

Abstract
AIM: To prove that neurons in the different structures of the brainstem that express tyrosine hydroxylase (TH) are involved in the transmission and modulation of visceral or somatic nociceptive information in rat.
METHODS: Immunohistochemical double-staining method was used to co-localize TH and Fos expression in neurons of the rat brainstem in visceral or subcutaneous noxious stimulation models.
RESULTS: Neurons co-expressing TH/Fos were observed in lateral reticular nucleus (LRT), rostroventrolateral reticular nucleus (RVL), solitary tract nucleus (SOL), locus coeruleus (LC), A5, A7 neuronal groups and ventrolateral subdivision of the periaqueductal gray (vlPAG) in both models. But the proportion and number of the double-labeled neurons responding to the two noxious stimuli were significantly different in the LRT, RVL and LC nuclei. The proportion and number of the TH/Fos double-labeled neurons in the visceral pain model were smaller than that in the subcutaneous pain model. However, in the case of SOL, they were similar in the two models.
CONCLUSION: Differences of Fos expression in TH immunoreactive neurons in animals after visceral and somatic noxious stimulation indicate that the mechanisms of the transmission and modulation of visceral nociceptive information in the brainstem may be different from that of somatic nociceptive information.
INTRODUCTION
Tyrosine hydroxylase (TH) is a catecholamine (including noradrenaline, adrenaline and dopamine) synthesis enzyme. In the brainstem, except that substantia nigra (SN) TH is mainly involved in the locomotive modulation, TH immunoreactive (TH-ir) neurons are mainly located in the lateral reticular nucleus (LRT), rostroventrolateral reticular nucleus (RVL), solitary tract nucleus (SOL), locus coeruleus (LC), A5, A7 neuronal groups and ventrolateral subdivision of the midbrain periaqueductal gray (vlPAG). All these regions, except for RVL and SOL, are regarded as the origins of the descending inhibitory system, which uses predominantly noradrenaline (NA) and 5-hydroxytryptamine (5-HT) as neurotransmitters[1,2]. Morphological evidence has shown that these nuclei directly or indirectly project to the superficial laminea of the spinal and medullary dorsal horns and SOL, forming intrinsic circuits that modulate the visceral and peripheral nociceptive message transmission[1]. Electrophysiolgical results proved that NTS, VLM, PBL, and vlPAG were involved in visceral message transmission and modulation and electrical stimulation of vlPAG depressed visceral message transmission[3-6]. RVL is mainly associated with cardiovascular regulation[7] and projects to spinal autonomic nuclei[8], while SOL is mainly involved in the visceral nociceptive message transmission, convergence and cardiovascular reflection[9,10].
c-fos proto-oncogene is expressed within the neurons following voltage-gated calcium entry into the cell[11,12]. Fos protein, which is encoded by c-fos, has been regarded as a third messenger molecule which coupled the short term extracellular signals with the long term alteration in cell function after neurons were excited[13,14]. Mapping studies have shown that noxious stimulation could induce Fos expression in the central neuronal system and visceral organs[15-21]. Thus, Fos is usually used as a marker to indicate the activation of neurons[22].
It is well known that the brainstem noradrenergic (NAergic) system plays an important role in the process of antinociception and that the mechanism of visceral noxious message modulation is different from those of somatic noxious message modulation[23]. It is reasonable for us to propose that the location of Fos expression, under the effects of different noxious stimuli in the structures of brainstem, which express TH, could be diverse. In order to prove this hypothesis, we studied the expression pattern of Fos induced by visceral or subcutaneous noxious stimuli as well as the co-localization of Fos and TH protein expression using an immunohistochemical double-staining method.
MATERIALS AND METHODS
Materials
Experiments were carried out using 40 Sprague-Dawley male rats (weighing 200-250 g). The rats were kept in a dark facility for 24 hours before the experiment, and then randomly divided into two experimental groups and two contro1 groups (for each groups, n = 10). All procedures described below were approved by the Committee for Animal Care and Usage for Research and Education from the Fourth Military Medical University (Xi’an, P. R. China).
