

Abstract
Short telomeres are known as one of the risk factors for human cancers. The present study was conducted to evaluate the association between 6 polymorphisms, which were related with short telomere length in the Korean population, and lung cancer risk using 1,100 cases and 1,096 controls. Among the 6 polymorphisms, TERT rs2853669 was significantly associated with increased lung cancer risk under a recessive model (odds ratio [OR]=1.38, 95% confidence interval [CI]=1.05-1.81, P=0.02). The effect of rs2853669 on lung cancer risk was significant in younger individuals (OR=1.73, 95% CI=1.18-2.54, P=0.005) and adenocarcinoma (OR=1.50, 95% CI=1.07-2.07, P=0.02). Our results suggest that a common functional promoter polymorphism, TERT rs2853669, may influence both telomere length and lung cancer risk in the Korean population.
Graphical Abstract
Keywords: Lung Cancer Risk, Polymorphism, Telomere Length, TERT
INTRODUCTION
The ends of chromosomes are composed of repetitive nucleotide sequences called telomeres. The main role of telomeres is to protect chromosome from degradation, end-to-end fusion, and atypical recombination by capping chromosome ends (1). Therefore, telomeres are essential for maintaining genomic integrity and stability (2). It is known that telomeres get shorter during mitosis due to incomplete replication of linear chromosomes by conventional DNA polymerases (1). When telomeres are shortened to a critical length, chromosome instability increases and cellular senescence or apoptosis occurs (3). Abnormally shortened telomeres can lead to diverse human diseases (4,5,6,7). A number of studies have also reported that short telomeres are associated with development of lung cancer (8,9,10).
Polymorphisms in telomere biology genes, such as telomerase reverse transcriptase (TERT) and telomerase RNA component (TERC), can influence telomere length. Recently, a number of candidate gene studies and genome-wide association studies (GWAS) have identified single nucleotide polymorphisms (SNPs) that are related to telomere length in Caucasians (11,12,13,14). In a previous study, we evaluated the telomere lengths of 17 SNPs related to telomere length in Caucasians and verified that 6 SNPs (TERT rs2853669, rs2736108, rs2736098, and rs7705526; TERC rs2293607; and ZNF676 rs412658) are associated with short telomere length in the Korean population (15). Here, we conducted a case-control study to investigate the association between the 6 SNPs related to short telomere length and lung cancer risk in the Korean population.
MATERIALS AND METHODS
Study population
This study involved 1,100 lung cancer patients and 1,096 healthy controls. The patients were first diagnosed with lung cancer between January 2008 and July 2012 at Kyungpook National University Hospital (KNUH), Daegu, Korea. There were no restrictions on gender, histologic subtype, or stage. The control subjects were frequency matched (1:1) to the cases based on gender and age (±5 yr) from a pool of healthy volunteers who visited the general healthy check-up center at KNUH during the same period. All subjects were of Korean ethnicity and residing in Daegu City or the surrounding regions. Blood samples were provided by the National Biobank of Korea, which is supported by the Ministry of Health, Welfare, and Family Affairs.
SNP selection and genotyping
In a previous study, we evaluated the associations between telomere length in the Korean population and 17 SNPs related to telomere length in Caucasians (15). Six of the 17 SNPs were associated with short telomere length in the Korean population and were selected for the present case-control study. Four SNPs in TERT (rs2853669, rs2736108, rs2736098, and rs7705526), one SNP in TERC (rs2293607), and one SNP in ZNF676 (rs412658) were genotyped. The rs7705526 polymorphism was genotyped by PCR-RFLP analysis, and the others were genotyped by melting-curve analysis using fluorescence-labeled hybridization probes (LightCycler 480, Roche Diagnostic, Mannheim, Germany). Samples that could not be scored by the LightCycler were re-genotyped by direct sequencing using an ABI 3730xl sequencer (Invitrogen, Garlsbad, CA, USA). The call rates for the SNPs were more than 95% (Supplementary Table 1). All genotyping analyses were blinded with respect to the case or control status to ensure quality control.
Statistical analysis
Comparisons between cases and controls were achieved by the Student t-test for continuous variables or the chi-square test for categorical variables. Hardy-Weinberg equilibrium was assessed by a goodness-of-fit chi-square test. To estimate lung cancer risk related to the SNPs, multivariate logistic regression analysis was used to calculate the adjusted odds ratio (aORs) and 95% confidence intervals (CIs) after adjusting for age, gender, and pack-years of smoking. For stratified analysis, individuals were grouped according to age (<62 yr vs. ≥62 yr), gender (male vs. female), smoking status (never vs. ever), histologic subtype (squamous cell carcinoma vs. adenocarcinoma vs. small cell carcinoma), and stage (I or II vs. III or IV). To compare aORs in each subgroup, homogeneity was tested using the Wald method. Statistical analyses were performed using the SPSS 21.0 software (SPSS Inc., Chicago, IL, USA).
