
Figure 5. PTAFR is required for ethanol-induced kidney tubule oxidation and PTAFR ligand formation.
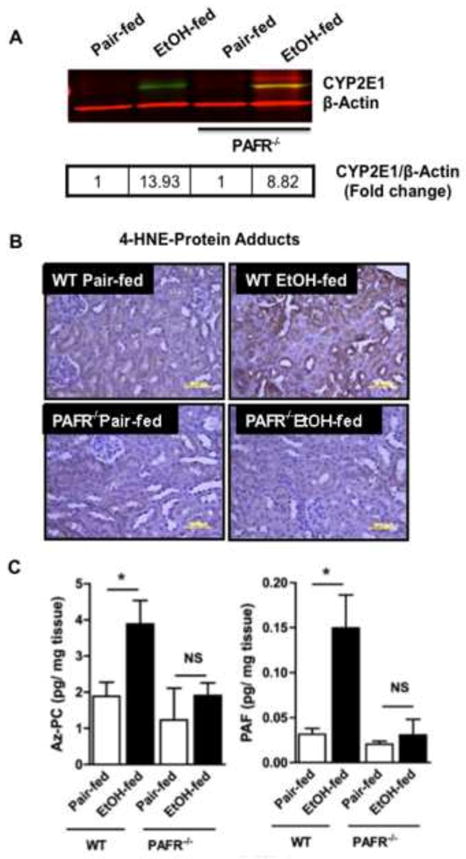
A) CYP2E1 induction is independent of PTAFR. Kidney homogenates from wild-type and PTAFR−/− mice fed control or ethanol-containing diets were resolved by SDS-PAGE (20 μg protein/well) prior to immunostaining for CYP2E1 with anti-rabbit CYP2E1 antibody followed by IRDye® 800 CW anti-rabbit secondary antibody prior to scanned in an Odyssey Imaging system. We again confirmed equal loading of protein simultaneously using anti-β-actin antibody and IRDye® 680 RD goat anti-mouse IgG1 secondary antibody. Fold change from pair fed control homogenates was calculated (box) following conversion to gray scale. B) 4-hydroxynonenal adduct formation in renal tubules requires PTAFR. Sections of kidney of C57/BL6 or PTAR−/− mice fed control or ethanol supplemented diets were immunostained with anti-4-hydroxynonenal antibody as in Fig. 3. The dark brown deposits of protein-4-hydroxynonenal adducts are representative of 2 to 3 images per kidney of 4 mice/group. C) PTAFR ligand formation in the kidneys of ethanol fed mice requires PTAFR. sn-2 Azelaoyl choline phospholipid (Az-PC) and Platelet-activating Factor (PAF) were quantified in kidneys of pair- and ethanol-fed wild-type and PTAFR−/− mice by liquid chromatography/electrospray ionization/tandem mass spectrometry (LC/MS/MS) using [2H]PAF as an internal standard as in the preceding figure. The data are expressed as mean±SEM (n=4), and p<0.05 was considered as significant.