Methods
All rats were anesthetized by being placed into a transparent glass container with methoxyflurane gas. After anesthesia, 2 mL of 100 mL/L formalin or 2 mL distilled water was perfused into the oesophagus and stomach through a fine plastic tube, or 0.1 mL of 100 mL/L formalin or 0.1 mL distilled water was injected subcutaneously into the left forepaw in the rats of experimental or control groups respectively. Then the animals were put back into the cages and 2 hours later, all rats were anesthetized with sodium pentobarbital intraperitoneally (60 mg/kg) were intracardially perfused with l00 mL of saline followed by 500 mL of 4% paraformaldehyde in 0.1 mol/L phosphate buffer (PB, pH7.4). The brainstems were immediately removed, placed into the same fresh fixative for 4 hours (4 °C), and saturated with a solution of 300 g/L sucrose in 0.1 mol/L PB (pH7.4) overnight at 4 °C. Thirty-μm thick serial frontal sections were cut on a cryostat and serially collected into 5 dishes separately. Then all sections were washed with 0.01 mol/L phosphate-buffered saline (PBS, pH7.4).
The first section was initially incubated overnight at room temperature with rabbit antisera against Fos (1:1000 dilution; Santa Cruz, USA) in 0.0l mol/L PBS containing 50 mL/L normal horse serum, 3 g/L Triton X-100, 0.05 g/L NaN3 and 5 g/L carrageenan (PBS-NHS). Secondly, the sections were incubated with goat anti-rabbit IgG (1:100 dilution; Capple, USA) in PBS-NHS for 3 hours, and followed by rabbit peroxidase anti-peroxidase (PAP) complex (1:200 dilution; Polysciences, USA) in PBS for l hour. Finally, Fos-immunoreactive (Fos-ir) neurons were visualized by incubated the sections with 0.05 mol/L Tris-HCl buffer (pH7.6) containing 0.4 g/L diaminobenzidine tetrahydrochloride (DAB) (Dojin, Japan), 0.03 mL/L H2O2 and 0.4 g/L Ni(NH4)2SO4 for 30-40 minutes.
The second section was incubated at room temperature with mouse antisera against TH (1:3000 dilution; Chemicon, USA) overnight, followed by biotinylated horse anti-mouse IgG (1:100 dilution; Vector, USA) for 3 hours, and then with ABC (avidin-biotin peroxidase comp1ex) Elite kit (1:50 dilution; Vector) for 1 hour. Finally, TH-immunoreactive (TH-ir) neurons were visualized by incubated the sections with 0.05 mol/L Tris-HCl buffer (pH7.6) containing 0.4 g/L DAB, 0.03 mL/L H2O2 and 0.4 g/L Ni(NH4)2SO4 for 30-40 minutes.
The third section was incubated overnight at room temperature with a mixture of rabbit anti-Fos and mouse anti-TH, followed by goat anti-rabbit IgG or biotinylated horse anti-mouse IgG for 3 hours, and then with rabbit PAP for 1 hour. The sections were firstly reacted with 0.05 mol/L Tris-HCl buffer (pH7.6) containing 0.4 g/L DAB, 0.03 mL/L H2O2 and 0.4 g/L Ni(NH4)2SO4 for 30-40 minutes to intensify DAB-based reaction to Fos-ir neurons. Then the reacted sections were washed three times in PBS for 30 minutes and incubated with ABC Elite kit for l hour, and finally reacted in the same reaction solution without Ni(NH4)2SO4 for 30-40 minutes to detect TH-ir neurons and TH/Fos double-labeled neurons. The number of TH-ir neurons, Fos-ir neurons, and TH/Fos double-1abeled neurons was counted from the sections from the third section of each rat. The distribution of the immunohistochemically stained neurons in two animals in each model was charted with the aid of a camera and one rat was used to represent each experimental group.
A mixture of normal rabbit serum and normal mouse serum was used to replace rabbit anti-Fos and mouse anti-TH antisera to incubate the fourth section. The subsequent staining procedures were the same as those used for the third section.
All the sections were washed and mounted onto gelatin-coated glass slides and examined with light microscope (BH-2; Olympus, Japan).
The fifth section was mounted onto gelatin-coated slides and processed for Nissl staining.
Statistical analysis
Values were expressed as -x ± s. Statistical differences between means of TH/Fos double-labeled neurons were determined by Student’s t test. The difference in proportion of TH/Fos double-labeled neurons to Fos-ir neurons was analyzed by Crosstab’s test. A P < 0.05 is considered to be significant.
RESULTS
On the first, second, and third sections from the experimental groups, three kinds of immunocytochemically positive neurons were identified in the nuclei of the brainstem: Fos-ir neurons, TH-ir neurons and TH/Fos double-labeled neurons. The cellular nuclei of the Fos-ir neurons were dark with oval or round shape, leaving their cytoplasm unstained. In contrast to Fos-ir neurons, TH-ir neurons showed brownish cytoplasm and dendrites with unstained nuclei. Scattered among those Fos- and TH-ir neurons, some neurons with dark nuclei and brownish cytoplasm exhibiting the co-localization of Fos and TH in the neurons were also observed.
