
Fig 7.
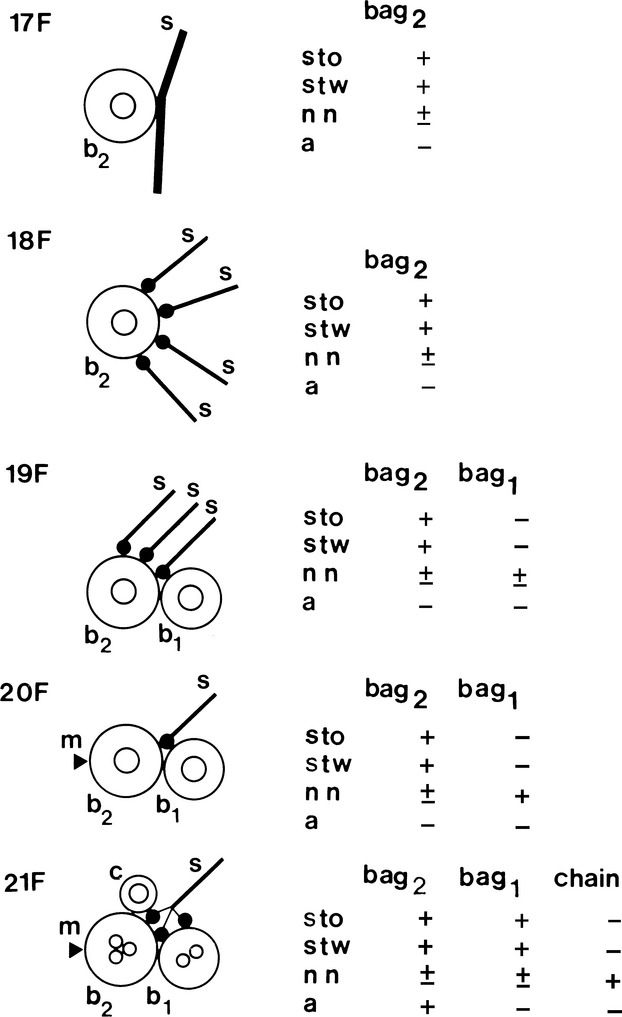
Simplified illustration of muscle spindle development showing the acquisition of sensory (s) and motor (m) innervation (based upon Kucera et al. 1989) and MyHC expression (based upon Pedrosa & Thornell, 1990). The earliest contacts between afferent terminals (s) and muscle cells, in rat soleus, are detectable on the 17th day of gestation (17F). Up till the 20th day in utero (20F), the nuclear bag2 myotubes (b2) are innervated by several afferents. Thereafter, each muscle spindle becomes innervated by a single primary afferent as in mature spindles, and the sensory endings are distributed to both immature nuclear bag2 and bag1 (b1) fibres and to the single intrafusal myoblasts that are the precursors of the nuclear chain fibres (c). Also by the 20th day in utero, the gamma motor innervation (arrowhead) reaches the muscle spindles initially innervating only the bag2 fibres. The nuclear bag1 and chain fibres acquire motor innervation later. Nuclear bag2 precursors are identifiable under light microscopy from the 17th day of gestation as the only primary myotubes expressing slow-tonic MyHC (sto), in addition to slow-twitch (stw) and neonatal (nn) MyHC. They acquire reactivity to anti-α-cardiac MyHC (a) by the 21st day of gestation. The future nuclear bag1 fibres are first seen at 19 days in utero as secondary myotubes in close apposition to the primary STO myotubes. Initially they express only neonatal MyHC and acquire reactivity to anti-slow-tonic and anti-slow-twitch MyHC 2 days later. Shortly before birth, the nuclear chain fibre precursors are identifiable as a 3rd myotube in each spindle strongly stained with anti-neonatal MyHC only. Three days after birth, the 2nd nuclear chain precursor arise and display a profile of reactivity identical to the 1st nuclear chain precursor. From Pedrosa et al. (1990).