

Abstract
The regulatory mechanisms of cytoplasmic Ca2+ after myocardial infarction-induced Ca2+ overload involve secretory pathway Ca2+-ATPase 1 and the Golgi apparatus and are well understood. However, the effect of Golgi apparatus on Ca2+ overload after cerebral ischemia and reperfusion remains unclear. Four-vessel occlusion rats were used as animal models of cerebral ischemia. The expression of secretory pathway Ca2+-ATPase 1 in the cortex and hippocampus was detected by immunoblotting, and Ca2+ concentrations in the cytoplasm and Golgi vesicles were determined. Results showed an overload of cytoplasmic Ca2+ during ischemia and reperfusion that reached a peak after reperfusion. Levels of Golgi Ca2+ showed an opposite effect. The expression of Golgi-specific secretory pathway Ca2+-ATPase 1 in the cortex and hippocampus decreased before ischemia and reperfusion, and increased after reperfusion for 6 hours. This variation was similar to the alteration of calcium in separated Golgi vesicles. These results indicate that the Golgi apparatus participates in the formation and alleviation of calcium overload, and that secretory pathway Ca2+-ATPase 1 tightly responds to ischemia and reperfusion in nerve cells. Thus, we concluded that secretory pathway Ca2+-ATPase 1 plays an essential role in cytosolic calcium regulation and its expression can be used as a marker of Golgi stress, responding to cerebral ischemia and reperfusion. The secretory pathway Ca2+-ATPase 1 can be an important neuroprotective target of ischemic stroke.
Keywords: nerve regeneration, brain injury, global cerebral ischemia, Golgi apparatus, Golgi stress, cytoplasmic Ca2+, homeostasis, Golgi Ca2+, Ca2+ pump, secretory pathway Ca2+-ATPase 1, neural protection, NSFC grant, neural regeneration
Introduction
The function of the Golgi apparatus in secretion and protein modification has long been known and is one of the earliest discoveries in the cell. These functions are important for biochemical reactions in neuronal cells, but it was not known whether the Golgi apparatus also played a role in cell signaling. In the early 1990s, experiments using electron-probe and fluorescence-marked confocal microscopy indicated that a high concentration of Ca2+ was present in the Golgi apparatus (Chandra et al., 1991; Xue et al., 1994; Pezzati et al., 1997). In recent years, IP3R, sarco/endoplasmic reticulum Ca2+-ATPase (SERCA) and secretory pathway Ca2+ ATPase (SPCA) have been demonstrated to sequentially localize on Golgi apparatus membranes and are involved in Golgi apparatus Ca2+ release and uptake (Wuytack et al., 2003; Missiaen et al., 2007). Various physiological and pathological implications induced by Golgi apparatus Ca2+ remodeling have been discovered and it is now widely accepted that the Golgi apparatus acts as an intracellular Ca2+ store and participates in the regulation of cytosolic Ca2+.
In neurons, as a Ca2+ store, the Golgi apparatus probably takes part in the modification of calcium signaling in several neuronal diseases, such as Alzheimer's disease, amyotrophic lateral sclerosis (ALS) and Huntington's disease (Fan et al., 2008). Lin et al. (2007) proved that CALNUC, a Ca2+ binding protein located in the Golgi apparatus, blocks the biosynthesis of beta-amyloid precursor protein. Experiments by Demestre and colleagues (2006) indicated that the alleviated role of ALS-IgG is related to improving Ca2+ concentration in endoplasmic reticulum and Golgi apparatus. Moreover, IP3R, a release factor of calcium on Golgi apparatus cisternae, is probably involved in the pathological process of Huntington's disease (Fan et al., 2008; McCue et al., 2012; Pressler et al., 2013; Wong et al., 2013; Kienzle et al., 2014). However, there have been few investigations into the role of calcium signaling regulated by Golgi apparatus in ischemia/reperfusion injury (IRI) (Shull et al., 2011; Vandecaetsbeek et al., 2011).
In 1982, Krino first elucidated the phenomenon of delayed neuronal death following ischemia in the gerbil hippocampus. Cell death at the site of ischemia and its penumbra is recognized as the predominant cause of cerebral IRI (Ferrer and Planas, 2003). It is well known that calcium overload is the vital signal for cell death and is also the prominent event in cerebral IRI (Uematsu et al., 1988). Although the Golgi apparatus, which is widely distributed in neurons, executes a vital role in Ca2+ signaling, it is not clear how their function in cytosolic Ca2+ regulation is involved in cerebral IRI. Thus, it is necessary to assess the role of Golgi apparatus in cytosolic Ca2+ regulation in cerebral IRI.
As previously reviewed (Li et al., 2013), there are several types of Ca2+ regulators present on the Golgi apparatus membrane and in the Golgi apparatus cisternae. These include calcium channels, calcium pumps, calcium binding proteins and other calcium regulating proteins. Among the Golgi apparatus Ca2+ regulators, SPCA, as a Ca2+ pump, is specifically located on the cisternae/network of trans-Golgi and secretory vesicles (Antebi and Fink, 1992; Durr et al., 1998). Its function is to maintain the relative Ca2+ homeostasis in trans-Golgi compartments and to participate in the intra-Golgi transportation (Lissandron, 2010; Micaroni and Mironov, 2010; Zak et al., 2011). Its role in Ca2+ regulation in the Golgi apparatus was elucidated unambiguously during clinical research of pathological mechanisms of Hailey-Hailey Disease and other diseases by inducing cell death (Hu et al., 2000; Sudbrak et al., 2000; Behne et al., 2003; Van Baelen, 2004). Thus, we hypothesized that SPCA1 in Ca2+ regulation should be the essential factor of Golgi apparatus stress in cerebral ischemia and reperfusion.
By using the four-vessel occlusion rat as a model, we detected the expression of SPCA1 and concentration of Ca2+ in cytoplasm and Golgi vesicles of nerve cells in the cortex of rat cerebrum. We also focused on the relationship between expression of SPCA1 and cytoplasmic/Golgi Ca2+ concentration during ischemia and reperfusion. Based on these determinations, we can accurately explain calcium alterations in Golgi apparatus stress related to cerebral IRI, which gives further insight into possible treatments for ischemic stroke.
