Callegari A, Chiavegato A, Gasparotto L, Piccoli M, Taiani J, Pozzobon M, Boldrin L, Okabe M, Cozzi E, et al. Amniotic fluid and bone marrow derived mesenchymal stem cells can be converted to smooth muscle cells in the cryo-injured rat bladder and prevent compensatory hypertrophy of surviving smooth muscle cells. J Urol. 2007;177:369–376. doi: 10.1016/j.juro.2006.09.103. [DOI] [PubMed] [Google Scholar]
- 38.Mirabella T, Gentili C, Daga A, Cancedda R. Amniotic fluid stem cells in a bone microenvironment: driving host angiogenic response. Stem Cell Res. 2013;11:540–551. doi: 10.1016/j.scr.2013.02.006. [DOI] [PubMed] [Google Scholar]
- 39.Bossolasco P, Montemurro T, Cova L, Zangrossi S, Calzarossa C, Buiatiotis S, Soligo D, Bosari S, Silani V, Deliliers GL, et al. Molecular and phenotypic characterization of human amniotic fluid cells and their differentiation potential. Cell Res. 2006;16:329–336. doi: 10.1038/sj.cr.7310043. [DOI] [PubMed] [Google Scholar]
- 40.De Coppi P, Bartsch G, Siddiqui MM, Xu T, Santos CC, Perin L, Mostoslavsky G, Serre AC, Snyder EY, Yoo JJ, et al. Isolation of amniotic stem cell lines with potential for therapy. Nat Biotechnol. 2007;25:100–106. doi: 10.1038/nbt1274. [DOI] [PubMed] [Google Scholar]
- 41.In ‘t Anker PS, Scherjon SA, Kleijburg-van der Keur C, Noort WA, Claas FH, Willemze R, Fibbe WE, Kanhai HH. Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood. 2003;102:1548–1549. doi: 10.1182/blood-2003-04-1291. [DOI] [PubMed] [Google Scholar]
- 42.Kaviani A, Perry TE, Dzakovic A, Jennings RW, Ziegler MM, Fauza DO. The amniotic fluid as a source of cells for fetal tissue engineering. J Pediatr Surg. 2001;36:1662–1665. doi: 10.1053/jpsu.2001.27945. [DOI] [PubMed] [Google Scholar]
- 43.Sessarego N, Parodi A, Podestà M, Benvenuto F, Mogni M, Raviolo V, Lituania M, Kunkl A, Ferlazzo G, Bricarelli FD, et al. Multipotent mesenchymal stromal cells from amniotic fluid: solid perspectives for clinical application. Haematologica. 2008;93:339–346. doi: 10.3324/haematol.11869. [DOI] [PubMed] [Google Scholar]
- 44.Pipino C, Di Tomo P, Mandatori D, Cianci E, Lanuti P, Cutrona MB, Penolazzi L, Pierdomenico L, Lambertini E, Antonucci I, et al. Calcium sensing receptor activation by calcimimetic R-568 in human amniotic fluid mesenchymal stem cells: correlation with osteogenic differentiation. Stem Cells Dev. 2014;23:2959–2971. doi: 10.1089/scd.2013.0627. [DOI] [PubMed] [Google Scholar]
- 45.Pozzobon M, Ghionzoli M, De Coppi P. ES, iPS, MSC, and AFS cells. Stem cells exploitation for Pediatric Surgery: current research and perspective. Pediatr Surg Int. 2010;26:3–10. doi: 10.1007/s00383-009-2478-8. [DOI] [PubMed] [Google Scholar]
- 46.Sarić T, Frenzel LP, Hescheler J. Immunological barriers to embryonic stem cell-derived therapies. Cells Tissues Organs. 2008;188:78–90. doi: 10.1159/000118784. [DOI] [PubMed] [Google Scholar]
- 47.Hentze H, Graichen R, Colman A. Cell therapy and the safety of embryonic stem cell-derived grafts. Trends Biotechnol. 2007;25:24–32. doi: 10.1016/j.tibtech.2006.10.010. [DOI] [PubMed] [Google Scholar]
- 48.Grinnemo KH, Sylvén C, Hovatta O, Dellgren G, Corbascio M. Immunogenicity of human embryonic stem cells. Cell Tissue Res. 2008;331:67–78. doi: 10.1007/s00441-007-0486-3. [DOI] [PubMed] [Google Scholar]
- 49.Daley GQ, Ahrlund Richter L, Auerbach JM, Benvenisty N, Charo RA, Chen G, Deng HK, Goldstein LS, Hudson KL, Hyun I, et al. Ethics. The ISSCR guidelines for human embryonic stem cell research. Science. 2007;315:603–604. doi: 10.1126/science.1139337. [DOI] [PubMed] [Google Scholar]
- 50.Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131:861–872. doi: 10.1016/j.cell.2007.11.019. [DOI] [PubMed] [Google Scholar]