On the first sections of experimental groups, Fos-ir neurons were mainly located in the caudal subnucleus of the spinal trigeminal nucleus (Sp5C), lateral reticular nucleus (LRT), rostroventrolateral reticular nucleus (RVL), nucleus solitary tract (SOL), nucleus raphe obscurus (ROb), nucleus raphe pallidus (RPa), paramedian raphe nucleus (PMN), lateral paragigantocellular nucleus (LPGi), alpha part of gigantocellular reticular nucleus (Gia), nucleus raphe magnus (NRM), locus coeruleus (LC), parabrachial nuclear complex (PBNC), nucleus raphe pontis (RPn), dorsal raphe nucleus (DR), ventrolateral subregion of the midbrain periaqueductal gray (vlPAG), and median raphe nucleus (MnR) (Figure 1, Figure 2). Fos-ir neurons in these nuclei of the subcutaneous noxious stimulation group was somewhat more than that in the visceral noxious stimulation group, but in the SOL, the number of the Fos-ir neurons in both visceral and subcutaneous noxious stimulations was similar between the experiment groups (Table 1). On the sections of the control groups, only a few Fos-ir neuronal cell bodies were observed in the superficial part of the Sp5C or SOL.
Figure 1.
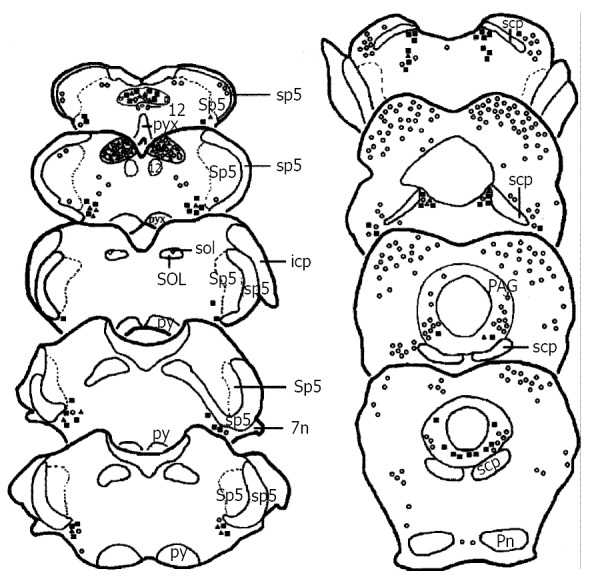
Drawing of the brainstem areas showing the distribution of TH, Fos and TH/Fos immunoreactive neurons after visceral noxious stimulation. Each■represents three TH-ir neurons. Each○represents five Fos-ir neurons. Each▲represents one TH/Fos double-labeled neuron. Abbreviations: 12: hypoglossal nuclei; 7n: facial nerve or its root; icp: inferior cerebellar peduncle; PAG: the midbrain periaqueductal gray; Pn: pontine nuclei; py: pyramidal tract; pyx: pyramidal decussation; scp: superior cerebellar peduncle; sol: solitary trace; SOL: solitary trace nuclei; sp5: spinal trigeminal tract; Sp5: spinal trigeminal nuclei.
Figure 2.
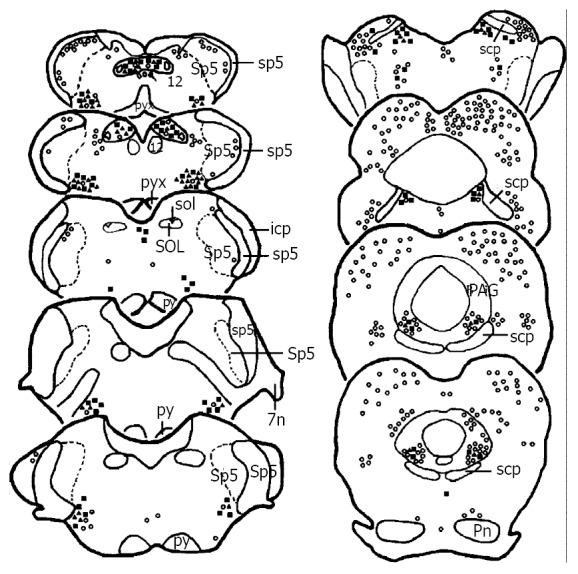
Drawing of the brainstem areas showing the distribution of TH (■), Fos (○) and TH/Fos (▲) immunoreactive neurons after subcutaneous noxious stimulation on the left forepaw. The ratios for each marker to present the number of TH, Fos and TH/Fos immunoreactive neurons and the abbreviations are the same as Figure 1.