Materials and Methods
Animals
One hundred and twenty healthy male Sprague-Dawley rats (Hunan Silaike Jingda Co., Animal license SCXK (Xiang) 2009-0004), weighing 200 ± 28 g and aged approximately 4 months were used in this study. All surgical procedures and postoperative care were performed in accordance with Guidelines of the Animal Care and Use Committee in Central South University in China.
Rats were divided into four groups: control, sham operation, ischemia and reperfusion. Five subgroups [3 hours (R3 h), 6 hours (R6 h), 24 hours (R24 h), 3 days (R3 d) and 7 days (R7 d)] were set in the reperfusion group (Li et al., 2012). Rats in the control group were fed at room temperature and had no treatments. Rats of the sham operation group were only subjected to the operation so as to expose the vertebral arteries and carotid arteries. Rats in the ischemia group were executed to occlude the carotid arteries for 20 minutes. The blood fluid of carotid arteries was recovered in the reperfusion sub groups at different time points.
Establishment of animal models
Rats from ischemia group and reperfusion groups were used to establish animal models according to the method of four-vessel occlusion supplied by Pulsineli et al. (1988). All rats were intraperitoneally anesthetized with hydro-chloro-aldehyde (10% w/v, 400 mg/kg). Subsequently, the first operation was carried out to assure complete occlusion of the vertebral arteries by electrocauterization through the alar foramina of the first cervical vertebra. Twenty-four hours later, the second operation was taken to control the occlusion and reperfusion of the parallel common carotid arteries. Animals from the ischemia group received four-vessel occlusion and were sacrificed immediately. Animals from the reperfusion groups received four-vessel occlusion, then reperfusion for 3, 6, 24 hours and 3, 7 days, and then were sacrificed as soon as possible. Whole brains of these rats in various groups and subgroups were peeled out immediately.
To verify model establishment, complete vertebral artery occlusion in 100% of the rats could be assured by direct visualization of the bony perimeter of the tunnel in which the vertebral arteries travel.
Fluorescence in situ detection of SPCA1
Five brain sections from different groups were fixed in 4% paraformaldehyde (w/v) for 72 hours. The temporal lobe and hippocampus were embedded in wax, sliced into coronal sections, and then subjected to immuno-detection. To detect SPCA1, fluorescent direct monoclonal antibody against SPCA1 was used: anti-SPCA1 (1:400; Sigma-Aldrich Inc., St. Louis, MO, USA). Direct immunostaining procedures in separated neuronal cells were performed as previously described in Short Protocol of Molecular Biology (Ausubel et al., 2008). Sections were incubated with direct fluorescent monoclonal antibody of SPCA1 in 10 mM phosphate buffer (mouse monoclonal, 1:400; pH 7.0) at 4°C overnight. A drop of buffer containing this antibody was coated on the specimen for 30 minutes at 37°C, followed by three washes in PBS (10 mM, pH 7.0) for 3 minutes. Immunofluorescence images were observed using a Leica DM 3000 microscope (Leica Microsystem, Wetzlar, Hassium, Germany).
Western blot assay of SPCA1
Five pieces of brains from different groups were homogenized in the high-speed homogenizer at 3,500 ×g to extract the total protein and to detect the protein of SPCA1 by western blot assay. The total extracted proteins were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. After western blot assay (Trans-blot SD semi-dry transfer cell, Bio-Rad), blots on nitrocellulose membrane were detected with mouse polyclonal antibodies against SPCA1 (Sigma-Aldrich Inc.) at 37°C for 3 hours in dilution 1:400. After being washed with 0.05% PBS-Tween, the membranes were incubated with a goat anti-mouse secondary antibody conjugated with horseradish peroxidase (1:3,000, Sigma-Aldrich Inc.) at 37°C for 1 hour and washed again with PBS-Tween. Last, the membranes were visualized with diaminobenzidine (BB0124, Bio Basic Canada Inc., Markham, Ontario, Canada) and detected by 2500R Doc Imaging System (Tannon Inc., Shanghai, China). β-Actin was used as a housekeeper reference (actin monoclonal antibody fetch from Beyotime Biotech Inc., Cat. No. AA128; Nantong, China). Five animals per experimental group/subgroup were analyzed. To reduce differences among animals, sample loading on sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and variability due to staining detection, western blot assay was performed for each reperfusion time point per animal in triplicate. The results were documented by gel imaging system (Tannon Inc., Shanghai, China).
Fluorescent spectrophotometer detection of Ca2+ in neuronal cytoplasm and Golgi vesicles
Nerve cell preparation
Five brain sections from different groups were used to separate neuronal cells to determine the calcium concentration of cytoplasm and Golgi vesicles. The nerve cell was prepared according to the method of Dildy et al. (1989) and Brethes et al. (1987), and Ca2+ in cytoplasm and Golgi vesicles was detected. The prepared rat brain was washed in 4°C pre-cold Hank's Balanced Salt Solution (Life Tech Inc., Carlsbad, CA, USA) to detach the meninx and was rewashed three times in deionized water. The brain was cut into tiny pieces and digested in 0.1% (w/v) trypsin solution for 10 minutes, and homogenized at 37°C for 30 minutes. The digestion was blocked by 4°C pre-cold Dulbecco's modified Eagle's medium (DMEM, Life Tech Inc.). The homogenate was centrifuged at 2,000 × g for 20 minutes and the precipitates were resuspended for cytoplasmic Ca2+ determination.
Golgi vesicle preparation
Golgi vesicles were isolated according to Graham's protocol (Graham, 1997). First, rat brains were washed in pre-chilled dextran-homogenized medium and homogenized at approximately 3,500 × g for 1 minute. The homogenate was centrifuged at 4°C, 5,000 × g for 15 minutes. The upper supernatant was discarded and the yellow precipitate in the remaining upper layer was mildly retrieved by a glass stick and resuspended in 5-mL dextran-homogenized medium. The suspension was set to 10-mL 1.2 M sucrose gradient and was centrifuged at 4°C, 100,000 × g for 30 minutes. Golgi vesicles were present in the upper layer of the gradient suspension.