- 51.Okita K, Yamanaka S. Induced pluripotent stem cells: opportunities and challenges. Philos Trans R Soc Lond B Biol Sci. 2011;366:2198–2207. doi: 10.1098/rstb.2011.0016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mukherjee S, Thrasher AJ. iPSCs: Unstable origins? Mol Ther. 2011;19:1188–1190. doi: 10.1038/mt.2011.116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Galende E, Karakikes I, Edelmann L, Desnick RJ, Kerenyi T, Khoueiry G, Lafferty J, McGinn JT, Brodman M, Fuster V, et al. Amniotic fluid cells are more efficiently reprogrammed to pluripotency than adult cells. Cell Reprogram. 2010;12:117–125. doi: 10.1089/cell.2009.0077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Pipino C, Mukherjee S, David AL, Blundell MP, Shaw SW, Sung P, Shangaris P, Waters JJ, Ellershaw D, Cavazzana M, et al. Trisomy 21 mid-trimester amniotic fluid induced pluripotent stem cells maintain genetic signatures during reprogramming: implications for disease modeling and cryobanking. Cell Reprogram. 2014;16:331–344. doi: 10.1089/cell.2013.0091. [DOI] [PubMed] [Google Scholar]
- 55.Spinelli V, Guillot PV, De Coppi P. Induced pluripotent stem (iPS) cells from human fetal stem cells (hFSCs) Organogenesis. 2013;9:101–110. doi: 10.4161/org.25197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Moschidou D, Mukherjee S, Blundell MP, Drews K, Jones GN, Abdulrazzak H, Nowakowska B, Phoolchund A, Lay K, Ramasamy TS, et al. Valproic acid confers functional pluripotency to human amniotic fluid stem cells in a transgene-free approach. Mol Ther. 2012;20:1953–1967. doi: 10.1038/mt.2012.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ramachandra DL, Shaw SS, Shangaris P, Loukogeorgakis S, Guillot PV, Coppi PD, David AL. In utero therapy for congenital disorders using amniotic fluid stem cells. Front Pharmacol. 2014;5:270. doi: 10.3389/fphar.2014.00270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Mauro A, Turriani M, Ioannoni A, Russo V, Martelli A, Di Giacinto O, Nardinocchi D, Berardinelli P. Isolation, characterization, and in vitro differentiation of ovine amniotic stem cells. Vet Res Commun. 2010;34 Suppl 1:S25–S28. doi: 10.1007/s11259-010-9393-2. [DOI] [PubMed] [Google Scholar]
- 59.Liu ZS, Xu YF, Feng SW, Li Y, Yao XL, Lu XL, Zhang C. Baculovirus-transduced mouse amniotic fluid-derived stem cells maintain differentiation potential. Ann Hematol. 2009;88:565–572. doi: 10.1007/s00277-008-0634-1. [DOI] [PubMed] [Google Scholar]
- 60.Antonucci I, Iezzi I, Morizio E, Mastrangelo F, Pantalone A, Mattioli-Belmonte M, Gigante A, Salini V, Calabrese G, Tetè S, et al. Isolation of osteogenic progenitors from human amniotic fluid using a single step culture protocol. BMC Biotechnol. 2009;9:9. doi: 10.1186/1472-6750-9-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Liu Y, Wu J, Zhu Y, Han J. Therapeutic application of mesenchymal stem cells in bone and joint diseases. Clin Exp Med. 2014;14:13–24. doi: 10.1007/s10238-012-0218-1. [DOI] [PubMed] [Google Scholar]
- 62.Liao HT, Chen CT. Osteogenic potential: Comparison between bone marrow and adipose-derived mesenchymal stem cells. World J Stem Cells. 2014;6:288–295. doi: 10.4252/wjsc.v6.i3.288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Barba M, Pirozzi F, Saulnier N, Vitali T, Natale MT, Logroscino G, Robbins PD, Gambotto A, Neri G, Michetti F, et al. Lim mineralization protein 3 induces the osteogenic differentiation of human amniotic fluid stromal cells through Kruppel-like factor-4 downregulation and further bone-specific gene expression. J Biomed Biotechnol. 2012;2012:813894. doi: 10.1155/2012/813894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.de Lara Janz F, Favero GM, Bohatch MS, Aguiar Debes A, Bydlowski SP. Simvastatin induces osteogenic differentiation in human amniotic fluid mesenchymal stem cells (AFMSC) Fundam Clin Pharmacol. 2014;28:211–216. doi: 10.1111/fcp.12006. [DOI] [PubMed] [Google Scholar]
