

Abstract
A violacein-producing bacterial strain was isolated and identified as a relative of Duganella violaceinigra YIM 31327 based upon phylogenetic analyses using the 16S rRNA, gyrB and vioA gene sequences and a fatty acid methyl ester (FAME) analysis. This new strain was designated D. violaceinigra str. NI28. Although these two strains appear related based upon these analyses, the new isolate was phenotypically different from the type strain as it grew 25% faster on nutrient media and produced 45-fold more violacein. When compared with several other violacein producing strains, including Janthinobacterium lividum, D. violaceinigra str. NI28 was the best violacein producer. For instance, the crude violacein yield with D. violaceinigra str. NI28 was 6.0 mg/OD at 24 hours, a value that was more than two-fold higher than all the other strains. Finally, the antibacterial activity of D. violaceinigra str. NI28 crude violacein was assayed using several multidrug resistant Staphylococcus aureus. Addition of 30 μM crude violacein led to a 96% loss in the initial S. aureus population while the minimum inhibitory concentration was 1.8 μM. Consequently, this novel isolate represents a phenotypic variant of D. violaceinigra capable of producing much greater quantities of crude violacein, an antibiotic effective against multidrug resistant S. aureus.
Violacein is a bisindole derived from the condensation of two molecules of tryptophan1. This compound is vibrant purple in color and is known to possess a broad biological activity, including performing as an antitumor2, antifungal3 and antiviral4. Although not tested within humans to date, violacein has been found to have no adverse impact on mice when administered at a concentration of 1 mg/kg2. It has recently garnered more attention, however, due to its antibacterial activity against Staphylococcus aureus5,6. Tests with this pathogen found that violacein is capable of both acting as a bacteriostatic and as a bacteriocidal agent against S. aureus depending on the concentration used7. Moreover, a recent study demonstrated that violacein works synergistically with other antibiotics, leading to significantly lower minimum inhibitory concentrations for S. aureus, Klebsiella pneumonia and Pseudomonas aeruginosa6. All three of these strains are included in the ESKAPE pathogen grouping, a list of multidrug resistant bacteria that are becoming more prominent within healthcare environments and nosocomial infections8,9. As such, research into the isolation and characterization of violacein producing bacteria, the cloning and heterogeneous expression of the vioABCDE genes and the fermentative production of violacein has recently blossomed with several key studies reported within the last couple of years10,11,12,13.
Violacein is produced by numerous bacterial strains spanning various genera, including Chromobacter14,15, Pseudoalteromonas16,17, Janthinobacterium18,19 and Duganella20. Moreover, violacein producing bacterial strains have been isolated from diverse environmental locales. Janthinobacterium18,21,22, for instance, was isolated from a glacier, Chromobacteria from a river14,23, Duganella from agricultural and forest soils24,25 and Collimonas from the sea26. In this study, we report on the isolation and initial characterization of a natural soil isolate, Duganella violaceinigra str. NI28, obtained from a temperate forest soil sample taken near Ulsan, South Korea. This strain produces violacein at much higher rate and levels than the type strain D. violaceinigra YIM 3132724,27.
Results
Isolation and Identification of D. violaceinigra str. NI28
Various natural bacterial isolates from a forest soil sample were grown on nutrient agar (NA) and a single colony that had a dark purple hue, suggesting that this strain produced the bisindole violacein, was selected for further characterization. Production of crude violacein by this strain was demonstrated using HPLC analysis (Fig. 1). When the crude violacein extracted from our new isolate was compared with a commercial preparation of crude violacein from Janthinobacterium lividum (Sigma-Aldrich, USA), they were basically indistinguishable from each other.
Figure 1. HPLC analysis of the violacein extracted from cultures of D. violaceinigra str.
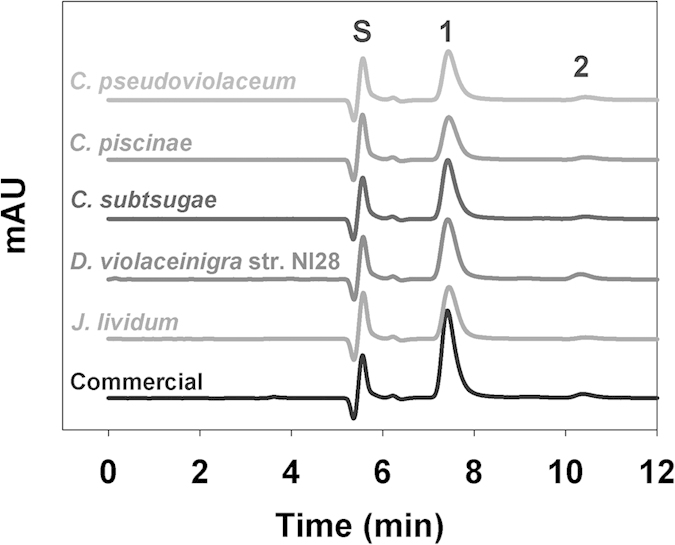
NI28 and several of the strains employed in this study, showing the presence of violacein (Peak #1–7.5 min) and a second peak (Peak #2–10.3 min) that is presumed to be deoxyviolacein. The solvent front (Peak S) is seen at 5.4 min. A plot generated using a commercially available violacein extracted from Janthinobacterium lividum is also provided for comparison and includes both Peak #1 and Peak #2.
Identification of this strain was next performed using the sequences obtained from three gene loci (16S rRNA, gyrB and vioA), with each confirming that it was closely related to Duganella violaceinigra YIM 31327 (Figs 2 and 3). The level of sequence similarities between NI28 and D. violaceinigra YIM 31327 were 98.8% for 16S rRNA gene, 95.9% for gyrB, and 88.8% for vioA, thus indicating that NI28 is close, but not identical to Duganella violaceinigra YIM 31327. This relationship was further supported by a fatty acid methyl ester (FAME) analysis and comparison of this strain and the type strain, Duganella violaceinigra YIM 31327, which showed only minor differences between the two strains (Table 1). Based upon the phylogenetic and FAME analyses, this strain was designated D. violaceinigra str. NI28 and deposited within the Korea Agricultural Culture Collection (www.genebank.go.kr) (KACC 91951P).