Table 1.
The number and distribution of the immunopositive neurons in the brainstem
| Nuclei |
Nuclei |
Fos |
TH/Fos |
|||
| Visc | Sub | Visc | Sub | Visc | Sub | |
| SOL | 203 ± 13 | 214 ± 17 | 254 ± 31 | 99 ± 7 | 60 ± 5 | 54 ± 5 |
| LRT and RVL | 42 ± 15 | 252 ± 24 | 57 ± 7 | 151 ± 13a | 31 ± 4 | 90 ± 10a |
| LC | 328 ± 26 | 330 ± 25 | 42 ± 6 | 103 ± 12a | 26 ± 3 | 72 ± 9a |
Data are shown as (-x ± s)
P < 0.05 Sub vs Visc. Visc: Visceral noxious stimulation; Sub: Subcutaneous noxious stimulation; TH: TH-ir neurons; Fos: Fos-ir neurons; TH/Fos: TH/Fos double-labeled neurons.
On the second sections of both experimental and control groups, the distribution of TH-ir neuronal cell bodies were mainly in LRT, RVL, SOL, A4, A5, LC, A7, vlPAG and substantia nigra (SN) (Figure 1 and Figure 2). There was no remarkable difference in the number and distribution of TH-ir neurons between the experimental and control groups.
Immunohistochemical staining of the third sections from experimental groups showed, except that TH-ir and Fos-ir neurons were the same as that seen on the first and second sections of the experimental groups, TH/Fos double-labeled neurons were mainly distributed in the LRT, RVL, SOL, A5, LC, A7 and vlPAG in both visceral (Figure 1, Figure 3a, Figure 3a’, Figure 3c, Figure 3c’, Figure 3e and Figure 3e’) and subcutaneous (Figure 2, Figure 3b, Figure 3b’, Figure 3d, Figure 3d’, Figure 3f and Figure 3f’) pain models. In LRT, RVL, A5, A7, LC, and vlPAG, the proportion and number of TH/Fos double-labeled neurons in visceral pain model were less than those in subcutaneous pain model (Table 1). It is interesting to note that in the SOL, TH/Fos double-labeled neurons in both visceral and subcutaneous pain models were similar (Table 1). TH/Fos double-labeled neurons, with medium to large (20-35 μm) fusiform, oval, multipolar shaped neuronal cellular bodies, were observed.
Figure 3.
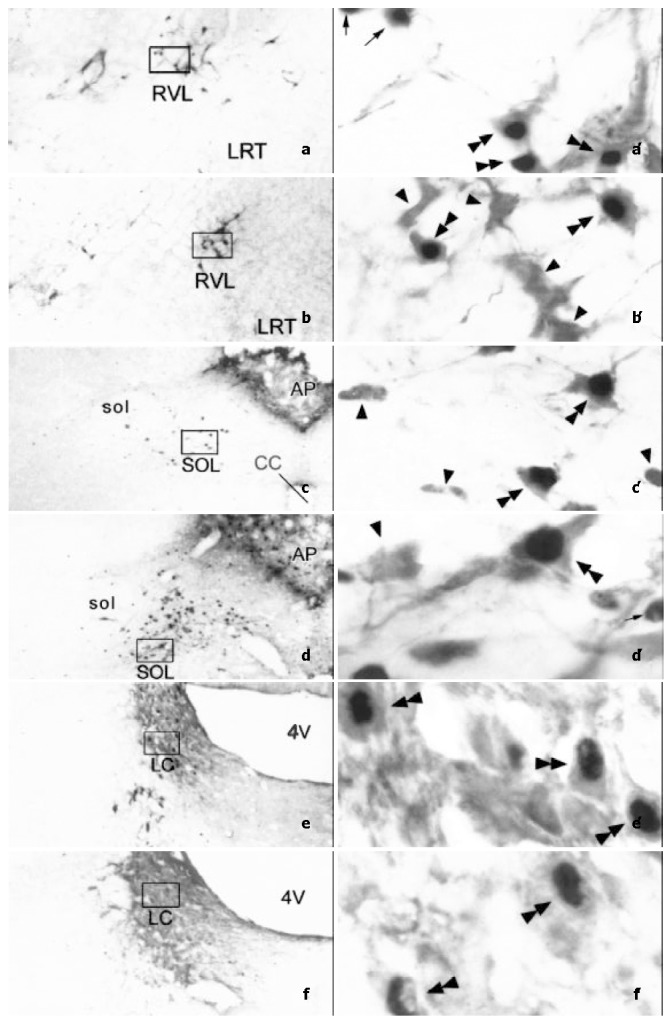
Photomicrographs showing the distribution and morphological characteristics of TH-ir neurons (arrowheads), Fos-ir neurons (arrows) and TH/Fos double-labeled neurons (double arrowheads) in SOL (a, a’ and b, b’), RVL (c, c’ and d, d’) and LC (e, e’ and f, f’). a, a’, c, c’ and e, e’ were taken from visceral noxious stimulating rats, while b, b’, d, d’ and f, f’ were taken from subcutaneous noxious stimulating rats, respectively. a, b, c, d, e and f are lower magnification photomicrograph (original magnification × 100). a’, b’, c’, d’, e’ and f’ are higher magnification photomicrograph (original magnification × 1000) of the rectangle in a, b, c, d, e and f, respectively. Abbreviations: 4V: The fourth ventricle. cc: Central canal. LC: Locus coeruleus. LRT: Lateral reticular nucleus. RVL: Rostroventrolateral reticular nucleus. sol: Solitary tract. SOL: Solitary tract nucleus.
In the brainstem, there was no expression of Fos and/or TH in the neurons from the fourth dish of both the groups.
DISCUSSION