Ethics statement
Written informed consent was obtained from each subject. This study protocol was reviewed and approved by the institutional review board of Kyungpook National University Hospital (KNUH 2012-01-011-004).
RESULTS
The demographics of the cases and controls are shown in Table 1. Significant differences in smoking status and pack-years of smoking were observed between cases and controls (P<0.001). Therefore, these differences were adjusted in the multivariate analyses. Basic information for the 6 SNPs, including gene locations and minor allele frequencies are shown in Supplementary Table 1. The genotype frequencies of the 6 SNPs were in Hardy-Weinberg equilibrium.
Table 1. Characteristics of the study population.
| Variable | Cases (n = 1,100) | Controls (n = 1,096) | P |
|---|---|---|---|
| Age (yr) | |||
| Mean ± SD | 61.4 ± 8.7 | 60.8 ± 7.4 | 0.12* |
| Gender | |||
| Male | 738 (67.1) | 736 (67.2) | 0.98† |
| Female | 362 (32.9) | 360 (32.8) | |
| Smoking status | |||
| Current | 520 (47.3) | 415 (37.9) | < 0.001† |
| Former | 267 (24.3) | 287 (26.2) | |
| Never | 313 (28.5) | 394 (35.9) | |
| Pack-years‡ | |||
| Mean ± SD | 38.7 ± 20.4 | 30.5 ± 17.4 | < 0.001* |
| Histologic type | |||
| Squamous cell carcinoma | 404 (36.7) | ||
| Adenocarcinoma | 507 (46.1) | ||
| Large cell carcinoma | 21 (1.9) | ||
| Small cell lung cancer | 148 (13.5) | ||
| NSCLC-NOS | 20 (1.8) | ||
| Stage | |||
| I | 236 (21.5) | ||
| II | 67 (6.1) | ||
| III | 361 (32.8) | ||
| IV | 436 (39.6) |
Numbers in parenthesis, column percentage. *t-test; †chi-square test; ‡in current and former smokers. SD, standard deviation; NSCLC-NOS, Non-small cell lung cancer, not otherwise specified.
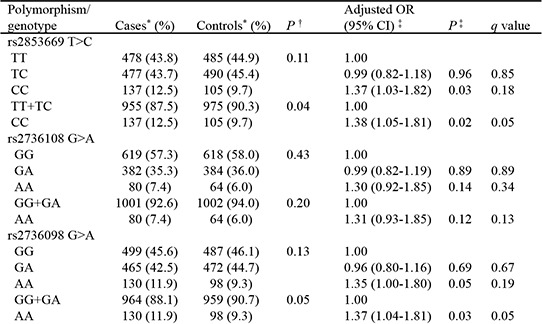
The genotype distributions of the 6 SNPs in the cases and controls are shown in Table 2. Among the 6 SNPs, the genotype frequencies of TERT rs2853669 and rs2736098 differed significantly between cases and controls. After adjusting for age, gender, and pack-years of smoking, individuals carrying the rs2853669 CC genotype had a significantly increased risk of lung cancer compared to those with the rs2853669 TT or TC genotypes (aOR=1.38, 95% CI=1.05-1.81, P=0.02). The rs2736098 G>A genotype was also significantly associated with lung cancer risk under a recessive model (aOR=1.37, 95% CI=1.04-1.81, P=0.03).
Table 2. Genotype frequencies of polymorphisms in lung cancer cases and controls.