Figure 2. Phylogenetic tree using the 16S rRNA gene sequence.

The numbers at nodes indicate levels of bootstrap support (%) based on 1,000 resampled dataset. The bars corresponds to 0.01 or 0.05 substitutions per nucleotide. In parentheses are the nucleotide sequence accession numbers of corresponding strains.
Figure 3. Phylogenetic analysis of the new isolate using the (A) gyrB and (B) vioA gene sequences.

The numbers at nodes indicate levels of bootstrap support (%) based on 1,000 resampled dataset. The bars corresponds to 0.01 or 0.05 substitutions per nucleotide. The nucleotide sequence accession numbers of the corresponding strains are listed in theparentheses.
Table 1. Fatty acid methyl ester (FAME) comparison between D. violaceinigra str. NI28 and D. violaceinigra YIM 31327.
| Fatty Acid (%) | NI28 | YIM 31327 |
|---|---|---|
| Straight Chain | ||
| C10:0 | − | 0.4 |
| C12:0 | 5.05 | 6.54 |
| C14:0 | 2.99 | 1.04 |
| C16:0 | 33.28 | 31.42 |
| Branched | ||
| Iso-C10:0 | 0.12 | − |
| Unsaturated | ||
| C14:1 | − | 0.26 |
| C18:1 | 3.8 | 3.69 |
| Hydroxy | ||
| C10:0 | − | 3.82 |
| C12:0 | 2.66 | 1.74 |
| C14:0 | 3.41 | 3.6 |
| Sum in Feature 3 | 48.68 | 47.49 |
*Summed features represent two or three fatty acids that cannot be separated by the Microbial Identification System. The fatty acids included in Feature 3 consist of C16 :1ω7c and/or iso-C15:0 2-OH.
D. violaceinigra str. NI28 is Phenotypically Variant from D. violaceinigra YIM 31327
Although both D. violaceinigra str. NI28 and D. violaceinigra YIM 31327 were isolated from forest soils and are related genetically based upon the above results, differences were readily apparent between them. For instance, D. violaceinigra str. NI28 was found to have trypsin activity using the API ZYM Kit (bioMerieux, France) while D. violaceinigra YIM 31327 was negative for this protease. Furthermore, D. violaceinigra str. NI28 grew remarkably well and produced a significant amount of violacein when cultured on NA (Fig. 4A). This was in stark contrast with D. violaceinigra YIM 31327, which grew slower and was much less proficient at producing violacein. Figure 4A shows that colonies of D. violaceinigra str. NI28 were larger and already producing violacein after 24 hours of growth while those of D. violaceinigra YIM 31327 were smaller and still pasty in color. Only after 60 hours did the D. violaceinigra YIM 31327 colonies achieve a similar size and hue as 24 hour-old the D. violaceinigra str. NI28 colonies (Fig. 4A).
Figure 4.

D. violaceinigra str. NI28 grows faster than D. violaceinigra YIM 31327. (A) Image of each strain grown on NB agar plates for 24 and 60 hours. The upper and bottom regions are from the same plate. Note the more rapid colony development and violacein production by D. violaceinigra str. NI28. (B) Growth of both strains in NB media confirming that D. violaceinigra str. NI28 grows faster. The data for each strain was obtained from three independent cultures.
Both of these findings were further evidenced in liquid cultures. As with the colonies, D. violaceinigra YIM 31327 was slower to grow in NB liquid media (Fig. 4B). The doubling time for D. violaceinigra YIM 31327 was 71 minutes based upon the logarithmic growth stage between 3 and 7 hours. In contrast, D. violaceinigra str. NI28 doubled every 53.7 minutes, a value that is 25% faster than D. violaceinigra YIM 31327. Not only did the newly isolated D. violaceinigra str. NI28 grow faster but the optical density after 24 hours was significantly higher (2.4-fold), as shown in Figs 4B and 5A. We also noticed that D. violaceinigra str. NI28 tended to form flocs when grown in liquid cultures while D. violaceinigra YIM 31327 cells generally remained suspended (data not shown). It is not clear what benefit D. violaceinigra str. NI28 gains from forming these aggregates, but it was reported that flocs may help protect against predation28.
Figure 5. Growth and violacein production by several different bacterial strains.