TH is a critical enzyme for the synthesis of catecholamine. NA is thought to be one of the two neurotransmitters in descending inhibitory system, which can exert potent superspinal control of nociceptive information via various pathways[24,25]. In the brainstem, the descending inhibitory system is mainly composed of LRT, LC, vlPAG, rostral ventral ventromedial medulla and its surrounding reticular nuclei and other brainstem raphe nuclei[1]. NA can inhibit either excitatory transmitters (glutamate, substance P, etc.) released from primary afferent terminals or the EPSCs produced by primary afferent fibers via postsynaptic G-protein[23,26]. Intrathecal injection of naloxone could reverse the NA-produced antinociception at the spinal level[27], indicating the involvement of endogenous opiate peptides in the antinociceptive effects of the NA. LRT and LC including NAergic neurons project to SOL and are thought to be involved in visceral nociceptive message modulation and antinociception, which is proved by the electrophysiolgical experiments[28,29]. These results suggest that NA is an important neurotransmitter for the control of the nociceptive transmission in the central nervous system. In RVL and SOL, the neurons containing TH receive nociceptive message and produce somatic-visceral or viscera-somatic reflection[7,10], which do not belong to the descending inhibitory system. There exist many diseases with pain in visceral organs, and therapeutic methods for visceral pain are similar to those for somatic pain[30-34]. But it is not clear whether the neurons containing NA play different roles in different pain models. In order to clarify the functional roles of the NAergic neurons in the visceral and somatic pain models, we observed the expression pattern of both Fos and TH in which Fos protein was used as a marker to indicate TH-ir neurons activation using a immunohistochemical double-staining method.
In the present study, we observed TH/Fos double-labeled neurons in the LRT, RVL, SOL, A5, LC, A7 and vlPAG and then compared the distribution, number and proportion of TH-ir neurons related to visceral and somatic noxious stimuli. Our results suggested that visceral and subcutaneous nociceptive inputs triggered similar TH immunoreactive in neurons in the brainstem, and indicated that these nuclei in the brainstem were involved in both visceral and subcutaneous nociceptive message convergence and modulation. Our results also indicated that the mechanisms involved could be different. As we know neither the factors resulting in c-fos expression nor the consequences of Fos protein expression, caution must be taken in interpreting these data. It might not be the case that all labeled neurons are nociceptive; on the other hand, the absence of Fos in these neurons does not exclude that these neurons are nociceptive since some nociceptive neurons might not be activated by the experimental conditions used. Nevertheless, this method allows us to monitor the response of a large number of neurons to noxious stimulation in awaken animals, which complements to the electrophysiolgical tests on anaesthetized animals.
It is accepted that anesthesia is effective in triggering Fos expression in many brain regions, such as descending inhibitory system, RVL and SOL. More Fos-ir neurons were found in the experiment groups than those in the control groups. Based on these phenomena, we suggest that in addition to anesthetic effect, the noxious stimuli might be one of the inducing factors of Fos expression.