- 65.Liu M, Li Y, Yang ST. Curculigoside improves osteogenesis of human amniotic fluid-derived stem cells. Stem Cells Dev. 2014;23:146–154. doi: 10.1089/scd.2013.0261. [DOI] [PubMed] [Google Scholar]
- 66.Liu M, Li Y, Yang ST. Effects of naringin on the proliferation and osteogenic differentiation of human amniotic fluid-derived stem cells. J Tissue Eng Regen Med. 2014:Jun 11; Epub ahead of print. doi: 10.1002/term.1911. [DOI] [PubMed] [Google Scholar]
- 67.Zavatti M, Resca E, Bertoni L, Maraldi T, Guida M, Carnevale G, Ferrari A, De Pol A. Ferutinin promotes proliferation and osteoblastic differentiation in human amniotic fluid and dental pulp stem cells. Life Sci. 2013;92:993–1003. doi: 10.1016/j.lfs.2013.03.018. [DOI] [PubMed] [Google Scholar]
- 68.Di Tomo P, Pipino C, Lanuti P, Morabito C, Pierdomenico L, Sirolli V, Bonomini M, Miscia S, Mariggiò MA, Marchisio M, et al. Calcium sensing receptor expression in ovine amniotic fluid mesenchymal stem cells and the potential role of R-568 during osteogenic differentiation. PLoS One. 2013;8:e73816. doi: 10.1371/journal.pone.0073816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.De Rosa A, Tirino V, Paino F, Tartaglione A, Mitsiadis T, Feki A, d’Aquino R, Laino L, Colacurci N, Papaccio G. Amniotic fluid-derived mesenchymal stem cells lead to bone differentiation when cocultured with dental pulp stem cells. Tissue Eng Part A. 2011;17:645–653. doi: 10.1089/ten.TEA.2010.0340. [DOI] [PubMed] [Google Scholar]
- 70.Choi E, Choi E, Hwang KC. MicroRNAs as novel regulators of stem cell fate. World J Stem Cells. 2013;5:172–187. doi: 10.4252/wjsc.v5.i4.172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Trohatou O, Zagoura D, Bitsika V, Pappa KI, Antsaklis A, Anagnou NP, Roubelakis MG. Sox2 suppression by miR-21 governs human mesenchymal stem cell properties. Stem Cells Transl Med. 2014;3:54–68. doi: 10.5966/sctm.2013-0081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Chen Q, Xiao P, Chen JN, Cai JY, Cai XF, Ding H, Pan YL. AFM studies of cellular mechanics during osteogenic differentiation of human amniotic fluid-derived stem cells. Anal Sci. 2010;26:1033–1037. doi: 10.2116/analsci.26.1033. [DOI] [PubMed] [Google Scholar]
- 73.Macsai CE, Foster BK, Xian CJ. Roles of Wnt signalling in bone growth, remodelling, skeletal disorders and fracture repair. J Cell Physiol. 2008;215:578–587. doi: 10.1002/jcp.21342. [DOI] [PubMed] [Google Scholar]
- 74.Liu G, Vijayakumar S, Grumolato L, Arroyave R, Qiao H, Akiri G, Aaronson SA. Canonical Wnts function as potent regulators of osteogenesis by human mesenchymal stem cells. J Cell Biol. 2009;185:67–75. doi: 10.1083/jcb.200810137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Alfadda TI, Saleh AM, Houillier P, Geibel JP. Calcium-sensing receptor 20 years later. Am J Physiol Cell Physiol. 2014;307:C221–C231. doi: 10.1152/ajpcell.00139.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Yamaguchi T. The calcium-sensing receptor in bone. J Bone Miner Metab. 2008;26:301–311. doi: 10.1007/s00774-008-0843-7. [DOI] [PubMed] [Google Scholar]
- 77.Marie PJ. The calcium-sensing receptor in bone cells: a potential therapeutic target in osteoporosis. Bone. 2010;46:571–576. doi: 10.1016/j.bone.2009.07.082. [DOI] [PubMed] [Google Scholar]
- 78.Oryan A, Alidadi S, Moshiri A, Maffulli N. Bone regenerative medicine: classic options, novel strategies, and future directions. J Orthop Surg Res. 2014;9:18. doi: 10.1186/1749-799X-9-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Ma PX. Biomimetic materials for tissue engineering. Adv Drug Deliv Rev. 2008;60:184–198. doi: 10.1016/j.addr.2007.08.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Osugi M, Katagiri W, Yoshimi R, Inukai T, Hibi H, Ueda M. Conditioned media from mesenchymal stem cells enhanced bone regeneration in rat calvarial bone defects. Tissue Eng Part A. 2012;18:1479–1489. doi: 10.1089/ten.tea.2011.0325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Hilfiker A, Kasper C, Hass R, Haverich A. Mesenchymal stem cells and progenitor cells in connective tissue engineering and regenerative medicine: is there a future for transplantation? Langenbecks Arch Surg. 2011;396:489–497. doi: 10.1007/s00423-011-0762-2. [DOI] [PubMed] [Google Scholar]