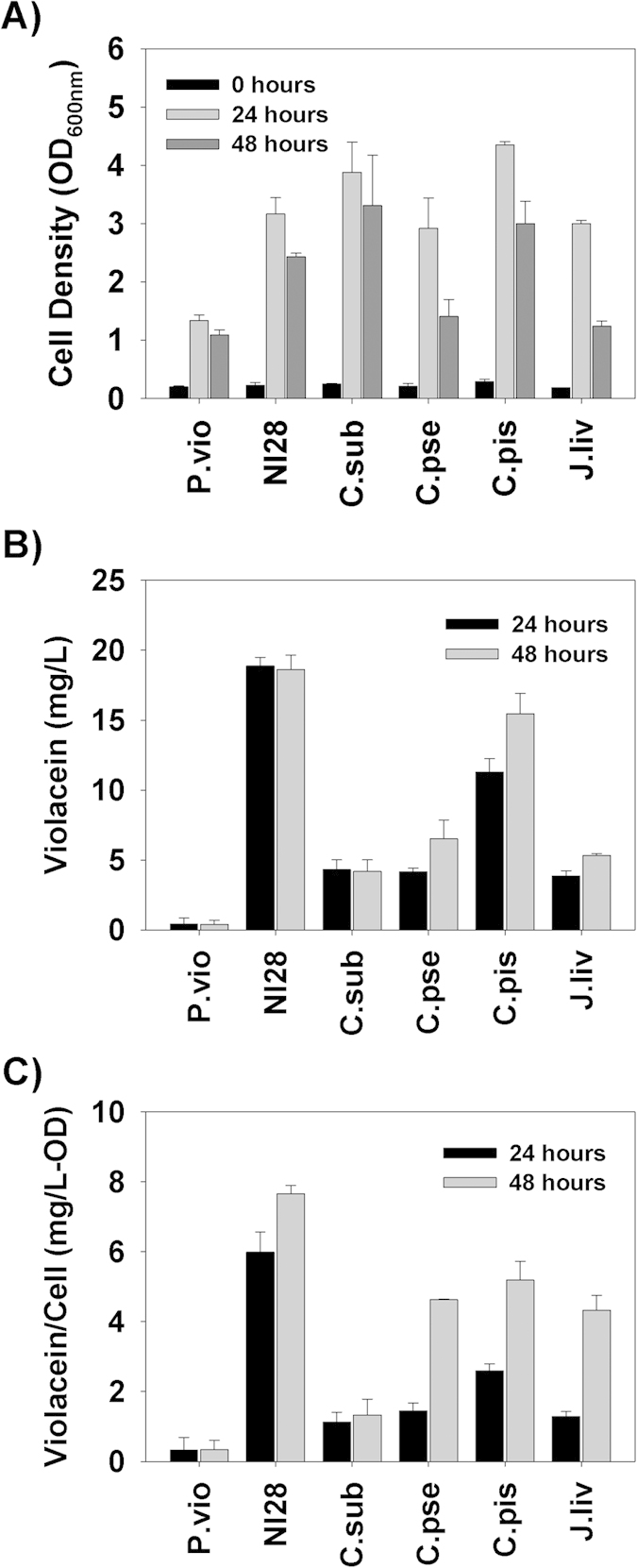
(A) Optical density values for each culture at 0, 24 and 48 hours. (B) Violacein concentration within each culture at 24 and 48 hours, showing the rapid and high level violacein generation by D. violaceinigra str. NI28 as compared with the other strains. (C) The relative concentration of violacein based upon the culture density, illustrating that D. violaceinigra str. NI28 gave the greatest yields.
D. violaceinigra str. NI28 is a Prolific Violacein Producer
All three of these growth characteristics (rate, yield and floc formation) may contribute to the higher violacein production seen with this new strain. As shown in Fig. 5B, D. violaceinigra str. NI28 was a much more proficient violacein producer than D. violaceinigra YIM 31327 under these conditions, generating as much as 18.9 mg/L of crude violacein after 24 hours. Further incubation for an additional 24 hours did not improve upon this yield with 18.6 mg/L from the two day culture. D. violaceinigra YIM 31327, on the other hand, produced only 0.42 mg/L during the first 24 hours, and this dropped to 0.38 mg/L after 48 hours. Considering both time points, the new strain produced approximately 45-fold more violacein than the type strain.
Although D. violaceinigra str. NI28 is clearly better than D. violaceinigra YIM 31327 at producing violacein, the literature also lists several other violacein producing strains, including Chromobacter species and Janthinobacterium lividum (Table 2). Consequently, we cultivated each of these strains in NB media and determined both the optical density (OD) and crude violacein yields after 24 and 48 hours (Fig. 5). The 24 and 48 hour D. violaceinigra str. NI28 optical densities were similar with most of the other violacein producing strains but both C. piscinae and C. subtsugae grew to higher ODs (Fig. 5A). However, of all the strains tested, the new isolate gave the highest crude violacein yields, particularly when grown for 24 hours (Fig. 5B). This figure also shows that it was one of only two strains, other than D. violaceinigra YIM 31327, which reached a maximum violacein concentration in 24 hours. This rapid and high level production by D. violaceinigra str. NI28 in NB clearly sets this strain apart from the other strains, particularly its closest relative, D. violaceinigra YIM 31327, which was the poorest violacein producer tested in this study.
Table 2. Strains used in this study.
| Strains | Description | Referencea |
|---|---|---|
| Duganella violaceinigra str. NI28 | Violacein producer | This study |
| Duganella violaceinigra | Violacein producer | YIM 31327 |
| Janthinobacterium lividum | Violacein producer | ATCC 12473 |
| Chromobacterium pseudoviolaceum | Violacein producer | ATCC 7461 |
| Chromobacterium piscinae | Violacein producer | LMG 3947 |
| Chromobacterium subtsugae | Violacein producer | DSM 17043 |
| Staphylococcus aureus | Type strain | ATCC 25923 |
| Staphylococcus aureus | Multidrug Resistant (MRSA) | Clinical Isolate |
| Staphylococcus aureus | − | CCARM 0201 |
| Staphylococcus aureus | Multidrug Resistant (MRSA) | CCARM 3090 |
| Staphylococcus aureus | Multidrug Resistant (MRSA) | CCARM 3840 |
| Klebsiella pneumoniae | Human pathogen | ATCC 13883 |
| Pseudomonas aeruginosa PAO1 | Human pathogen |
aYIM (Yunnan Institute of Microorganisms); ATCC (American Type Culture Collection); LMG (Belgian Coordinated Collection of Microorganisms); DSM (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH); CCARM (Culture Collection of Antimicrobial Resistant Microbes).
When the quantity of violacein produced by all six strains was normalized using the OD, i.e., the specific productivity, the newly isolated D. violaceinigra str. NI28 gave the best results (Fig. 5C). This was particularly true for the 24 hour sample where the specific productivity was 6.0 mg crude violacein/OD, a value that was more than twice that obtained for the next closest strain, C. piscinae (2.6 mg crude violacein/OD). In comparison, the specific productivity from the type strain, D. violaceinigra YIM 31327, was the lowest of all the strains tested (0.33 and 0.34 mg/OD at 24 and 48 hours, respectively). As such, D. violaceinigra str. NI28’s high level production of violacein implies that this strain contains more violacein per cell than the other strains, a finding which may contribute to making downstream purification of violacein easier.