In LRT and RVL, there were fewer TH/Fos double-labeled neurons in visceral pain model than those in subcutaneous pain model. Both visceral and subcutaneous noxious stimuli can induce Fos expression in TH-ir neurons of LRT and RVL, indicating the occurrence of nociceptive neurons in this area. As this area is known to be crucial for vasomotor functions[28,35], and connected with SOL[36], NAergic neurons in this region are probably involved in eliciting the neurovegetative reaction induced by pain and producing somatic-visceral reflection.
Several evidences have suggested that the LC played a significant role in the descending inhibitory system[1,4]. TH/Fos double-labeled neurons were observed in the LC in both visceral and subcutaneous noxious stimuli models. This indicates the induction of NAergic neuronal activation in the LC in both models. The localization of these neurons is correspondent with the functional data implicating a role of LC in antinociception[29]. Therefore, we suggest that NAergic neurons in the LC probably receive the visceral nociceptive convergence and probably mediate the behavioral and physiological characteristics of visceral pain as well. The difference in the proportion and number of TH/Fos double-labeled neurons in visceral and subcutaneous pain models shows that NAergic neurons in LRT, RVL and LC may be mainly involved in somatic nociceptive message transmission and modulation.
SOL, which receives most of the visceral primary terminations from the majority of organs in the neck, chest and abdomen, is closely related to visceral nociceptive message transmission and convergence and pain-induced sensory-cardiovascular reflection[2,4,37]. Neurons in the SOL also projected to lateral parabrachial nucleus (LPB)[36]. These results provide anatomical evidence for physiological data, confirms the involvement of this nucleus in nociceptive transmission. In addition, our data proved that TH/Fos double-labeled neurons were mainly located in the middle portion of SOL in both models, but TH/Fos double-labeled neurons in the subcutaneous nociceptive model were more rostral than those in the visceral nociceptive model. These results indicate that TH/Fos double-labeled neurons are distributed topographically in two pain models and the mechanism that the NAergic neurons in SOL are involved in transmission and modulation of nociceptive information in visceral pain models is different from that in subcutaneous pain models.
ACKNOWLEDGMENTS
The authors are grateful to Ms. Yue-Ping Yuan and Ms. Miao-Li Zhang for their photographic assistance.
Footnotes
Supported by Grants-in-Aid from the National Natural Science Foundation of China (Nos. 39970239, 30070389, 3000052), the Foundation for University Key Teacher by the Ministry of Education of China and the National Program of Basic Research of China (G1999054004)
Edited by Yuan HT and Zhang JZ
References
- 1.Jones SL. Descending noradrenergic influences on pain. Prog Brain Res. 1991;88:381–394. doi: 10.1016/s0079-6123(08)63824-8. [DOI] [PubMed] [Google Scholar]
- 2.Mason P. Central mechanisms of pain modulation. Curr Opin Neurobiol. 1999;9:436–441. doi: 10.1016/S0959-4388(99)80065-8. [DOI] [PubMed] [Google Scholar]
- 3.Hudson PM, Semenenko FM, Lumb BM. Inhibitory effects evoked from the rostral ventrolateral medulla are selective for the nociceptive responses of spinal dorsal horn neurons. Neuroscience. 2000;99:541–547. doi: 10.1016/s0306-4522(00)00210-4. [DOI] [PubMed] [Google Scholar]
- 4.Benarroch EE. Pain-autonomic interactions: a selective review. Clin Auton Res. 2001;11:343–349. doi: 10.1007/BF02292765. [DOI] [PubMed] [Google Scholar]
- 5.Krout KE, Jansen AS, Loewy AD. Periaqueductal gray matter projection to the parabrachial nucleus in rat. J Comp Neurol. 1998;401:437–454. [PubMed] [Google Scholar]
- 6.Okada K, Murase K, Kawakita K. Effects of electrical stimulation of thalamic nucleus submedius and periaqueductal gray on the visceral nociceptive responses of spinal dorsal horn neurons in the rat. Brain Res. 1999;834:112–121. doi: 10.1016/s0006-8993(99)01593-0. [DOI] [PubMed] [Google Scholar]