Fluorescent spectrophotometer detection of Ca2+
The specimen of nerve cells and Golgi vesicles were resuspended in Dulbecco's modified Eagle's medium with Fura-2/AM. Subsequently, the suspension was incubated at 37°C for 60 minutes and then centrifuged at 2,000 × g for 5 minutes. The supernatant was resuspended in 5-mL Hank's Balanced Salt Solution to detect the Ca2+ concentration.
The sample in Hank's Balanced Salt Solution was detected in the fluorescent spectrophotometer (HITACH F-7000), and the excited wavelength was 380 nm. The real-time fluorescence (F) was detected at 510 nm. The maximum fluorescence (Fmax) was determined after adding Triton X-100 and the minimum fluorescence (Fmin) was measured after appending 20 mM ethylenebis(oxyethylenenitrilo)tetraacetic acid, followed by the manipulation of Triton. In addition, before formal determination, fluorescent wave scanning was taken to verify whether the maximum excited wavelength could reach 380 nm.
Calcium concentration was calculated by the following formula: [Ca2+] = Kd × (F–Fmin)/(Fmax–F), and Kd in this formula is the constant of 224 nM.
Statistical analysis
The data were evaluated as the mean ± SEM, and analyzed by SPSS 13.0 software (SPSS, Chicago, IL, USA). Statistical comparisons of calcium concentration of cytoplasm/Golgi vesicles and fluorescent density of SPCA1 antibodies from different groups or subgroups were performed by one-way analysis of variance respectively. The least significance difference test was used for post hoc testing. A value of P < 0.05 was considered statistically significant.
Results
Ca2+ concentration changes in neuronal cytoplasm during global cerebral ischemia and reperfusion
Calcium detection in isolated nerve cells showed that cytoplasmic Ca2+ increased rapidly in ischemia and early reperfusion and then slightly but appreciably declined. After reperfusion of 3 hours, the Ca2+ labeled by Fura-2/AM was at its maximum value, 1,695.5 nM, which was eight times higher than the control (P = 0.001). Calcium overload was sustained and the concentration remained twice higher than the control (P = 0.003) until the time of reperfusion for 7 days. Although calcium overload remained to the time of reperfusion for 7 days, there was an obvious alleviation in late reperfusion (Figure 1A).
Figure 1.
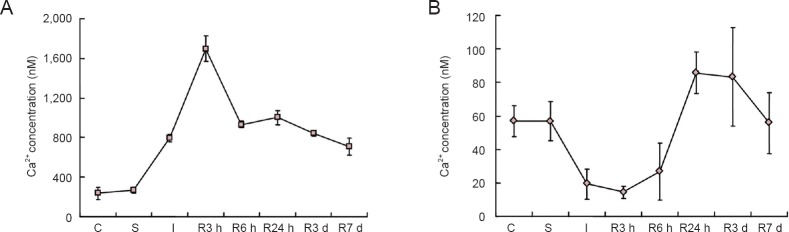
Changes in Ca2+ concentration in cytoplasm and separated Golgi vesicles in nerve cells during cerebral ischemia and reperfusion.
(A) Changes in cytoplasmic Ca2+, which increased rapidly during ischemia and early reperfusion and then slightly but appreciably declined. (B) Changes in Golgi Ca2+, which showed an opposite trend to cytoplasmic Ca2+. C: Control group; S: sham operation group; I: ischemia group; R: reperfusion group; h: hours; d: days. The data are expressed as the mean ± SEM (calculated from data of five pieces of brain section) and oneway analysis of variance followed by the least significance difference test was used.
Ca2+ concentration changes in neuronal Golgi vesicles during global cerebral ischemia and reperfusion
Calcium determination in isolated Golgi vesicles of nerve cells showed that Ca2+ decreased rapidly during ischemia and early reperfusion and then sharply increased. When reperfusion was prolonged, the Ca2+ labeled by Fura-2/AM achieved 14.35 μM, the minimum value, which is only one quarter of the control or sham operation (P = 0.005). In the late phase of reperfusion, from 6 to 24 hours, the calcium content of Golgi vesicles was elevated to a much higher level than control or sham operation group. Over reperfusion for 7 days, it recovered to the original state (Figures 1B and 2).
Figure 2.
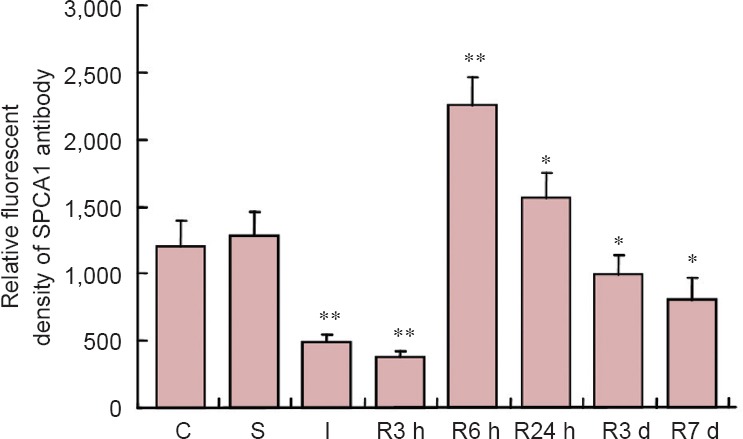
Data of fluorescent density of SPCA1 primary antibody during ischemia and reperfusion as detected by fluorescence immunohistochemistry.
*P < 0.05, **P < 0.01, vs. C and S. The data are expressed as the mean ± SEM (calculated from data of five pieces of brain section) and oneway analysis of variance and the least significance difference test were used. C: Control group; S: sham operation group; I: ischemia group; R: reperfusion group; SPCA1: secretory pathway Ca2+ ATPase 1; h: hours; d: days.