Duganella violaceinigra str. NI28 Violacein is Bacteriocidal Towards Staphylococcus aureus
Several recent articles highlighted the activity of violacein against the human pathogen Staphylococcus aureus and showed that a crude violacein concentration of 17 μM was capable of inhibiting growth of this strain5,6. Similarly, we tested the activity of D. violaceinigra str. NI28 violacein extracts with two S. aureus isolates that were obtained from culture repositories (ATCC 25923 and the CCARM strains) and from a patient infected with a multidrug-resistant S. aureus (Clinical) (Table 3). As shown in Fig. 6, the crude violacein prepared from D. violaceinigra str. NI28 was capable of blocking growth of both S. aureus strains when added to a final concentration of 15 μM, or approximately 5 mg/L. Moreover, this concentration led to a 70% loss in viability after 24 hours when compared with the initial S. aureus colony-forming unit (CFU) values while higher concentrations led to comparably greater losses in viability. For instance, 60 μM violacein reduced both S. aureus populations by an average of 99.5% when compared with the initial CFU numbers.
Table 3.
| Strain | Vana | Rifa | Cipa | Clina | Oxaa | Erya | Genta | Toba |
|---|---|---|---|---|---|---|---|---|
| S. aureus | ||||||||
| ATCC 25923 | 19–21(S)b | 32–36(S) | 30–31(S) | 27–30(S) | 22–25(S) | 27–30(S) | 24–27(S) | 26–27(S) |
| Clinical Isolate | 21–22(S) | 0(R) | 11–18(R) | 0(R) | 0(R) | 0(R) | 0(R) | 0(R) |
| CCARM 0201 | 18–19(S) | 31–34(S) | 25–30(S) | 23–28(S) | 25–27(S) | 21–30(S) | 22–24(S) | 20–23(S) |
| CCARM 3090 | 19–21(S) | 36–38(S) | 0(R) | 0(R) | 0(R) | 0(R) | 26–27(S) | 26–28(S) |
| CCARM 3840 | 19–24(S) | 33–40(S) | 0(R) | 0(R) | 0(R) | 0(R) | 0(R) | 0(R) |
| K. pneumoniae ATCC 13883 | NDc | ND | 35–37(S) | ND | ND | ND | 22–25(S) | 24–25(S) |
| P. aeruginosa PAO1 | ND | ND | 33–35(S) | ND | ND | ND | 23–24(S) | 29–30(S) |
Antibiotic susceptibility results. Tests were performed using disc-diffusion plates with the results listed representing the range of diameters (mm) measured for the inhibitory zones from three independent tests. Strains showing resistance are listed in bold.
aThe antibiotics tested were vancomycin (Van), rifampin (Rif), ciprofloxacin (Cip), clindamycin (Clin), oxacillin (Oxa), erythromycin (Ery), gentamycin (Gent) and tobramycin (Tob).
bRange of diameters (mm) seen for the inhibitory zones. (S) – susceptible; (R) – resistant.
cNot done.
Figure 6. Inhibition and killing of Staphylococcus aureus by D. violaceinigra str.
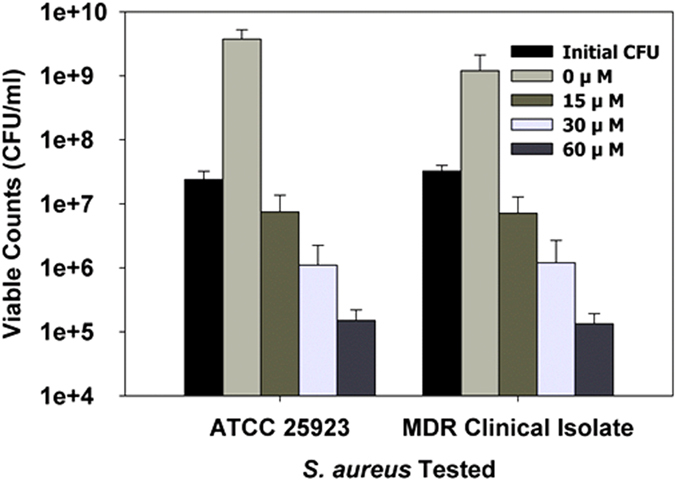
NI28 violacein. Two S. aureus strains were tested: ATCC 25923 and a clinical isolate. The initial CFU was determined just prior to addition of the violacein. The viability of the other samples was determined after a 24 hour exposure to the violacein at the concentrations listed, showing that the addition of 15 μM or greater violacein was bactericidal towards both pathogenic strains. The samples were prepared so that each had the same amount of DMSO (1.3%) added. The concentrations listed are based upon the commercial standard.
We next tested the efficacy of the D. violaceinigra str. NI28 crude violacein against a selection of bacterial strains, including several multidrug resistant S. aureus strains that have diverse antibacterial resistance footprints according to the EUCAST breakpoints (Table 3). Table 4 lists the minimal inhibitory concentrations (MICs) of violacein found for each of the strains tested in this study. Growth of all S. aureus cultures was inhibited by a violacein concentration of 15 μM (Table 4) regardless of the antibiotic resistant nature of the pathogen. Furthermore, MIC tests performed using Pseudomonas aeruginosa and Klebsiella pneumoniae found both of these pathogens were resistant to violacein. Parallel tests performed with a commercially available crude violacein from J. lividum gave identical results (Table 4), demonstrating that the bioactivity of the crude violacein from D. violaceinigra str. NI28 against these pathogens is equivalent to that of the commercially available J. lividum crude violacein.