| Polymorphism/genotype | Cases* (%) | Controls* (%) | P† | Adjusted OR (95% CI)‡ | P‡ | q value |
|---|---|---|---|---|---|---|
| rs2853669 T > C | ||||||
| TT | 478 (43.8) | 485 (44.9) | 0.11 | 1.00 | ||
| TC | 477 (43.7) | 490 (45.4) | 0.99 (0.82-1.18) | 0.96 | 0.85 | |
| CC | 137 (12.5) | 105 (9.7) | 1.37 (1.03-1.83) | 0.03 | 0.18 | |
| TT+TC | 955 (87.5) | 975 (90.3) | 0.04 | 1.00 | ||
| CC | 137 (12.5) | 105 (9.7) | 1.38 (1.05-1.81) | 0.02 | 0.05 | |
| rs2736108 G > A | ||||||
| GG | 619 (57.3) | 618 (58.0) | 0.43 | 1.00 | ||
| GA | 382 (35.3) | 384 (36.0) | 0.99 (0.82-1.19) | 0.89 | 0.89 | |
| AA | 80 (7.4) | 64 (6.0) | 1.30 (0.92-1.85) | 0.14 | 0.34 | |
| GG+GA | 1001 (92.6) | 1002 (94.0) | 0.20 | 1.00 | ||
| AA | 80 (7.4) | 64 (6.0) | 1.31 (0.93-1.85) | 0.12 | 0.13 | |
| rs2736098 G > A | ||||||
| GG | 499 (45.6) | 487 (46.1) | 0.13 | 1.00 | ||
| GA | 465 (42.5) | 472 (44.7) | 0.96 (0.80-1.16) | 0.69 | 0.67 | |
| AA | 130 (11.9) | 98 (9.3) | 1.35 (1.00-1.80) | 0.05 | 0.19 | |
| GG+GA | 964 (88.1) | 959 (90.7) | 0.05 | 1.00 | ||
| AA | 130 (11.9) | 98 (9.3) | 1.37 (1.04-1.81) | 0.03 | 0.05 | |
| rs7705526 C > A | ||||||
| CC | 418 (39.6) | 433 (41.1) | 0.64 | 1.00 | ||
| CA | 474 (44.9) | 470 (44.6) | 1.05 (0.87-1.27) | 0.62 | 0.66 | |
| AA | 164 (15.5) | 150 (14.2) | 1.16 (0.89-1.50) | 0.28 | 0.54 | |
| CC+CA | 892 (84.5) | 903 (85.8) | 0.41 | 1.00 | ||
| AA | 164 (15.5) | 150 (14.2) | 1.13 (0.89-1.44) | 0.33 | 0.33 | |
| rs2293607 G > A | ||||||
| GG | 401 (37.3) | 406 (38.6) | 0.82 | 1.00 | ||
| GA | 514 (47.8) | 491 (46.6) | 1.06 (0.88-1.28) | 0.53 | 0.65 | |
| AA | 161 (15.0) | 156 (14.8) | 1.07 (0.82-1.39) | 0.63 | 0.66 | |
| GG+GA | 915 (85.0) | 897 (85.2) | 0.92 | 1.00 | ||
| AA | 161 (15.0) | 156 (14.8) | 1.03 (0.81-1.31) | 0.80 | 0.80 | |
| rs412658 C > T | ||||||
| CC | 400 (38.2) | 401 (38.4) | 0.46 | 1.00 | ||
| CT | 500 (47.8) | 478 (45.8) | 1.07 (0.89-1.29) | 0.48 | 0.65 | |
| TT | 146 (14.0) | 164 (15.7) | 0.91 (0.70-1.19) | 0.51 | 0.65 | |
| CC+CT | 900 (86.0) | 879 (84.3) | 0.26 | 1.00 | ||
| TT | 146 (14.0) | 164 (15.7) | 0.88 (0.69-1.12) | 0.31 | 0.31 |
*Numbers may not add up total number of patients due to genotype failures; †P value for chi-square test; ‡ORs, 95% CIs and their corresponding P values were calculated using multivariate regression analysis, adjusted for age, gender, and pack-years smoking. OR, odds ratio; CI, confidence interval.
The TERT rs2853669 and rs2736098 polymorphisms were in strong linkage disequilibrium (LD) (|D'|=0.95, Supplementary Fig. 1). The rs2853669 polymorphism is known to be a potentially functional polymorphism (16). Therefore, we examined the association between TERT rs2853669 and lung cancer risk after stratifying the subjects by age, gender, smoking status, histologic subtype, and stage (Table 3). When stratified according to age, the effect of rs2853669 on lung cancer risk was significant in younger individuals (aOR=1.73, 95% CI=1.18-2.54, P=0.005) but not in older individuals (aOR=1.08, 95% CI=0.74-1.59, P=0.68; P value for homogeneity test [PH]=0.09). When stratified according to histologic subtype, significant association between rs2853669 and lung cancer risk was observed in adenocarcinoma (AC) (aOR=1.50, 95% CI=1.07-2.07, P=0.02) but not in squamous cell carcinoma (SCC) or small cell lung cancer (SCLC) (aOR=1.16, 95% CI=0.78-1.73, P=0.46; aOR=1.18, 95% CI=0.66-2.11, P=0.57, respectively).