Fluorescence in situ detection of SPCA1
Direct fluorescence in situ detection of SPCA1 demonstrated that SPCA1 was widely expressed in cerebral tissues including cortex and hippocampus neurons (Figure 3) and showed an obvious change in response to ischemia and reperfusion (Figure 4A–H). When ischemia and early reperfusion occurred, the fluorescent density of SPCA1 primary antibodies was much weaker than the control, in particular, the density was at its lowest point during reperfusion (Figure 4C and D). After cerebral ischemia had ended, at reperfusion for approximately 6 hours, the fluorescent density increased to almost 200% of the control (P < 0.01; Figures 2 and 4E). Although the level of fluorescent density increased rapidly, it recovered to the normal level slowly (Figure 4F–H).
Figure 3.
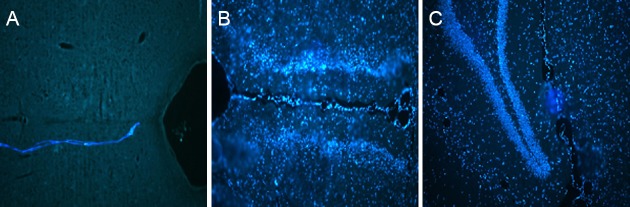
Broad expression of SPCA1 in the brain (immunofluorescence, fluorescence microscope).
(A) Image of specimen without SPCA1 antibody, used as a reference. No fluorescent signals are detected. (B, C) SPCA1 expression in the brain, particularly in the cortex and hippocampus neurons, at 6 hours after reperfusion. SPCA1-immunoreactive cells show blue fluorescence. SPCA1: Secretory pathway Ca2+ ATPase 1.
Figure 4.
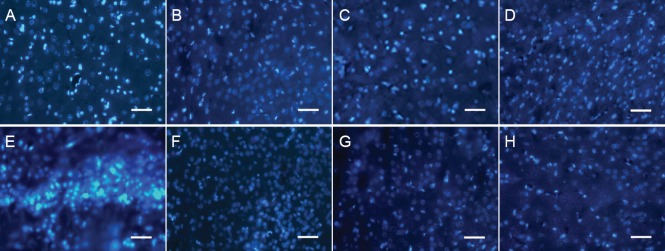
Fluorescent immunoreactivity of SPCA1 during ischemia and reperfusion (immunofluorescence, fluorescence microscope).
SPCA1 expression was analyzed by fluorescence in situ immunohistochemistry. Neuronal cells expressing SPCA1 are marked by blue fluorescence. (A, B) Images of control and sham operation groups; (C) ischemia group; (D–H) reperfusion group, R3 h, R6 h, R24 h, R3 d and R7 d. SPCA1-immunoreactive cells show blue fluorescence. Scale bars: 20 μm. R: Reperfusion; SPCA1: secretory pathway Ca2+ ATPase 1; h: hours; d: days.
Protein expression of SPCA1
Western blot assay for SPCA1 protein provided a similar but alternative process to immunohistological detection. Protein expression of SPCA1 manifested a low-high-low tendency during cerebral ischemia and reperfusion (Figure 5). The lowest state occurred in the phase of reperfusion and the highest state occurred in the adjacent 6 hours.
Figure 5.

Western blot assay of SPCA1 during ischemia and reperfusion.
In C and S, almost no difference was detected in SPCA1 expression. In I and R3 h, an obvious decline occurred. In R6 h, a dramatic increase appeared. After R24 h, the expression decreased very slowly. C: Control group; S: sham operation group; I: ischemia group; R: reperfusion group; SPCA1: secretory pathway Ca2+ ATPase 1; h: hours; d: days.
Discussion
Golgi regulation of neuronal cytoplasmic calcium during ischemia and reperfusion
Calcium overload and subsequent sustained calcium levels cause neuronal cell death and are essential events of cerebral IRI (Krino, 1982; Li et al., 2013). Generally, calcium overload is attributed to the release of inner and outer Ca2+ stores. On one hand, reactive oxygen species and reactive nitrogen species produced by oxidative stress should attack cell membranes and inner membranes of the endoplasmic reticulum and mitochondria, and cause the leakage of Ca2+ into the cytoplasm (Penna et al., 2009). On the other hand, opening of Ca2+ channels on the cell membrane and endoplasmic reticulum and mitochondria should also release Ca2+ into the cytoplasm (Murphy and Steenbergen, 2008; Senkal et al., 2011).
Traditionally, the inner Ca2+ store, which can regulate cytoplasmic calcium concentration, refers to endoplasmic reticulum and mitochondria, but the Golgi apparatus can condense a high level of calcium and act as a store in cytoplasm and its role in calcium regulation is not clear (Wuytack, 2003; Vandecaetsbeek, 2011). The Golgi apparatus contains IP3R, SERCA and the high-affinity pump SPCA (Pinton et al., 1998), and thus it is reasonable to assume it takes part in cytoplasmic regulation.
Our results show changes in calcium content in Golgi vesicles during ischemia and reperfusion and indicate the changes are opposite to the calcium concentration in cytoplasm. At point of ischemia and during early reperfusion, the dramatic decline of Golgi vesicles suggests that the Golgi apparatus participates in establishing calcium overload of cytoplasm through the pathway of the only calcium channel, IP3R. In late reperfusion, the Ca2+ concentration in Golgi vesicles recovers rapidly, which suggests that Golgi apparatus can “alleviate” the situation of calcium overload. This also provides evidence that the response of Golgi apparatus stress to cerebral ischemia and reperfusion can be realized in remodeling homeostasis of cytoplasmic Ca2+.
It is also a valuable question to study further: how does Ca2+-ATPase contribute to the “alleviating” role of the Golgi apparatus? The current four-vessel occlusion research model is too extreme to assess the “alleviating” role of Golgi apparatus accurately. Cells exposed to oxygen and glucose deprivation are probably a more appropriate choice.