Table 4. Minimum inhibitory concentration results.
| Strain | Commercial Violacein | NI28 Violacein | Commercial Violacein | NI28 Violacein |
|---|---|---|---|---|
| Mueller-Hinton Media | Modified M9 Media | |||
| S. aureus | ||||
| ATCC 25923 | 15 μM | 15 μM | 1.8 μM | 1.8 μM |
| Clinical Isolate | 15 μM | 15 μM | 0.9 μM | 1.8 μM |
| CCARM 0201 | 15 μM | 15 μM | 1.8 μM | 1.8 μM |
| CCARM 3090 | 15 μM | 15 μM | 1.8 μM | 1.8 μM |
| CCARM 3840 | 15 μM | 15 μM | 0.9 μM | 1.8 μM |
| K. pneumoniae ATCC 13883 | >30 μM | >30 μM | >30 μM | >30 μM |
| P. aeruginosa PAO1 | >30 μM | >30 μM | >30 μM | >30 μM |
The concentration listed is that which showed no growth after 24 hours in the resazurin microplate tests using the modified M9 media. The highest crude violacein concentration tested was 60 μM.
Discussion
As a bacterial secondary metabolite, violacein has garnered renewed interest lately. A vibrant violet-hued bisindole derivative of tryptophan, violacein has been shown to have multiple biological activities, including as an anticancer2,13 and antibiotic, particularly against S. aureus. It is this multifaceted character of violacein, and the fact that resistance to this antibiotic has not yet been described, that makes it an attractive product for bioprocesses.
This study reports on the isolation and characterization of a novel high-level violacein-producing bacterial strain. This strain was identified as Duganella using both FAME and phylogenetic analyses and found to be similar based upon these tests with the type strain D. violaceinigra YIM 31327, which also produces violacein. However, when these two strains were compared with each other, we found that they were quite distinct from each other. For instance, the new strain grew faster and produced significantly more violacein (45-fold) than D. violaceinigra YIM 31327. Moreover, this new isolate also has a tendency to form flocs, another attribute that has been noted as being more economical when harvesting microbial biomass29. When this strain, D. violaceinigra str. NI28, was compared with several other well-known violacein producers, it grew faster, produced more violacein and had the best yields per biomass. Both the rapid, high-level production by D. violaceinigra str. NI28 and its higher concentration within the cells are clearly advantageous and desirable from an economic viewpoint since the first would reduce the time needed for production of violacein while the second minimizes the biomass needed for its subsequent extraction and purification. These characteristics of D. violaceinigra str. NI28 make it a potentially attractive and alternative strain for the commercial production of violacein.
When the violacein from D. violaceinigra str. NI28 cultures was purified and analyzed, we found that it was similar with a commercially available preparation from J. lividum, as well as that extracted from the various strains employed in this study. As noted above, this bisindole is quite effective against S. aureus and, as such, this pathogen was selected for this study due to its overwhelming presence within nosocomial infections30,31 and its broad resistance to antibiotics32. Previous studies reported the minimum inhibitory concentration (MIC) of violacein was 5.7 μg/ml, or approximately 17 μM, with S. aureus6. The preparation from D. violaceinigra str. NI28 was equally potent with an MIC of 5 μg/ml, or approximately 15 μM. This concentration was not only inhibitory but also bactericidal as it led to a 69% reduction in the viability with the ATCC strain and a 78% decrease with the clinical isolate. Both of these findings clearly illustrate that the crude violacein purified from cultures of D. violaceinigra str. NI28 is capable of not only inhibiting S. aureus growth but also killing this pathogen.
Moreover, the multidrug resistant nature of the S. aureus strains did not contribute to their survival when exposed to violacein. Tests using a total of five S. aureus strains with different antibiotic resistance footprints, showing that several were multidrug resistant (Table 3), found that each was equally sensitive to the crude violacein from D. violaceinigra str. NI28, with MICs of 15 μM (Table 4). These values are basically identical with that reported by Subramaniam et al. (2014), where they found the MIC for S. aureus to be 5.7 μg/ml (17 μM). When we performed further MIC tests in a modified M9 media, the effective concentration dropped to only 0.9 or 1.8 μM, a value that is significantly lower than those found with the Mueller-Hinton media and is likely attributable to the nutrient poor nature of the medium used. Similar impacts of the media richness and bacterial responses has been documented previously33, where the authors found the use of a nutrient poor medium made the bacteria more susceptible to chemicals. Consequently, the use of a minimal media within this study increased the susceptibility of the bacteria to violacein and does not indicate a greater activity against these pathogens. Further evidence of this is seen in the similar MICs obtained with the commercially available J. lividum crude violacein, which was also significantly lower (Table 4). The HPLC analyses of these two crude violacein preparations, shown in Fig. 1, found them to be similar. The second peak was not identified but is presumed to be deoxyviolacein since this compound is also known to be generated alongside violacein. Although it may also play a role and contribute to S. aureus inhibition, a recent study found that deoxyviolacein was less effective against this pathogen than violacein34.
Two additional pathogens were also tested, P. aeruginosa PAO1 and K. pneumoniae, but neither of these was sensitive to either of the violacein samples (Table 4). This was not unexpected as several articles previously highlighted the tendency for Gram-positive strains to be more sensitive than Gram-negative strains6,25. These findings suggest that the presence of the outer membrane within these bacteria provides protection against violacein and its activity and is an area for further evaluation in a later study.