Table 3. Association between TERT rs2853669 and lung cancer risk according to age, gender, smoking status, tumor histology, and stage.
| Variables | Cases | Controls | P† | Adjusted OR (95% CI)* | |||||
|---|---|---|---|---|---|---|---|---|---|
| TT or TC | CC | TT or TC | CC | TT or TC | CC | P | PH‡ | ||
| Age (yr) | |||||||||
| <62 | 424 (85.5) | 72 (14.5) | 500 (90.9) | 50 (9.1) | 0.01 | 1.00 | 1.73 (1.18-2.54) | 0.005§ | 0.09 |
| ≥ 62 | 531 (59.1) | 65 (10.9) | 475 (89.6) | 55 (10.4) | 0.77 | 1.00 | 1.08 (0.74-1.59) | 0.68§ | |
| Gender | |||||||||
| Male | 643 (87.8) | 89 (12.2) | 662 (90.7) | 68 (9.3) | 0.08 | 1.00 | 1.33 (0.95-1.87) | 0.09∥ | 0.79 |
| Female | 312 (86.7) | 48 (13.3) | 313 (89.4) | 37 (10.6) | 0.26 | 1.44 (0.91-2.29) | 0.12∥ | ||
| Smoking status | |||||||||
| Never | 264 (84.9) | 47 (15.1) | 344 (89.6) | 40 (10.4) | 0.06 | 1.00 | 1.47 (0.93-2.32) | 0.10¶ | 0.69 |
| Ever | 691 (88.5) | 90 (11.5) | 631 (90.7) | 65 (9.3) | 0.17 | 1.00 | 1.31 (0.94-1.84) | 0.11¶ | |
| Histology | |||||||||
| SCC | 356 (89.2) | 43 (10.8) | 975 (90.3) | 105 (9.7) | 0.55 | 1.00 | 1.16 (0.78-1.73) | 0.46** | 0.33 |
| AC | 433 (85.9) | 71 (14.1) | 975 (90.3) | 105 (9.7) | 0.01 | 1.00 | 1.50 (1.07-2.07) | 0.02** | |
| SCLC | 132 (89.2) | 16 (10.8) | 975 (90.3) | 105 (9.7) | 0.68 | 1.00 | 1.18 (0.66-2.11) | 0.57** | 0.50 |
| Stage | |||||||||
| I, II | 259 (86.9) | 39 (13.1) | 975 (90.3) | 105 (9.7) | 0.09 | 1.00 | 1.39 (0.94-2.06) | 0.10** | 0.98 |
| III, IV | 696 (87.7) | 98 (12.3) | 975 (90.3) | 105 (9.7) | 0.07 | 1.00 | 1.38 (1.03-1.86) | 0.03** | |
*ORs and 95% CIs and their corresponding P-value were calculated by unconditional logistic regression analysis; †P value for chi-square test; ‡P value for homogeneity test; §Adjusted for gender and pack-years of smoking; ∥Adjusted for age and pack-years of smoking; ¶Adjusted for age and gender; **Adjusted for age, gender, and pack-years of smoking. SCC, squamous cell carcinoma; AC, adenocarcinoma; SCLC, small cell lung cancer; OR, odds ratio; CI, confidence interval.
We conducted interaction analysis between rs2853669 and smoking status. When non-smokers with rs2853669 TT or TC genotype was considered as reference group, smokers with rs2853669 CC genotype had a significantly increased risk of lung cancer (OR=3.71, 95% CI=2.35-5.86, P=1.9×10-8, Supplementary Table 2).
DISCUSSION
In this study, we investigated the association between 6 SNPs related to short telomere length in the Korean population and lung cancer risk via a case-control study. Among the 6 SNPs, two SNPs (rs2853669 and rs2736098) in TERT were significantly associated with increased lung cancer risk in the Korean population.
TERT is one of the major components of telomerase (17,18). Telomerase is a reverse transcriptase that can add nucleotide sequence to telomere ends, thus protecting telomere length from progressive shortening (19). Polymorphisms in TERT can influence telomere length (16,20,21) and can also affect cancer susceptibility (20,22,23). In the present study, the TERT rs2736098 AA genotype had an increased risk of lung cancer compared with the TERT rs2736098 GG or GA genotypes. The rs2736098 polymorphism is located in exon 2 of TERT and is a synonymous polymorphism (A305A). In the present study, rs2736098 was strongly linked to rs2853669 (|D'|=0.95), which is located in the promoter region of TERT. TERT rs2853669 is known to be a potentially functional polymorphism (16). Therefore, the effect of rs2736098 on lung cancer risk may be due to LD with rs2853669. In the present study, individuals that had the TERT rs2853669 CC genotype had a significantly increased risk of lung cancer compared with those that had the rs2853669 TT or TC genotypes. In the previous study, relative telomere length of individual with rs2853669 CC genotype was significantly shorter than those with rs2853669 TT or TC genotype (1.97±0.6 vs. 2.37±0.6, P=0.03) (15). Increased risk of cancer in individual with short telomere is biologically plausible, especially in light of the function of telomere. Main function of telomere is to keep genomic integrity and stability from deterioration or atypical fusion of chromosome (2). Therefore, the shortened telomere can lead to chromosomal instability and may thereby increase cancer risk. Recently, it was reported that the TERT rs2853669 C allele was associated with an increased risk of lung cancer in a Chinese population (23). That the results from these two independent studies are consistent is strong evidence that TERT rs2853669 is associated with lung cancer risk.
