
Fig. 4.
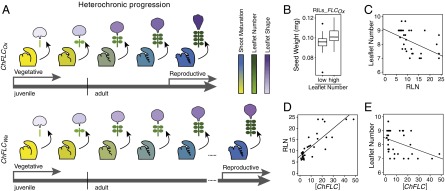
The impact of ChFLC on heteroblasty, the correlation with seed weight, and the prevalence of ChFLC-dependent leaflet number variation in C. hirsuta. (A) Cartoon summarizing how ChFLC-dependent shoot maturation influences heteroblasty in C. hirsuta. The weaker ChFLCOx allele causes increased leaflet number and rapid terminal leaflet shape changes at equivalent nodes compared with the stronger ChFLCWa allele, owing to faster progression through the adult vegetative phase of the shoot. Differential age-dependent variation of leaflet number and terminal leaflet shape results in adult leaves with divergent morphologies. In addition, the weaker allele results in earlier flowering. Gray arrows mark the juvenile, adult vegetative, and reproductive phases. Meristems with developing leaves are colored following a gradient from yellow (not competent to flower) to dark blue (flowering), which reflects shoot maturation. Schematic representations of heteroblastic leaf traits are colored following a gradient from low leaflet number and juvenile leaflet shape (light green and purple, respectively) to high leaflet number and adult leaflet shape (dark green and purple, respectively). Leaf primordia and lateral and terminal leaflets are not drawn to scale. (B) Boxplot showing the median and interquartile range of seed weight of RILs selected for their leaflet number and for having the Ox allele at the ChFLC locus. Mean seed weight of low leaflet number (5–5.6) and high leaflet number (6–8) RILs is significantly different (P < 0.05 by ANOVA). (C) Differences in RLN (x axis) can explain variation in leaflet number (y axis) in a sample of 33 C. hirsuta strains (R2 = 0.25; P < 0.03). (D and E) Variation in ChFLC expression measured by qRT-PCR (x axis) can explain differences in RLN (D; y axis; R2 = 0.68; P < 0.001; n = 35) and leaflet number on leaf 7 (E; y axis; R2 = 0.15; P < 0.01; n = 33). Transcript abundance is shown relative to that of the Ber-1 strain, which had the lowest ChFLC expression levels in our sample.