- 82.Meirelles Lda S, Fontes AM, Covas DT, Caplan AI. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009;20:419–427. doi: 10.1016/j.cytogfr.2009.10.002. [DOI] [PubMed] [Google Scholar]
- 83.Kikuchi M. Hydroxyapatite/collagen bone-like nanocomposite. Biol Pharm Bull. 2013;36:1666–1669. doi: 10.1248/bpb.b13-00460. [DOI] [PubMed] [Google Scholar]
- 84.Pastorino L, Dellacasa E, Scaglione S, Giulianelli M, Sbrana F, Vassalli M, Ruggiero C. Oriented collagen nanocoatings for tissue engineering. Colloids Surf B Biointerfaces. 2014;114:372–378. doi: 10.1016/j.colsurfb.2013.10.026. [DOI] [PubMed] [Google Scholar]
- 85.Berardinelli P, Valbonetti L, Muttini A, Martelli A, Peli R, Zizzari V, Nardinocchi D, Vulpiani MP, Tetè S, Barboni B, et al. Role of amniotic fluid mesenchymal cells engineered on MgHA/collagen-based scaffold allotransplanted on an experimental animal study of sinus augmentation. Clin Oral Investig. 2013;17:1661–1675. doi: 10.1007/s00784-012-0857-3. [DOI] [PubMed] [Google Scholar]
- 86.Sun H, Feng K, Hu J, Soker S, Atala A, Ma PX. Osteogenic differentiation of human amniotic fluid-derived stem cells induced by bone morphogenetic protein-7 and enhanced by nanofibrous scaffolds. Biomaterials. 2010;31:1133–1139. doi: 10.1016/j.biomaterials.2009.10.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Woo KM, Jun JH, Chen VJ, Seo J, Baek JH, Ryoo HM, Kim GS, Somerman MJ, Ma PX. Nano-fibrous scaffolding promotes osteoblast differentiation and biomineralization. Biomaterials. 2007;28:335–343. doi: 10.1016/j.biomaterials.2006.06.013. [DOI] [PubMed] [Google Scholar]
- 88.Kolambkar YM, Peister A, Ekaputra AK, Hutmacher DW, Guldberg RE. Colonization and osteogenic differentiation of different stem cell sources on electrospun nanofiber meshes. Tissue Eng Part A. 2010;16:3219–3230. doi: 10.1089/ten.tea.2010.0004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Maraldi T, Riccio M, Resca E, Pisciotta A, La Sala GB, Ferrari A, Bruzzesi G, Motta A, Migliaresi C, Marzona L, et al. Human amniotic fluid stem cells seeded in fibroin scaffold produce in vivo mineralized matrix. Tissue Eng Part A. 2011;17:2833–2843. doi: 10.1089/ten.tea.2011.0062. [DOI] [PubMed] [Google Scholar]
- 90.Rodrigues MT, Lee SJ, Gomes ME, Reis RL, Atala A, Yoo JJ. Amniotic fluid-derived stem cells as a cell source for bone tissue engineering. Tissue Eng Part A. 2012;18:2518–2527. doi: 10.1089/ten.tea.2011.0672. [DOI] [PubMed] [Google Scholar]
- 91.Peister A, Deutsch ER, Kolambkar Y, Hutmacher DW, Guldberg RE. Amniotic fluid stem cells produce robust mineral deposits on biodegradable scaffolds. Tissue Eng Part A. 2009;15:3129–3138. doi: 10.1089/ten.tea.2008.0536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Kim J, Jeong SY, Ju YM, Yoo JJ, Smith TL, Khang G, Lee SJ, Atala A. In vitro osteogenic differentiation of human amniotic fluid-derived stem cells on a poly(lactide-co-glycolide) (PLGA)-bladder submucosa matrix (BSM) composite scaffold for bone tissue engineering. Biomed Mater. 2013;8:014107. doi: 10.1088/1748-6041/8/1/014107. [DOI] [PubMed] [Google Scholar]
- 93.Zavatti M, Bertoni L, Maraldi T, Resca E, Beretti F, Guida M, La Sala GB, De Pol A. Critical-size bone defect repair using amniotic fluid stem cell/collagen constructs: effect of oral ferutinin treatment in rats. Life Sci. 2015;121:174–183. doi: 10.1016/j.lfs.2014.10.020. [DOI] [PubMed] [Google Scholar]
- 94.Rosset P, Deschaseaux F, Layrolle P. Cell therapy for bone repair. Orthop Traumatol Surg Res. 2014;100:S107–S112. doi: 10.1016/j.otsr.2013.11.010. [DOI] [PubMed] [Google Scholar]