When stratified according to histologic subtype, TERT rs2853669 was associated with lung cancer risk in AC but not in SCC or SCLC. Zhong et al. (23) also reported that increased risk of lung cancer due to a variant of rs2853669 was more prominent in AC. Although AC and SCC are categorized as NSCLC, AC has a different molecular pattern, carcinogenesis, and treatment response from SCC (24,25,26). Differential expression of TERT between lung AC and SCC or SCLC was also reported in a previous study (27). Therefore, TERT rs2853669 may influence lung cancer susceptibility differently according to histologic subtype. Additional studies are required to confirm this finding.
Interestingly, the effect of TERT rs2853669 on lung cancer risk was different by age. TERT rs2853669 was associated with a significantly increased risk of lung cancer in younger individuals but not in older individuals; this difference showed a borderline significance (PH=0.09). This suggests that short telomere length due to TERT rs2853669 could play a predominant role in the development of lung cancer at earlier ages.
Among the 6 SNPs related to short telomere length in the Korean population, only two SNPs (TERT rs2736098 and rs2853669) were associated with lung cancer risk. The TERT rs2736108 AA genotype tended to associate with increased lung cancer risk, but the association was not statistically significant. TERT rs7705526, TERC rs2293607, and ZNF676 rs412658 were not associated with lung cancer risk. Since carcinogenesis is a multistep process (28), multiple genetic and epigenetic changes are required to transform a normal cell into a cancer cell (29). A mouse model with shortened telomeres and mutant p53 showed increased hepatocellular carcinoma progression, while a model with shortened telomeres and functional p53 did not (30). Therefore, a single nucleotide change that influences telomere length may not have sufficient power to affect lung cancer risk. Additionally, a polymorphism that causes short telomere length may influence lung cancer risk separately from telomere length, and so the increased risk of lung cancer due to short telomere length can be weakened by another biologic effect of the polymorphism.
In summary, we previously investigated the associations between lung cancer risk and 17 SNPs related to telomere length in Caucasians. We identified 6 SNPs associated with short telomere length in the Korean population. Among these, TERT rs2853669 and rs2736098 were associated with increased lung cancer risk in the present study. This suggests that a common functional promoter polymorphism, TERT rs2853669, may contribute to both telomere length and lung cancer risk in the Korean population.
Footnotes
Funding: This work was supported by the R&D program of MKE/KEIT (10040393, Development and commercialization of molecular diagnostic technologies for lung cancer through clinical validation).
DISCLOSURE: The authors have no potential conflicts of interest to disclose.
AUTHOR CONTRIBUTION: Conception and design of the study: Yoo SS, Do SK, Park JY. Acquisition of data: Yoo SS, Lee SY, Lee J, Cha SI, Kim CH, Park JY. Analyzed the data: Do SK, Choi JE. First draft of manuscript: Yoo SS, Do SK, Park JY. Revision and critical review of the manuscript: Yoo SS, Do SK, Choi JE, Lee J, Cha SI, Kim CH, Park JY. Criteria for authorship and accepting final manuscript: all authors.
Supplementary Materials
Supplementary Table 1. Locations and minor allele frequencies of polymorphisms.
| Polymorphism | Gene | Region | Minor allele | Call rates (%) | MAF | |||
|---|---|---|---|---|---|---|---|---|
| Cases | Controls | Korean | JPT* | CEU* | ||||
| rs2853669 | TERT | Promoter | C | 99.3 | 98.5 | 0.37 | - | 0.38 |
| rs2736108 | TERT | Promoter | A | 98.3 | 97.3 | 0.26 | 0.17 | 0.35 |
| rs2736098 | TERT | Exon2 | A | 99.5 | 96.4 | 0.37 | 0.25 | 0.37 |
| rs7705526 | TERT | Intron | A | 96.0 | 96.1 | 0.36 | - | 0.50 |
| rs2293607 | TERC | 3' UTR | A | 97.8 | 96.1 | 0.40 | 0.33 | 0.75 |
| rs412658 | ZNF676 | 3' UTR | T | 95.1 | 95.2 | 0.44 | 0.33 | 0.33 |
*Data from NCBI (http://www.ncbi.nlm.nih.gov/SNP). MAF, minor allele frequency; UTR, untranslated region.