Possible involvement of SPCA1 in Golgi apparatus stress to cerebral ischemia and reperfusion
Based on the discussion above, there is no doubt that Golgi apparatus stress in Ca2+ regulation is involved in cerebral ischemia and reperfusion, and the “alleviating” role of Golgi apparatus is dependent on Ca2+-ATPase. As a pump with high affinity to Ca2+ and located specifically on the Golgi apparatus membrane, SPCA probably participates in the Golgi apparatus stress to cerebral ischemia and reperfusion. Furthermore, the conclusion that SPCA1 gene is housekeeping (Antebi et al., 1992; Wuytack et al., 2003; Missiaen et al., 2007) is also proved by our immunochemical observation of broad distribution in neuronal tissues. Its characteristics of housekeeping allow them to realize the function of Ca2+ regulation in neuronal cells.
Pavlikova et al. (2009) proved that SPCA1 diminishes in ischemia and early reperfusion and increases in late reperfusion. Our immunochemistry and western blot assay of SPCA1 also demonstrated the following two points: (1) SPCA1 is sensitive to hypoxia and low sugar supply during cerebral ischemia and reperfusion. During ischemia and early reperfusion, the expression of SPCA1 is obviously limited by hypoxia and low glucose supply. On the contrary, 6 hours after reperfusion, which is much earlier than Golgi Ca2+ elevation, SPCA1 expression showed a recovery. (2) The expression of SPCA1 is consistent with the alteration of Ca2+ in Golgi vesicle and opposite to cytoplasmic calcium. For example, when cytoplasmic Ca2+ reached the peak, the expression level of SPCA1 arrived at its lowest point, which responded to Golgi apparatus calcium variation. Moreover, from reperfusion to 6 hours, when the activity of SPCA1 recovered rapidly, which caused the rapid rise of Ca2+ in Golgi vesicles, cytoplasmic Ca2+ declined quickly, though it maintained the situation of overload. According to this, it can be concluded that SPCA1 is tightly related to the cytoplasmic Ca2+ regulation in the Golgi apparatus stress to cerebral ischemia and reperfusion. In later reperfusion, the “alleviation” role of Golgi apparatus should certainly contribute to the activity of SPCA1.
Although the special role of SPCA1 on Golgi apparatus in cerebral ischemia and reperfusion has been well described here and in some other studies (Wuytack et al., 2002, 2003; Missiaen et al., 2007; Lehotský et al., 2009; Lissandron et al., 2010; Micaroni, 2012; Li et al., 2013), the function of SERCA, which is also located on the Golgi apparatus membrane and possesses higher calcium transport efficiency and lower calcium affinity, and its cooperation with SPCA1 has been not assessed. Studies of Golgi apparatus Ca2+ regulation showed that SERCA provides a significant Ca2+ uptake into Golgi apparatus cisternae (Wuytack et al., 2002; Dode et al., 2005), thus more attention should be paid to its potential impact, although it is mainly located on endoplasmic reticulum.
Golgi apparatus regulation of Ca2+ homeostasis and its response to cerebral IRI
Ca2+ is a highly versatile intracellular signal that can regulate many different cellular functions (Berridge et al., 2000). The Ca2+-signaling system operates in many different ways to regulate cellular function. For example, it triggers exocytosis within microseconds, controls muscle contraction within milliseconds, operates over minutes to hours to drive events such as gene transcription and cell proliferation. In addition, all these actions contribute to calcium regulators, including organelles, factors and effectors (Carafoli et al., 2001; Berridge et al., 2003; Pavlikova et al., 2009). In any case, a constant intracellular store is required for intracellular Ca2+ homeostasis variation during cerebral ischemia and reperfusion at least. Intracellular Ca2+ homeostasis variation caused by Golgi apparatus which acts as an intracellular Ca2+ store should be identified in this procedure. The Golgi apparatus is the intermediate organelle of the inner membrane system, which “links” to endoplasmic reticulum and mitochondria (Dolman and Tepikin, 2006; Zanni et al., 2009; Micaroni et al., 2010; Pinchai et al., 2010). Roles of Golgi apparatus in Ca2+ regulation are widely found in neural development and diseases. Sepúlveda et al. (2009) found that the establishment of neural polarity was dependent on the Golgi Ca2+ homeostasis. Vanoevelen et al. (2007) had reviewed the disease in relation to Golgi apparatus calcium regulation. Therefore, we consider the Golgi apparatus a central point in cytoplasmic calcium signaling networks (Li et al., 2013). All these Golgi apparatus-related Ca2+ signals can contribute to the Golgi-specific Ca2+ pump, SPCA.
In our early study and review, Hu (2007) and Jiang et al. (2011) found that morphological changes of Golgi apparatus and its response to oxidative stress existed during cerebral IRI and that they were more or less related to the Golgi apparatus calcium regulatory effect.
Regarding morphological alteration, experiments by Zeng (2007) showed that there was no evidence of Golgi apparatus degradation in gerbils suffering from transient ischemia attack. This means Golgi apparatus degradation needs much more time and should be the terminal event of neuronal cell death in neurodegenerative disease (Fan et al., 2008). That is to say, morphological modification of Golgi apparatus results from cell death, but is not induced by apoptosis. With regards to control of Ca2+ homeostasis, our results and the associated function of Golgi apparatus in Ca2+ regulation discussed in neurodegenerative disease indicates that calcium overload in mild cerebral ischemia and reperfusion can be rescued by Golgi apparatus and other Ca2+-regulated organelles to avoid neuronal cell death. Both the morphological alteration and Ca2+ homeostasis self-maintenance should be the exact explanation on the self-recovery of transient ischemia attack.
Oxidative stress is known to be the most direct response in the event of injury of cerebral ischemia and reperfusion. It not only induces and promotes calcium overload, but also can be enhanced by calcium overload. Jiang's review appropriately elucidated the role of SPCA on oxidative stress and argued that Golgi apparatus stress played an essential role, which is almost identical to endoplasmic reticulum stress (Jiang et al., 2011). Based on the function of SPCA in oxidative stress to cerebral ischemia and reperfusion, it can be concluded that Golgi apparatus, especially the mature cisternae, can alleviate the calcium overload through declining the oxidative stress. Obviously, on account of discussion in self-repairing during cerebral ischemia and reperfusion, neuronal protective drugs would be the best choice for clinical treatment for these diseases. It is interesting that SPCA played an important role in alleviation of cadmium toxicity in yeast for its function in decreasing the heavy metal ion concentration and declining the oxidative stress (Lauer Júnior et al., 2008).