- 7.Dampney RA. Functional organization of central pathways regulating the cardiovascular system. Physiol Rev. 1994;74:323–364. doi: 10.1152/physrev.1994.74.2.323. [DOI] [PubMed] [Google Scholar]
- 8.Romagnano MA, Harshbarger RJ, Hamill RW. Brainstem enkephalinergic projections to spinal autonomic nuclei. J Neurosci. 1991;11:3539–3555. doi: 10.1523/JNEUROSCI.11-11-03539.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Boscan P, Paton JF. Nociceptive afferents selectively modulate the cardiac component of the peripheral chemoreceptor reflex via actions within the solitary tract nucleus. Neuroscience. 2002;110:319–328. doi: 10.1016/s0306-4522(01)00585-1. [DOI] [PubMed] [Google Scholar]
- 10.Boscan P, Paton JF. Role of the solitary tract nucleus in mediating nociceptive evoked cardiorespiratory responses. Auton Neurosci. 2001;86:170–182. doi: 10.1016/S1566-0702(00)00255-1. [DOI] [PubMed] [Google Scholar]
- 11.Morgan JI, Curran T. Role of ion flux in the control of c-fos expression. Nature. 1986;322:552–555. doi: 10.1038/322552a0. [DOI] [PubMed] [Google Scholar]
- 12.Premkumar DR, Adhikary G, Overholt JL, Simonson MS, Cherniack NS, Prabhakar NR. Intracellular pathways linking hypoxia to activation of c-fos and AP-1. Adv Exp Med Biol. 2000;475:101–109. doi: 10.1007/0-306-46825-5_10. [DOI] [PubMed] [Google Scholar]
- 13.Curran T, Morgan JI. Superinduction of c-fos by nerve growth factor in the presence of peripherally active benzodiazepines. Science. 1985;229:1265–1268. doi: 10.1126/science.4035354. [DOI] [PubMed] [Google Scholar]
- 14.Saria A, Fischer HS, Humpel C, Pfattner A, Schatz DS, Schuligoi R. Margatoxin and iberiotoxin, two selective potassium channel inhibitors, induce c-fos like protein and mRNA in rat organotypic dorsal striatal slices. Amino Acids. 2000;19:23–31. doi: 10.1007/s007260070030. [DOI] [PubMed] [Google Scholar]
- 15.Yuan SL, Huang RM, Wang XJ, Song Y, Huang GQ. Reversing effect of Tanshinone on malignant phenotypes of human hepatocarcinoma cell line. World J Gastroenterol. 1998;4:317–319. doi: 10.3748/wjg.v4.i4.317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.He SW, Shen KQ, He YJ, Xie B, Zhao YM. Regulatory effect and mechanism of gastrin and its antagonists on colorectal carcinoma. World J Gastroenterol. 1999;5:408–416. doi: 10.3748/wjg.v5.i5.408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Feng DY, Zheng H, Tan Y, Cheng RX. Effect of phosphorylation of MAPK and Stat3 and expression of c-fos and c-jun proteins on hepatocarcinogenesis and their clinical significance. World J Gastroenterol. 2001;7:33–36. doi: 10.3748/wjg.v7.i1.33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wang X, Wang BR, Zhang XJ, Xu Z, Ding YQ, Ju G. Evidences for vagus nerve in maintenance of immune balance and transmission of immune information from gut to brain in STM-infected rats. World J Gastroenterol. 2002;8:540–545. doi: 10.3748/wjg.v8.i3.540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Omote K, Kawamata T, Kawamata M, Namiki A. Formalin-induced nociception activates a monoaminergic descending inhibitory system. Brain Res. 1998;814:194–198. doi: 10.1016/s0006-8993(98)01086-5. [DOI] [PubMed] [Google Scholar]
- 20.Traub RJ, Murphy A. Colonic inflammation induces fos expression in the thoracolumbar spinal cord increasing activity in the spinoparabrachial pathway. Pain. 2002;95:93–102. doi: 10.1016/s0304-3959(01)00381-5. [DOI] [PubMed] [Google Scholar]
- 21.Liu XJ, Yang L, Mao YQ, Wang Q, Huang MH, Wang YP, Wu HB. Effects of the tyrosine protein kinase inhibitor genistein on the proliferation, activation of cultured rat hepatic stellate cells. World J Gastroenterol. 2002;8:739–745. doi: 10.3748/wjg.v8.i4.739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Harris JA. Using c-fos as a neural marker of pain. Brain Res Bull. 1998;45:1–8. doi: 10.1016/s0361-9230(97)00277-3. [DOI] [PubMed] [Google Scholar]