Supplementary Table 2. Stratified and interaction analysis between rs2853669 and smoking status associated with lung cancer risk.
| Genotype | Smoking status | |||||
|---|---|---|---|---|---|---|
| Nonsmokers | Smokers | |||||
| Cases/Controls | OR (95% CI)* | P | Cases/Controls | OR (95% CI)* | P | |
| TT + TC | 264/344 | Reference | 691/631 | 2.82 (2.02-3.94) | 1.1 × 10-9 | |
| CC | 47/40 | 1.49 (0.94-2.35) | 0.09 | 90/65 | 3.71 (2.35-5.86) | 1.9 × 10-8 |
| Pmult, Pmult† | 4.1 × 10-9, 2.4 × 10-8 | |||||
| Padd, Padd† | 0.02, 0.14 | |||||
*ORs and 95% CI were calculated by unconditional logistic regression after adjusting for sex and age; †P was modified by bonferroni correction for multiple comparisons. OR, odds ratio; CI, confidence interval; Pmult, P value for multiplicative interaction term; Padd, P value for additive interaction model.
Reconstructed LD plots of 4 polymorphisms in TERT using the study population. The plots were generated using the Haploview program and the method proposed by Gabriel et al. (31). The numbers in the squares are |D'| (×100) values. The triangle indicates haplotype block.
References
- 1.Blasco MA. Telomeres and human disease: ageing, cancer and beyond. Nat Rev Genet. 2005;6:611–622. doi: 10.1038/nrg1656. [DOI] [PubMed] [Google Scholar]
- 2.Campisi J, Kim SH, Lim CS, Rubio M. Cellular senescence, cancer and aging: the telomere connection. Exp Gerontol. 2001;36:1619–1637. doi: 10.1016/s0531-5565(01)00160-7. [DOI] [PubMed] [Google Scholar]
- 3.Gilley D, Tanaka H, Herbert BS. Telomere dysfunction in aging and cancer. Int J Biochem Cell Biol. 2005;37:1000–1013. doi: 10.1016/j.biocel.2004.09.003. [DOI] [PubMed] [Google Scholar]
- 4.Alter BP, Giri N, Savage SA, Rosenberg PS. Cancer in dyskeratosis congenita. Blood. 2009;113:6549–6557. doi: 10.1182/blood-2008-12-192880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Brouilette SW, Moore JS, McMahon AD, Thompson JR, Ford I, Shepherd J, Packard CJ, Samani NJ West of Scotland Coronary Prevention Study Group. Telomere length, risk of coronary heart disease, and statin treatment in the West of Scotland Primary Prevention Study: a nested case-control study. Lancet. 2007;369:107–114. doi: 10.1016/S0140-6736(07)60071-3. [DOI] [PubMed] [Google Scholar]
- 6.Cawthon RM, Smith KR, O'Brien E, Sivatchenko A, Kerber RA. Association between telomere length in blood and mortality in people aged 60 years or older. Lancet. 2003;361:393–395. doi: 10.1016/S0140-6736(03)12384-7. [DOI] [PubMed] [Google Scholar]
- 7.Willeit P, Willeit J, Mayr A, Weger S, Oberhollenzer F, Brandstätter A, Kronenberg F, Kiechl S. Telomere length and risk of incident cancer and cancer mortality. JAMA. 2010;304:69–75. doi: 10.1001/jama.2010.897. [DOI] [PubMed] [Google Scholar]
- 8.Wu X, Amos CI, Zhu Y, Zhao H, Grossman BH, Shay JW, Luo S, Hong WK, Spitz MR. Telomere dysfunction: a potential cancer predisposition factor. J Natl Cancer Inst. 2003;95:1211–1218. doi: 10.1093/jnci/djg011. [DOI] [PubMed] [Google Scholar]
- 9.Ma H, Zhou Z, Wei S, Liu Z, Pooley KA, Dunning AM, Svenson U, Roos G, Hosgood HD, 3rd, Shen M, et al. Shortened telomere length is associated with increased risk of cancer: a meta-analysis. PLoS One. 2011;6:e20466. doi: 10.1371/journal.pone.0020466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jang JS, Choi YY, Lee WK, Choi JE, Cha SI, Kim YJ, Kim CH, Kam S, Jung TH, Park JY. Telomere length and the risk of lung cancer. Cancer Sci. 2008;99:1385–1389. doi: 10.1111/j.1349-7006.2008.00831.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Prescott J, Kraft P, Chasman DI, Savage SA, Mirabello L, Berndt SI, Weissfeld JL, Han J, Hayes RB, Chanock SJ, et al. Genome-wide association study of relative telomere length. PLoS One. 2011;6:e19635. doi: 10.1371/journal.pone.0019635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Codd V, Mangino M, van der Harst P, Braund PS, Kaiser M, Beveridge AJ, Rafelt S, Moore J, Nelson C, Soranzo N, et al. Wellcome Trust Case Control Consortium. Common variants near TERC are associated with mean telomere length. Nat Genet. 