Until now, there have been very few studies on the role of Golgi apparatus stress in neuronal ischemia injury, so the function of Golgi apparatus, especially the function of cytoplasmic calcium regulation in cerebral ischemia and reperfusion, is not so clear. In fact, the Ca2+ regulation function of Golgi apparatus is usually ignored by many researchers. The possible involvement of Golgi apparatus stress in the pathology and pathophysiology of common cardio-cerebrovascular disease should be investigated, as it may afford a new angle to the therapy of these diseases.
In summary, the expression of SPCA1 responds to cerebral IRI. Calcium overload is sustained in all processes of ischemia and reperfusion in cerebral neuronal cells. However, the situation of Ca2+ overload alleviated a little after reperfusion for 6 hours, and this is attributed to the regulatory role of Golgi apparatus. Moreover, based on the relationship between the expression of SPCA1 and calcium concentration in neuronal cytoplasm and Golgi apparatus, we can conclude that SPCA1, which is located on the Golgi apparatus membrane, may play an essential role in Golgi apparatus stress to cerebral IRI and alleviate calcium overload.
Acknowledgments
We thank Dr. Fei Gao from the School of Medicine of Jishou University in China for his help in establishing the animal model.
Footnotes
Funding: This study was supported by the National Natural Science Foundation of China, No. 81171239.
Conflicts of interest: None declared.
Copyedited by Paul P, Pack M, Wang J, Qiu Y, Li CH, Song LP, Zhao M
References
- Antebi A, Fink GR. The yeast Ca 2+ -ATPase homologue PMR1 is required for normal Golgi function and localizes in a novel Golgi-like distribution. Mol Biol Cell. 1992;3:633–654. doi: 10.1091/mbc.3.6.633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ausubel FM, Brent R, Kingston RE. Short Protocol of Molecular Biology. 5th Edition. Singapore: Johns-Wiley Press, Singapore; 2008. [Google Scholar]
- Behne MJ, Tu CL, Aronchik I, Epstein E, Bench G, Bikle DD, Pozzan T, Mauro TM. Human keratinocyte ATP2C1 localizes to the Golgi and controls Golgi Ca 2+ stores. J Invest Dermatol. 2003;121:688–694. doi: 10.1046/j.1523-1747.2003.12528.x. [DOI] [PubMed] [Google Scholar]
- Berridge MJ, Lipp P, Bootman MD. The versatility and universality of calcium signalling. Nature Rev Mol Cell Biol. 2000;1:11–21. doi: 10.1038/35036035. [DOI] [PubMed] [Google Scholar]
- Berridge MJ, Bootman MD, Roderick HL. Calcium signaling: dynamics, homeostasis and remodeling. Nat Rev. 2003;4:517–529. doi: 10.1038/nrm1155. [DOI] [PubMed] [Google Scholar]
- Brethes D, Dayanithi G, Letellier L, Nordmann JJ. Depolarization-induced Ca2 + increase in isolated neurosecretory nerve terminals measured with fura-2. Proc Nat Acad Sci. 1987;84:1439–1443. doi: 10.1073/pnas.84.5.1439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carafoli E, Santella L, Brance D, Brisi M. Generation control, and processing of cellular calcium signals. Crit Rev Biochem Mol Biol. 2001;36:107–260. doi: 10.1080/20014091074183. [DOI] [PubMed] [Google Scholar]
- Chandra SE, Kable P, Morrison GH, Webb WW. Calcium sequestration in the Golgi apparatus of cultured mammalian cells revealed by laser scanning confocal microscopy and ion microscopy. J Cell Sci. 1991;100:747–752. doi: 10.1242/jcs.100.4.747. [DOI] [PubMed] [Google Scholar]
- Demestre M, Howard RS, Orrell RW, Pullen AH. Serine proteases purified from sera of patients with amyotrophic lateral sclerosis (ALS) induce contrasting cytopathology in murine motoneurones to IgG. Neuropathol Appl Neurobiol. 2006;32:141–156. doi: 10.1111/j.1365-2990.2006.00712.x. [DOI] [PubMed] [Google Scholar]
- Dildy JE, Leslie SW. Ethanol inhibits NMDA-induced increases in free intracellular Ca 2+ in dissociated brain cells. Brain Res. 1989;499:383–387. doi: 10.1016/0006-8993(89)90789-0. [DOI] [PubMed] [Google Scholar]
- Dode L, Andersen JP, Raeymaekers L, Missiaen L, Vilsen B, Wuytack F. Functional comparison between secretory pathway Ca 2+/Mn 2+ -ATPase (SPCA) 1 and sarcoplasmic reticulum Ca 2+ -ATPase (SERCA) 1 isoforms by steady-state and transient kinetic analyses. J Biol Chem. 2005;280:39124–39134. doi: 10.1074/jbc.M506181200. [DOI] [PubMed] [Google Scholar]
- Dolman NJ, Tepikin AV. Calcium gradients and the Golgi. Cell Calcium. 2006;40:505–512. doi: 10.1016/j.ceca.2006.08.012. [DOI] [PubMed] [Google Scholar]
- Durr G, Strayle J, Plemper R, Elbs S, Klee SK, Catty P, Wolf DH, Rudolph HK. The medial-Golgi ion pump Pmr1 supplies the yeast secretory pathway with Ca 2+ and Mn 2+ required for glycosylation sorting and endoplasmic reticulum-associated protein degradation. Mol Biol Cell. 1998;9:1149–1162. doi: 10.1091/mbc.9.5.1149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan J, Hu ZP, Zeng LW, Lu W, Tang XQ, Zhang J, Li T. Golgi apparatus and neurodegenerative diseases. Int J Devl Neurosci. 2008;26:523–534. doi: 10.1016/j.ijdevneu.2008.05.006. [DOI] [PubMed] [Google Scholar]