In conclusion, a violacein-producing strain was isolated from a forest soil sample in Ulsan, South Korea. This strain, D. violaceinigra str. NI28, produces violacein at quicker and at higher levels than other comparable strains, particularly the type strain D. violaceinigra YIM 31327, when grown under the conditions used in this study. Although the violacein concentrations produced in this study are low, a group recently reported achieving as much as 0.82 g/L with C. violaceum by optimizing the conditions for fermentation and the media used35. As D. violaceinigra str. NI28 is a new strain and, based upon the violacein production and yields found in this study, is generally equivalent to or better than the other strains tested, expanding its growth and violacein production through similar techniques would likely increase the yields. Moreover, other recent studies have shown the successful expression of the vioABCDE operon within other bacterial hosts to produce even higher levels of violacein. The characteristics of D. violaceinigra str. NI28 make its vioABCDE operon an interesting addition to these studies and, as such, cloning of these genes for their heterologous expression may likewise be beneficial.
Methods
Isolation of Duganella violaceinigra str. NI28
The novel strain characterized in this study, i.e., Duganella violaceinigra str. NI28, was isolated using soil obtained from a temperate forest nearby the campus. For this, a 50 g sample of soil was mixed with 500 ml of sterile tap water and shaken at 30 °C for 2 hours. Afterwards, the soil particulates were centrifuged at 500 × g for 5 minutes and the supernatant sampled. Aliquots (100 μl) of this solution were spread out on nutrient agar (NA) (1.6% agar; Difco, USA) plates. A total of forty plates were prepared and these were then incubated at 30 °C for 48 hours. A single colony showing a violet hue on one of the plates was selected for further evaluation.
Bacterial Strains and Growth
The bacterial strains used in this study are listed in Table 2. All of the strains were grown from −80 °C freezer stocks that were prepared individually. For the violacein producing strains, the stocks were prepared by adding DMSO (Sigma-Aldrich, USA) to a final concentration of 10%. The Staphylococcus aureus strains were stored using glycerol (Sigma-Aldrich, USA) at a final concentration of 25%.
Each violacein producing strain was struck out on NA plates and grown at 30 °C overnight. A single colony was transferred to a sterile 15 ml conical tube (SPL, Korea) containing 3 ml of nutrient broth (Difco, USA). This culture was grown at 30 °C and 250 rpm overnight. To 20 ml of fresh NB medium in a 100 ml baffled flask to increase aeration, 2 ml (10% v/v) of the overnight culture was added and this was again shaken at 30 °C and 250 rpm for 48 hours. At set times during this cultivation (24 and 48 hours), aliquots were taken to determine the culture optical density and violacein concentration as described below. For the growth curves shown in Fig. 4, 100 ml cultures were cultivated within 250 ml flasks. All the other strains were grown using the same protocols except at 37 °C and in TSB media (Difco, USA). Other media was used for the antibiotic resistance characterization and minimum inhibitory concentration determination as described below.
Identification of D. violaceinigra str. NI28 by 16S using phylogenetic markers
The genus of isolate NI28 was identified using three genes, namely 16S rRNA, gyrB and vioA. The primers for PCR amplification of the genes were 27f (5′-AGA GTT TGA TCM TGG CTC AG-3′) and 1492r (5′-GGT TAC CTT GTT ACG ACT T-3′) for the 16S rRNA gene, Up-1G (5′-YGC SGG CGG YAA GTT CGA-3′) and Up-2G (5′-CCR TCG ACG TCV GCR TCG GT-3′) for gyrB36, and VPA3 (5′-CCR CAG CTS CAY CCG CAT TTC CAG-3′) and VPA4 (5′-CAG GCY GCC CTC CAT CCA GCC RCA-3′) for vioA26. The PCR conditions for the gyrB gene were: 95 °C for 5 minutes followed by 35 cycles of 95 °C for 30 s, 60 °C for 15 s and 72 °C for 90 s. The final extension was at 72 °C for 5 minutes. The vioA gene was amplified in the same manner but with an annealing temperature of 62 ºC. The phylogenetic analysis between NI28 and the related taxa including Duganella was performed using MEGA 6.037. Neighbor-joining trees were inferred based on Jukes-Cantor evolutionary distances for each gene. The trees were evaluated using bootstrap analysis with 1,000 resampled datasets. The sequences of each gene from strain NI28 were deposited within the GenBank database under the accession numbers (KM087998, KM087999 and KM088000).
Fatty Acid Methyl Ester Preparation and Analysis
Colonies of D. violaceinigra str. NI28 and D. violaceinigra YIM 31327 grown on NA plates were used for this analysis. To prepare the samples, several colonies were transferred to a test tube and 1 ml of Reagent #1 (45 g NaOH (ACS Grade, Sigma-Aldrich, USA), 150 ml HPLC Grade methanol (Sigma-Aldrich, USA) and 150 ml deionzed distilled water) was added. After vortexing to suspend the bacteria, this sample was placed at 100 °C for 5 minutes, vortexed once more and then incubated at 100 °C once more for 25 minutes. Afterwards, the tubes and samples were cooled down in a room temperature water bath. To initiate methylation of the fatty acids, 2 ml of Reagent #2 (325 ml 6 N HCl (Sigma-Aldrich, USA) mixed with 275 ml HPLC Grade methanol (Sigma-Aldrich, USA)) was added to each tube, which was vortexed and then incubated at 80 °C for 10 minutes. Afterwards, the tubes were quickly cooled by gently shaking them in a room temperature water bath. To each tube, 1.25 ml Reagent #3 (prepared using an equal volume of HPLC Grade hexane (Sigma-Aldrich, USA) and HPLC Grade methyl tert-butyl ether (Sigma-Aldrich, USA)) was added and the contents were gently mixed end-over-end for 10 minutes using a rotator. The aqueous phase was removed from the tubes and discarded. To remove any free fatty acids, the samples were washed with 3 ml of Reagent #4 (10.8 g ACS Grade sodium hydroxide in 900 ml deionized distilled water) by gently mixing them as above for 5 minutes. Once more, the aqueous phase was removed and discarded. The organic phase was used for gas chromatography (GC) analysis. For the GC analysis, the column was an Ultra 2 (25 m, 0.2 mm, Agilent, USA). The protocol used was the same as used previously for the D. violaceinigra type strain24.