2010;42:197–199. doi: 10.1038/ng.532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kote-Jarai Z, Saunders EJ, Leongamornlert DA, Tymrakiewicz M, Dadaev T, Jugurnauth-Little S, Ross-Adams H, Al Olama AA, Benlloch S, Halim S, et al. COGS-CRUK GWAS-ELLIPSE (Part of GAME-ON) Initiative; UK Genetic Prostate Cancer Study Collaborators/British Association of Urological Surgeons' Section of Oncology; UK ProtecT Study Collaborators; PRACTICAL Consortium. Fine-mapping identifies multiple prostate cancer risk loci at 5p15, one of which associates with TERT expression. Hum Mol Genet. 2013;22:2520–2528. doi: 10.1093/hmg/ddt086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Mangino M, Hwang SJ, Spector TD, Hunt SC, Kimura M, Fitzpatrick AL, Christiansen L, Petersen I, Elbers CC, Harris T, et al. Genome-wide meta-analysis points to CTC1 and ZNF676 as genes regulating telomere homeostasis in humans. Hum Mol Genet. 2012;21:5385–5394. doi: 10.1093/hmg/dds382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Do SK, Yoo SS, Choi YY, Choi JE, Jeon H, Lee WK, Lee SY, Lee J, Cha SI, Kim CH, et al. Replication of the results of genome-wide and candidate gene association studies on telomere length in Korean population. Korean J Intern Med. 2015;30:719–726. doi: 10.3904/kjim.2015.30.5.719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hsu CP, Hsu NY, Lee LW, Ko JL. Ets2 binding site single nucleotide polymorphism at the hTERT gene promoter--effect on telomerase expression and telomere length maintenance in non-small cell lung cancer. Eur J Cancer. 2006;42:1466–1474. doi: 10.1016/j.ejca.2006.02.014. [DOI] [PubMed] [Google Scholar]
- 17.Cohen SB, Graham ME, Lovrecz GO, Bache N, Robinson PJ, Reddel RR. Protein composition of catalytically active human telomerase from immortal cells. Science. 2007;315:1850–1853. doi: 10.1126/science.1138596. [DOI] [PubMed] [Google Scholar]
- 18.Autexier C, Lue NF. The structure and function of telomerase reverse transcriptase. Annu Rev Biochem. 2006;75:493–517. doi: 10.1146/annurev.biochem.75.103004.142412. [DOI] [PubMed] [Google Scholar]
- 19.Chan SR, Blackburn EH. Telomeres and telomerase. Philos Trans R Soc Lond B Biol Sci. 2004;359:109–121. doi: 10.1098/rstb.2003.1370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bojesen SE, Pooley KA, Johnatty SE, Beesley J, Michailidou K, Tyrer JP, Edwards SL, Pickett HA, Shen HC, Smart CE, et al. Multiple independent variants at the TERT locus are associated with telomere length and risks of breast and ovarian cancer. Nat Genet. 2013;45:371–384. 384e1–384e2. doi: 10.1038/ng.2566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Matsubara Y, Murata M, Yoshida T, Watanabe K, Saito I, Miyaki K, Omae K, Ikeda Y. Telomere length of normal leukocytes is affected by a functional polymorphism of hTERT. Biochem Biophys Res Commun. 2006;341:128–131. doi: 10.1016/j.bbrc.2005.12.163. [DOI] [PubMed] [Google Scholar]
- 22.Wu H, Qiao N, Wang Y, Jiang M, Wang S, Wang C, Hu L. Association between the telomerase reverse transcriptase (TERT) rs2736098 polymorphism and cancer risk: evidence from a case-control study of non-small-cell lung cancer and a meta-analysis. PLoS One. 2013;8:e76372. doi: 10.1371/journal.pone.0076372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhong R, Liu L, Zou L, Zhu Y, Chen W, Zhu B, Shen N, Rui R, Long L, Ke J, et al. Genetic variations in TERT-CLPTM1L locus are associated with risk of lung cancer in Chinese population. Mol Carcinog. 2013;52:E118–E126. doi: 10.1002/mc.22043. [DOI] [PubMed] [Google Scholar]
- 24.Perez-Moreno P, Brambilla E, Thomas R, Soria JC. Squamous cell carcinoma of the lung: molecular subtypes and therapeutic opportunities. Clin Cancer Res. 2012;18:2443–2451. doi: 10.1158/1078-0432.CCR-11-2370. [DOI] [PubMed] [Google Scholar]