- Graham JM. Subcellular Fractionation-A Practical Approach. Oxford: Oxford University Press, UK; 1997. Homogenization of cells and tissues. [Google Scholar]
- Hu Z, Bonifas JM, Beech J, Bench G, Shigihara T, Ogawa H, Ikeda S, Mauro T, Epstein EH., Jr Mutations in ATP2C1, encoding a calcium pump, cause Hailey-Hailey disease. Nat Genet. 2000;24:61–65. doi: 10.1038/71701. [DOI] [PubMed] [Google Scholar]
- Hu ZP, Zeng LW, Xie LS. Morphological alteration of Golgi apparatus and subcellular compartmentalization of TGF-β1 in Golgi apparatus in gerbils following transient forebrain ischemia. Neurochem Res. 2007;32:1927–1931. doi: 10.1007/s11064-007-9382-1. [DOI] [PubMed] [Google Scholar]
- Ferrer I, Planas AM. Signaling of cell death and cell survival following focal cerebral ischemia: life and death struggle in the penumbra. J Neuropatho Exp Neuro. 2003;62:329–339. doi: 10.1093/jnen/62.4.329. [DOI] [PubMed] [Google Scholar]
- Jiang Z, Hu ZP, Zeng LW, Lu W, Zhang HN, Li T, Xiao H. The role of the Golgi apparatus in oxidative stress: is this organelle less significant than mitochondria. Free Radic Biol Med. 2011;50:907–917. doi: 10.1016/j.freeradbiomed.2011.01.011. [DOI] [PubMed] [Google Scholar]
- Kienzle C, von Blume J. Secretory cargo sorting at the trans-Golgi network. Trends Cell Biol. 2014;24:584–93. doi: 10.1016/j.tcb.2014.04.007. [DOI] [PubMed] [Google Scholar]
- Krino T. Delayed neuronal death in the gerbil hippocampus following ischemia. Brain Res. 1982;239:57–69. doi: 10.1016/0006-8993(82)90833-2. [DOI] [PubMed] [Google Scholar]
- Lauer CM, Júnior, Bonatto D, Mielniczki-Pereira AA. The Pmr1 protein, the major yeast Ca 2+ -ATPase in the Golgi, regulates intracellular levels of the cadmiumion. FEMS Microbiol Lett. 2008;285:79–88. doi: 10.1111/j.1574-6968.2008.01214.x. [DOI] [PubMed] [Google Scholar]
- Lehotský J, Raèay P, Pavlíková M, Tatarková Z, Urban P, Chomová M, Kovalská M, Kaplán P. Cross-talk of intracellular calcium stores in the response to neuronal ischemia and ischemic tolerance. Gen Physiol Biophys. 2009;28:F104–F114. [PubMed] [Google Scholar]
- Li LH, Tian XR, Jiang Z, Zeng LW, He WF, Hu ZP. The Golgi apparatus: panel point of cytosolic Ca 2+ regulation. Neurosignal. 2013;21:272–284. doi: 10.1159/000350471. [DOI] [PubMed] [Google Scholar]
- Li T, Mo X, Jiang Z, He W, Lu W, Zhang H, Zhang J, Zeng L, Yang B, Xiao H, Hu Z. Study of αB-crystallin expression in gerbil BCAO model of transient global cerebral ischemia. Oxid Med Cell Longev. 2012;2012:945071. doi: 10.1155/2012/945071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin P, Li F, Zhang YW, Huang H, Tong G, Farquhar MG, Xu H. Calnuc binds to Alzheimer's beta-amyloid precursor protein and affects its biogenesis. J Neurochem. 2007;100:1505–1514. doi: 10.1111/j.1471-4159.2006.04336.x. [DOI] [PubMed] [Google Scholar]
- Lissandron V, Podini P, Pizzo P. Unique characteristics of Ca 2+ homeostasis of the trans-Golgi compartment. Proc Natl Acad Sci U S A. 2010;107:9198–9203. doi: 10.1073/pnas.1004702107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCue HV, Patel P, Herbert AP. Solution NMR structure of the Ca 2+ -bound N-terminal domain of CaBP7: a regulator of golgi trafficking. J Biol Chem. 2012;287:38231–38243. doi: 10.1074/jbc.M112.402289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Micaroni M, Mironov AA. Roles of Ca2+ and secretory pathway Ca-ATPase pump type 1 (SPCA1) in intra-Golgi transport. Commun Integr Biol. 2010;3:504–507. doi: 10.4161/cib.3.6.13211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Micaroni M, Perinetti G, Di Giandomenico D, Bianchi K, Spaar A, Mironov AA. Synchronous intra- Golgi transport induces the release of Ca 2+ from the Golgi apparatus. Exp Cell Res. 2010;316:2071–2086. doi: 10.1016/j.yexcr.2010.04.024. [DOI] [PubMed] [Google Scholar]
- Micaroni M. Calcium around the Golgi apparatus: implication for intracellular membrane trafficking. Adv Exp Med Biol. 2012;740:439–460. doi: 10.1007/978-94-007-2888-2_18. [DOI] [PubMed] [Google Scholar]
- Missiaen L, Dode L, Vanoevelen J, Raeymaekers L, Wuytack F. Calcium in the Golgi apparatus. Cell Calcium. 2007;41:405–416. doi: 10.1016/j.ceca.2006.11.001. [DOI] [PubMed] [Google Scholar]
- Murphy E, Steenbergen C. Mechanism underlying Acute protection from cardio ischemia and reperfusion injury. Physiol Rev. 2008;88:581–609. doi: 10.1152/physrev.00024.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pavlikova M, Tatarkova Z, Sivoňova M, Kaplan P, Križanova O, Lehotsky J. Alterations induced by ischemic preconditioning on secretory pathways Ca 2+ -ATPase (SPCA) gene expression and oxidative damage after global cerebral ischemia/reperfusion in rats. Cell Mol Neurobiol. 2009;29:909–916. doi: 10.1007/s10571-009-9374-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penna C, Mancardi D, Rastaldo R, Pagliaro P. Cardioprotection: a radical view free radicals in pre and post-conditioning. Biochim Biophysic Acta. 2009;1787:781–793. doi: 10.1016/j.bbabio.2009.02.008. [DOI] [PubMed] [Google Scholar]