- 23.Leão RM, Von Gersdorff H. Noradrenaline increases high-frequency firing at the calyx of Held synapse during development by inhibiting glutamate release. J Neurophysiol. 2002;87:2297–2306. doi: 10.1152/jn.2002.87.5.2297. [DOI] [PubMed] [Google Scholar]
- 24.Al-Adawi S, Dawe GS, Bonner A, Stephenson JD, Zarei M. Central noradrenergic blockade prevents autotomy in rat: implication for pharmacological prevention of postdenervation pain syndrome. Brain Res Bull. 2002;57:581–586. doi: 10.1016/s0361-9230(01)00747-x. [DOI] [PubMed] [Google Scholar]
- 25.Li P, Zhuo M. Cholinergic, noradrenergic, and serotonergic inhibition of fast synaptic transmission in spinal lumbar dorsal horn of rat. Brain Res Bull. 2001;54:639–647. doi: 10.1016/s0361-9230(01)00470-1. [DOI] [PubMed] [Google Scholar]
- 26.Devilbiss DM, Waterhouse BD. Norepinephrine exhibits two distinct profiles of action on sensory cortical neuron responses to excitatory synaptic stimuli. Synapse. 2000;37:273–282. doi: 10.1002/1098-2396(20000915)37:4<273::AID-SYN4>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- 27.Ma J, Qiao JT, Dafny N. Opiate-like substances mediate norepinephrine-induced but not serotonin-induced antinociception at spinal level: reevaluation by an electrophysiological model of formalin test in rats. Life Sci. 2001;69:969–976. doi: 10.1016/s0024-3205(01)01191-2. [DOI] [PubMed] [Google Scholar]
- 28.Potts JT, Lee SM, Anguelov PI. Tracing of projection neurons from the cervical dorsal horn to the medulla with the anterograde tracer biotinylated dextran amine. Auton Neurosci. 2002;98:64–69. doi: 10.1016/s1566-0702(02)00034-6. [DOI] [PubMed] [Google Scholar]
- 29.Pérez H, Ruiz S, Laurido C, Hernández A. Locus coeruleus-mediated inhibition of chemosensory responses in the rat nucleus tractus solitarius is mediated by alpha2-adrenoreceptors. Neurosci Lett. 1998;249:37–40. doi: 10.1016/s0304-3940(98)00387-5. [DOI] [PubMed] [Google Scholar]
- 30.Xia PY, Zheng J, Zhou H, Pan WD, Qin XJ, Xiao GX. Relationship between lymphocyte apoptosis and endotoxin translocation after thermal injury in rats. World J Gastroenterol. 2002;8:546–550. doi: 10.3748/wjg.v8.i3.546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Meng AH, Ling YL, Zhang XP, Zhang JL. Anti-inflammatory effect of cholecystokinin and its signal transduction mechanism in endotoxic shock rat. World J Gastroenterol. 2002;8:712–717. doi: 10.3748/wjg.v8.i4.712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Diao TJ, Yuan TY, Li YL. Immunologic role of nitric oxide in acute rejection of golden hamster to rat liver xenotransplantation. World J Gastroenterol. 2002;8:746–751. doi: 10.3748/wjg.v8.i4.746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rabe C, Pilz T, Klostermann C, Berna M, Schild HH, Sauerbruch T, Caselmann WH. Clinical characteristics and outcome of a cohort of 101 patients with hepatocellular carcinoma. World J Gastroenterol. 2001;7:208–215. doi: 10.3748/wjg.v7.i2.208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.He XS, Su Q, Chen ZC, He XT, Long ZF, Ling H, Zhang LR. Expression, deletion [was deleton] and mutation of p16 gene in human gastric cancer. World J Gastroenterol. 2001;7:515–521. doi: 10.3748/wjg.v7.i4.515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lima D, Albino-Teixeira A, Tavares I. The caudal medullary ventrolateral reticular formation in nociceptive-cardiovascular integration. An experimental study in the rat. Exp Physiol. 2002;87:267–274. doi: 10.1113/eph8702354. [DOI] [PubMed] [Google Scholar]
- 36.Acuña-Goycolea C, Fuentealba P, Torrealba F. Anatomical substrate for separate processing of ascending and descending visceral information in the nucleus of the solitary tract of the rat. Brain Res. 2000;883:229–232. doi: 10.1016/s0006-8993(00)02845-6. [DOI] [PubMed] [Google Scholar]
- 37.Kwon YB, Kang MS, Han HJ, Beitz AJ, Lee JH. Visceral antinociception produced by bee venom stimulation of the Zhongwan acupuncture point in mice: role of alpha(2) adrenoceptors. Neurosci Lett. 2001;308:133–137. doi: 10.1016/s0304-3940(01)01989-9. [DOI] [PubMed] [Google Scholar]