
Fig. 5.
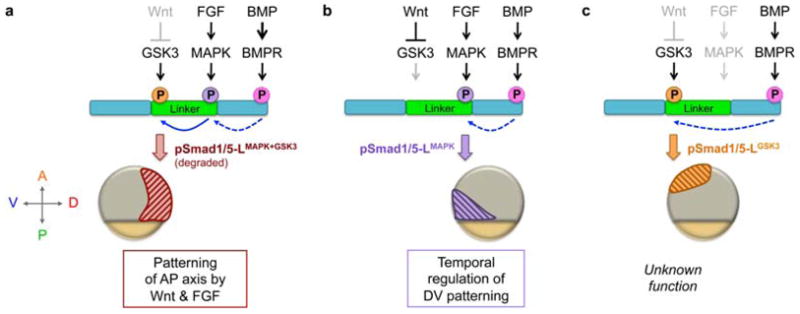
Model of spatially restricted functions of distinct phosphorylated Smad1/5 linker forms, depicted in mid-gastrula stage embryos. The N- and C-termini of Smad1/5 are shown in teal and the linker region is in green. C-terminal phosphorylation of Smad1/5 (Ct-pSmad1/5) a prerequisite to linker phosphorylation (dashed blue arrow). (a) In dorsal regions, MAPK and GSK3 sequentially phosphorylate the Smad1/5 linker (solid blue arrow), leading to pSmad1/5-LMAPK+GSK3 degradation and blockage of BMP signaling. This mechanism may be most important during blastula stages when BMP signaling is more widespread and present dorsally (Section 3.1). (b) In ventral-posterior regions both FGF and Wnt are present, activating MAPK and inhibiting GSK3, respectively, to generate pSmad1/5-LMAPK. pSmad1/5-LMAPK may have reduced activity, which regulates timing of DV patterning. During epiboly, pSmad1/5-LMAPK localizes progressively more posteriorly with the margin, patterning anterior DV tissues in its wake (not shown). (c) In animal regions GSK3 is uninhibited by Wnt, resulting in pSmad1/5-LGSK3, though its significance remains unknown. Since pSmad1/5-LGSK3 does not overlap with pSmad1/5-LMAPK, it is possible that in the zebrafish embryo (versus in vitro studies) either GSK3 does not require MAPK to prime the linker or those residues may be rapidly de-phosphorylated.