- Pezzati R, Bossi M, Podini P, Meldolesi J, Grohovaz F. High-resolution calcium mapping of the ER-Golgi-exocytic membrane system. Electron energy loss imaging analysis of quick frozen-freeze dried PC12 cells. Mol Biol Cell. 1997;8:1501–1512. doi: 10.1091/mbc.8.8.1501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinchai N, Juvvadi PR, Fortwendel JR. The Aspergillus fumigatus P- type Golgi apparatus Ca 2+/Mn 2+ ATPase PmrA is involved in cation homeostasis and cell wall integrity but is not essential for pathogenesis. Eukaryot Cell. 2010;9:472–476. doi: 10.1128/EC.00378-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinton P, Pozzan T, Rizzuto R. The Golgi apparatus is an inositol 1, 4, 5-trisphosphate- sensitive Ca 2+ store with functional properties distinct from those of the endoplasmic reticulum. EMBO J. 1998;17:5298–5308. doi: 10.1093/emboj/17.18.5298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pressler RT, Rozman PA, Strowbridge BW. Voltage-dependent intrinsic bursting in olfactory bulb Golgi cells. Learn Mem. 2013;20:459–466. doi: 10.1101/lm.031856.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pulsinelli WA, Buchan AM. The four-vessle occlusion rat model: method for complete occlusion of vertebral circulation. Stroke. 1988;19:913–914. doi: 10.1161/01.str.19.7.913. [DOI] [PubMed] [Google Scholar]
- Senkal CE, Ponnusamy S, Manevich Y. Alteration of ceramide synthase 6/C16-ceramide induces activating transcription factor 6-mediated endoplasmic reticulum (ER) stress and apoptosis via perturbation of cellular Ca 2+ and ER/Golgimembrane network. J Biol Chem. 2011;286:42446–42458. doi: 10.1074/jbc.M111.287383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sepúlveda MR, Vanoevelen J, Raeymaekers L. Silencing the SPCA1 (secretory pathway Ca 2+ -ATPase isoform 1) impairs Ca 2+ homeostasis in the Golgi and disturbs neural polarity. J Neurosci. 2009;29:12174–12182. doi: 10.1523/JNEUROSCI.2014-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shull GE, Miller ML, Prasad V. Secretory pathway stress responses as possible mechanisms of disease involving Golgi Ca 2+ pump dysfunction. Biofactors. 2011;37:150–158. doi: 10.1002/biof.141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudbrak R, Brown J, Dobson-Stone C, Carter S, Ramser J, White J, Healy E, Dissanayake M, Larregue M, Perrussel M, Lehrach H, Munro CS, Strachan T, Burge S, Hovnanian A, Monaco AP. Hailey-Hailey disease is caused by mutations in ATP2C1 encoding a novel Ca2+ pump. Hum Mol Genet. 2000;9:1131–1140. doi: 10.1093/hmg/9.7.1131. [DOI] [PubMed] [Google Scholar]
- Uematsu D, Greenberg DJH, Reivich M, Karp A. In vivo measurement of cytosolic free calcium during cerebral ischemia and reperfusion. Ann Neurol. 1988;24:420–428. doi: 10.1002/ana.410240311. [DOI] [PubMed] [Google Scholar]
- Van Baelen K, Dode L, Vanoevelen J, Callewaert G, De Smedt H, Missiaen L, Parys JB, Raeymaekers L, Wuytack F. The Ca 2+/Mn 2+ pumps in the Golgi apparatus. Biochim Biophys Acta. 2004;1742:103–112. doi: 10.1016/j.bbamcr.2004.08.018. [DOI] [PubMed] [Google Scholar]
- Vandecaetsbeek I, Vangheluwe P, Raeymaekers L, Wuytack F, Vanoevelen J. The Ca 2+ pumps of the endoplasmic reticulum and Golgi apparatus. Cold Spring Harb Perspect Biol. 2011;3 doi: 10.1101/cshperspect.a004184. pii:a004184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanoevelen J, Dode L, Raeymaekers L. Diseases involving the Golgi calcium pump. Subcell Biochem. 2007;45:385–404. doi: 10.1007/978-1-4020-6191-2_14. [DOI] [PubMed] [Google Scholar]
- Wong AK, Capitanio P, Lissandron V. Heterogeneity of Ca2+ handling among and within Golgi compartments. J Mol Cell Biol. 2013;5:266–276. doi: 10.1093/jmcb/mjt024. [DOI] [PubMed] [Google Scholar]
- Wuytack F, Raeymaekers L, Missiaen L. Molecular physiology of the SERCA and SPCA pumps. Cell Calcium. 2002;32:279–305. doi: 10.1016/s0143416002001847. [DOI] [PubMed] [Google Scholar]
- Wuytack F, Raeymaekers L, Missiaen L. PMR1/SPCA Ca 2+ pumps and the role of the Golgi apparatus as a Ca 2+ store. Pflugers Arch Euro J Physiol. 2003;446:148–153. doi: 10.1007/s00424-003-1011-5. [DOI] [PubMed] [Google Scholar]
- Xue SB, Nicoud MR, Cui J, Jovin DJA. High concentration of calcium ions in Golgi apparatus. Cell Res. 1994;4:97–108. [Google Scholar]
- Zak M, Pfister M, Blin N. The otoferlin interactome in neurosensory hair cells: significance for synaptic vesicle release and trans-Golgi network. Int J Mol Med. 2011;28:311–314. doi: 10.3892/ijmm.2011.716. [DOI] [PubMed] [Google Scholar]
- Zanni E, Farina F, Ricci A, Mancini P, Frank C, Palleschi C, Uccelletti D. The Golgi alpha-1, 6 mannosyltransferase KlOch1p of Kluyveromyces lactis is required for Ca 2+/calmodulin-based signaling and for proper mitochondrial functionality. BMC Cell Biol. 2009;10:86. doi: 10.1186/1471-2121-10-86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng LW. Changsha: Central South University, China; 2007. TGF-β1 and Golgi apparatus alteration in ischemia and minocycline treated gerbils and subcellular compartmentalization of TGF-β1 in Golgi apparatus. [Google Scholar]