Phenotypic Assessment of D. violaceinigra str. NI28
A comparative phenotypic analysis of D. violaceinigra str. NI28 and YIM 31327 was performed using the API 20NE and API ZYM test kits (bioMerieux, France) according to the manufacturer’s suggested protocol.
Extraction and HPLC Analysis of the Violacein Produced by D. violaceinigra str. NI28
After growth for 24 and 48 hours, 1 ml of the culture was pelleted by centrifugation for one minute at 16,200 × g and room temperature. After aspirating off the media, the pellet was resuspended in an equal volume of ethanol and shaken at 37 °C and 250 rpm for 24 hours. The solution was then transferred to another centrifuge tube and the absorbance measured at 575 nm. This value was used to calculate the violacein concentration using an extinction coefficient of 73.4 L mg−1 cm−1 12. The crude violacein concentration within the ethanol extracts was determined using standards prepared with a crude violacein purchased from Sigma-Aldrich (USA, Cat. No. V9389).
The crude violacein sample was also analyzed using HPLC 1200 (Agilent, USA) as described previously12. For this, 10 μl samples of the ethanol extracted violacein were analyzed using a C-18 column (Hypersil ODS, 5 μm, 250 × 4.6 mm) at 30 ºC. The mobile phase was 50% denatured ethanol (HPLC Grade, Sigma-Aldrich, USA) and detection was performed at 575 nm using an Agilent 1260 Infinity ELSD. The violacein concentration within the ethanol extracts was determined as described above using crude violacein purchased from Sigma-Aldrich (USA, Cat. No. V9389) as a known standard. For the viability and MIC tests, the ethanol was dried off and the crude violacein powder solubilized within DMSO at a known concentration.
Antibiotic Disc Diffusion Assay
Each of the strains characterized is listed in Table 3. Each strain was grown on tryptone soy agar plates overnight and several colonies were transferred to 5 ml fresh sterile tryptone soy broth (TSB). They were then grown for overnight (250 rpm, 37 ºC), diluted into fresh sterile TSB (0.1%) and grown until the optical density (OD) was equivalent to a 0.5 McFarland standard, which was provided with the API tests used above. After reaching this OD, the culture was spread out on Mueller-Hinton agar plates using sterile, non-toxic cotton swabs. The plates were air-dried for 10 minutes before applying the antibiotic discs (BBL Sensi-Disc, BD, U.S.A.). The antibiotics tested were ciprofloxacin, clindamycin, erythromycin, gentamycin, oxacillin, rifampin, tobramycin and vancomycin. The zones of growth inhibition were determined after 18 hours of incubation at 37 °C for all the antibiotics except vancomycin and oxacillin, which were determined after 24 hours. Strain resistance was determined using the latest breakpoint tables available at the European Committee on Antimicrobial Susceptibility Testing (EUCAST) website (http://www.eucast.org/clinical_breakpoints/).
Violacein Killing of Staphylococcus aureus
The extracted crude violacein was concentrated by drying down multiple preparations under nitrogen gas and resuspending it in DMSO. The concentration was once more determined as described above.
Two different strains of S. aureus (ATCC 25923 and a clinical isolate) were used to evaluate the activity of the violacein extracted from P. sp. NI28 cultures. After growth overnight, the S. aureus cultures were diluted 1:100 into 3 ml M9 modified minimal media containing 1 g/L tryptone and 0.2% glucose. We chose this media as it is slightly modified from one that was published recently and used for MIC testing38. Violacein (M.W. 343.33) in DMSO was added to these cultures at a final concentration of 0, 15, 30 or 60 μM. The final DMSO concentration in all cultures was 1.3%. The cultures were then incubated at 37 °C and 250 rpm for 24 hours, after which the viable S. aureus numbers were determined using serial dilutions of the cultures that were spread out on TSB agar plates and grown overnight at 37 ºC.
Minimum Inhibitory Concentration Determination
The crude violacein samples were prepared as above and the commercial violacein was purchased from Sigma-Aldrich (USA). Determining the minimum inhibitory concentration was performed as described previously using resazurin39, with some modification. Briefly, the bacteria were grown overnight in cation adjusted Mueller-Hinton broth (10 mg/L of both Ca2+ Mg2+) medium at 37 °C and 250 rpm and then sub-cultured into fresh media and grown to an OD600nm of 0.1 under the same conditions. When they reached this OD, the cultures were diluted 1:100 into either cation adjusted Mueller-Hinton broth or modified M9 minimal media (containing 1 g/L tryptone and 0.2% glucose).
The media for the plate was prepared separately. Briefly, 3 ml of a sterile 0.2% resazurin solution prepared using distilled water was diluted into 12 ml of the modified M9 media. A 1.5 ml sample of this was then taken and crude violacein was added to a final concentration of 60 μM. Within the first row of the plate (row A), 200 μl of either cation adjusted Mueller-Hinton broth or modified M9 media containing 60 μM violacein was added. Each of the other wells had 100 μl of the same media containing resazurin added to them. The violacein containing media was then diluted two-fold sequentially from rows A-G but not into the final row (H). To the wells, 100 μl of the prepared bacterial cultures were added, giving a final cell concentration of 4 × 105 CFU/well, the highest violacein concentration of 30 μM and a resazurin concentration of 0.02% in each well. The final row (H) was used as a positive control to demonstrate that each bacterium was capable of growing. Each strain was tested independently in triplicate. The control wells had only the media and the violacein added to them to test for contamination. All the wells had a final DMSO concentration of 1%.
Data Analysis
Each of the experiments was performed in triplicate for error analysis. The standard deviations are presented as error bars in each graph where appropriate.
Additional Information
How to cite this article: Choi, S.Y. et al. High-level production of violacein by the newly isolated Duganella violaceinigra str. NI28 and its impact on Staphylococcus aureus. Sci. Rep. 5, 15598; doi: 10.1038/srep15598 (2015).
