
FIGURE 1.
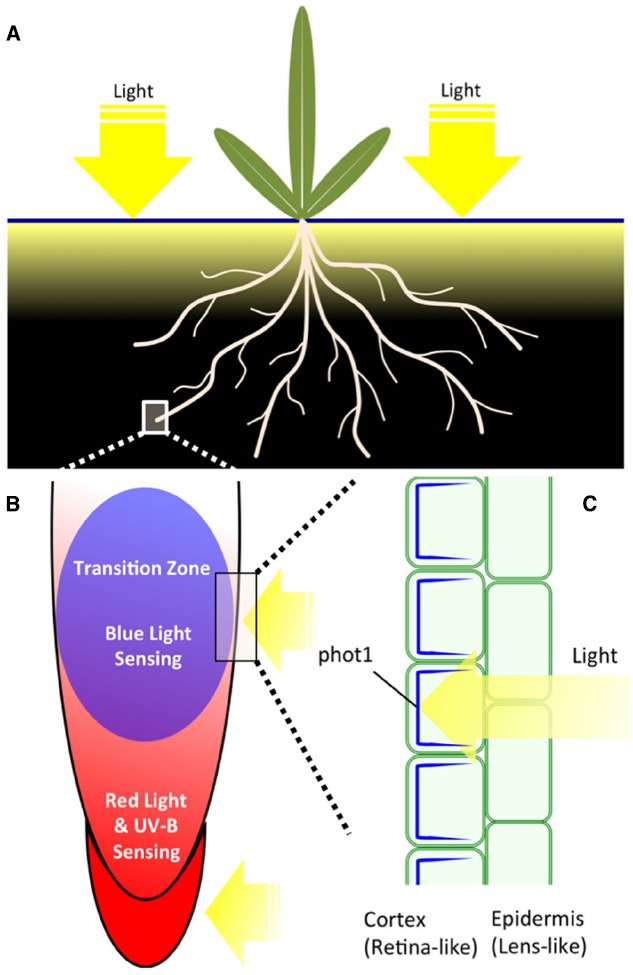
(A) Plant organs and their light environment. Shoot part of plants is fully exposed to light during a day. Root part is exposed only to limited amounts of light which penetrates into the soil during a day. Actual light mosaics in the soil depend on numerous factors and it changes with the depth (Woolley and Stoller, 1978; Tester and Morris, 1987; Kasperbauer and Hunt, 1988). (B) Root apex zonation with respect of light-sensitivity. Root cap, meristem and transition zone are expressing phytochromes (Adam et al., 1994; Somers and Quail, 1995a,b; Goosey et al., 1997) whereas only the transition zone is abundantly expressing phototropin phot1 (Wan et al., 2008, 2012). UVR8 is expressed, similarly as phytochromes, in all zones of Arabidopsis root apex (Rizzini et al., 2011; Yokawa et al., 2014). RUS1 and RUS2 are also expressed preferentially in cells of the transition zone (Leasure et al., 2009; Yu et al., 2013). (C) Tissue-specific and polar distribution of phot1 in cells of the transition zone. While epidermis cells do not express phot1, this blue light photoreceptor essential for negative phototropism of roots is abundant and polarly distributed (shown in blue) in underlying cortex cells (Wan et al., 2008) and controls PIN2 distribution and recycling (Wan et al., 2012). This tissue-specific expression and polarity of phot1 fits nicely to the plant “ocelli” concept (the epidermis act as lens-like tissue and the sub-epidermis as retina-like tissue) as proposed by Haberland for shoots (Haberlandt, 1904; Darwin, 1907; von Guttenberg, 1955).