- 25.Molina-Pinelo S, Gutiérrez G, Pastor MD, Hergueta M, Moreno-Bueno G, Garcia-Carbonero R, Nogal A, Suárez R, Salinas A, Pozo-Rodriguez F, et al. MicroRNA-dependent regulation of transcription in non-small cell lung cancer. PLoS One. 2014;9:e90524. doi: 10.1371/journal.pone.0090524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Scagliotti GV, Parikh P, von Pawel J, Biesma B, Vansteenkiste J, Manegold C, Serwatowski P, Gatzemeier U, Digumarti R, Zukin M, et al. Phase III study comparing cisplatin plus gemcitabine with cisplatin plus pemetrexed in chemotherapy-naive patients with advanced-stage non-small-cell lung cancer. J Clin Oncol. 2008;26:3543–3551. doi: 10.1200/JCO.2007.15.0375. [DOI] [PubMed] [Google Scholar]
- 27.Lantuejoul S, Soria JC, Moro-Sibilot D, Morat L, Veyrenc S, Lorimier P, Brichon PY, Sabatier L, Brambilla C, Brambilla E. Differential expression of telomerase reverse transcriptase (hTERT) in lung tumours. Br J Cancer. 2004;90:1222–1229. doi: 10.1038/sj.bjc.6601643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Barrett JC. Mechanisms of multistep carcinogenesis and carcinogen risk assessment. Environ Health Perspect. 1993;100:9–20. doi: 10.1289/ehp.931009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Blanco D, Vicent S, Fraga MF, Fernandez-Garcia I, Freire J, Lujambio A, Esteller M, Ortiz-de-Solorzano C, Pio R, Lecanda F, et al. Molecular analysis of a multistep lung cancer model induced by chronic inflammation reveals epigenetic regulation of p16 and activation of the DNA damage response pathway. Neoplasia. 2007;9:840–852. doi: 10.1593/neo.07517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Farazi PA, Glickman J, Horner J, Depinho RA. Cooperative interactions of p53 mutation, telomere dysfunction, and chronic liver damage in hepatocellular carcinoma progression. Cancer Res. 2006;66:4766–4773. doi: 10.1158/0008-5472.CAN-05-4608. [DOI] [PubMed] [Google Scholar]
- 31.Gabriel SB, Schaffner SF, Nguyen H, Moore JM, Roy J, Blumenstiel B, Higgins J, DeFelice M, Lochner A, Faggart M, et al. The structure of haplotype blocks in the human genome. Science. 2002;296:2225–2229. doi: 10.1126/science.1069424. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Table 1. Locations and minor allele frequencies of polymorphisms.
| Polymorphism | Gene | Region | Minor allele | Call rates (%) | MAF | |||
|---|---|---|---|---|---|---|---|---|
| Cases | Controls | Korean | JPT* | CEU* | ||||
| rs2853669 | TERT | Promoter | C | 99.3 | 98.5 | 0.37 | - | 0.38 |
| rs2736108 | TERT | Promoter | A | 98.3 | 97.3 | 0.26 | 0.17 | 0.35 |
| rs2736098 | TERT | Exon2 | A | 99.5 | 96.4 | 0.37 | 0.25 | 0.37 |
| rs7705526 | TERT | Intron | A | 96.0 | 96.1 | 0.36 | - | 0.50 |
| rs2293607 | TERC | 3' UTR | A | 97.8 | 96.1 | 0.40 | 0.33 | 0.75 |
| rs412658 | ZNF676 | 3' UTR | T | 95.1 | 95.2 | 0.44 | 0.33 | 0.33 |
*Data from NCBI (http://www.ncbi.nlm.nih.gov/SNP). MAF, minor allele frequency; UTR, untranslated region.
Supplementary Table 2. Stratified and interaction analysis between rs2853669 and smoking status associated with lung cancer risk.
| Genotype | Smoking status | |||||
|---|---|---|---|---|---|---|
| Nonsmokers | Smokers | |||||
| Cases/Controls | OR (95% CI)* | P | Cases/Controls | OR (95% CI)* | P | |
| TT + TC | 264/344 | Reference | 691/631 | 2.82 (2.02-3.94) | 1.1 × 10-9 | |
| CC | 47/40 | 1.49 (0.94-2.35) | 0.09 | 90/65 | 3.71 (2.35-5.86) | 1.9 × 10-8 |
| Pmult, Pmult† | 4.1 × 10-9, 2.4 × 10-8 | |||||
| Padd, Padd† | 0.02, 0.14 | |||||
*ORs and 95% CI were calculated by unconditional logistic regression after adjusting for sex and age; †P was modified by bonferroni correction for multiple comparisons. OR, odds ratio; CI, confidence interval; Pmult, P value for multiplicative interaction term; Padd, P value for additive interaction model.
Reconstructed LD plots of 4 polymorphisms in TERT using the study population. The plots were generated using the Haploview program and the method proposed by Gabriel et al. (31). The numbers in the squares are |D'| (×100) values. The triangle indicates haplotype block.