Acknowledgments
This work was supported by grants from the Korea Health Industry Development Institute (KHIDI) (Grant #HI13C13550000), the National Research Foundation of Korea (Grant No. 2013R1A1A2008590), and the Korea Institute for Advancement of Technology (KIAT) through the EUREKA program (Grant #N019800009). SBK also acknowledges support from the 2014 CNU Research Fund (Grant #2014-0784-01). We are all thankful for the support. We would also like to thank Dr. Sujin Lee, MD, at the Pusan National University Hospital-Yang San for the clinical isolate of Staphylococcus aureus isolate used in this study.
Footnotes
Author Contributions S.Y.C., S.L. and R.J.M. designed the study; S.Y.C., S.K. and S.L. performed the experiments; S.Y.C. and S.B.K. performed the phylogenetic analyses; S.Y.C., S.K. and R.J.M. wrote the paper. All authors reviewed the manuscript.
References
- Hoshino T., Kondo T., Uchiyama T. & Ogasawara N. Studies on the Biosynthesis of Violacein .1. Biosynthesis of Violacein - a Novel Rearrangement in Tryptophan-Metabolism with a 1,2-Shift of the Indole Ring. Agr Biol Chem Tokyo 51, 965–968 (1987). [Google Scholar]
- Bromberg N. et al. Growth inhibition and pro-apoptotic activity of violacein in Ehrlich ascites tumor. Chem Biol Int 186, 43–52, 10.1016/j.cbi.2010.04.016 (2010). [DOI] [PubMed] [Google Scholar]
- Brucker R. M. et al. Amphibian chemical defense: antifungal metabolites of the microsymbiont Janthinobacterium lividum on the salamander Plethodon cinereus. J Chem Ecol 34, 1422–1429, 10.1007/s10886-008-9555-7 (2008). [DOI] [PubMed] [Google Scholar]
- Andrighetti-Frohner C. R., Antonio R. V., Creczynski-Pasa T. B., Barardi C. R. & Simoes C. M. Cytotoxicity and potential antiviral evaluation of violacein produced by Chromobacterium violaceum. Mem Inst Oswaldo Cruz 98, 843–848 (2003). [DOI] [PubMed] [Google Scholar]
- Cazoto L. L., Martins D., Ribeiro M. G., Duran N. & Nakazato G. Antibacterial activity of violacein against Staphylococcus aureus isolated from bovine mastitis. J Antibiot 64, 395–397, 10.1038/ja.2011.13 (2011). [DOI] [PubMed] [Google Scholar]
- Subramaniam S., Ravi V. & Sivasubramanian A. Synergistic antimicrobial profiling of violacein with commercial antibiotics against pathogenic micro-organisms. Pharm Biol 52, 86–90, 10.3109/13880209.2013.815634 (2014). [DOI] [PubMed] [Google Scholar]
- Nakamura Y., Sawada T., Morita Y. & Tamiya E. Isolation of a psychrotrophic bacterium from the organic residue of a water tank keeping rainbow trout and antibacterial effect of violet pigment produced from the strain. Biochem Eng J 12, 79–86 (2002). [Google Scholar]
- Sanji N. et al. Antibiotic Sensitivity of the E. coli, Staph. aureus, Klebsiella, Acinetobacter, Pseudomonas and Enterococci (Eskape) Organisms Isolated From the Endotracheal Tubes From Intensive Care Units of a Tertiary Care Hospital. Indian J Pharmacol 45, S194–S195 (2013). [Google Scholar]
- Boucher H. W. et al. Bad Bugs, No Drugs: No ESKAPE! An Update from the Infectious Diseases Society of America. Clin Infect Dis 48, 1–12, 10.1086/595011 (2009). [DOI] [PubMed] [Google Scholar]
- Jiang P. X., Wang H. S., Zhang C., Lou K. & Xing X. H. Reconstruction of the violacein biosynthetic pathway from Duganella sp B2 in different heterologous hosts. Appl Microbiol Biotech 86, 1077–1088, 10.1007/s00253-009-2375-z (2010). [DOI] [PubMed] [Google Scholar]
- Jiang P. X. et al. Pathway redesign for deoxyviolacein biosynthesis in Citrobacter freundii and characterization of this pigment. Appl Microbiol Biotech 94, 1521–1532, 10.1007/s00253-012-3960-0 (2012). [DOI] [PubMed] [Google Scholar]
- Rodrigues A. L. et al. Microbial production of the drugs violacein and deoxyviolacein: analytical development and strain comparison. Biotechnol Lett 34, 717–720, 10.1007/s10529-011-0827-x (2012). [DOI] [PubMed] [Google Scholar]
- Rodrigues A. L. et al. Systems metabolic engineering of Escherichia coli for production of the antitumor drugs violacein and deoxyviolacein. Metab Eng 20, 29–41, 10.1016/j.ymben.2013.08.004 (2013). [DOI] [PubMed] [Google Scholar]
- Moss M. O. & Ryall C. Distribution of Chromobacteria in a Lowland River. Microbial Ecol 7, 139–149, 10.1007/Bf02032496 (1981). [DOI] [PubMed] [Google Scholar]
- Hoshino T. Violacein and related tryptophan metabolites produced by Chromobacterium violaceum: biosynthetic mechanism and pathway for construction of violacein core. Appl Microbiol Biotech 91, 1463–1475, 10.1007/s00253-011-3468-z (2011). [DOI] [PubMed] [Google Scholar]
- Mccarthy S. A., Johnson R. M., Kakimoto D. & Sakata T. Effects of Various Agents on the Pigment (Violacein) and Antibiotic Production of Alteromonas-Luteoviolacea. B Jpn Soc Sci Fish 51, 1115–1121 (1985). [Google Scholar]
- Zhang X. & Enomoto K. Characterization of a gene cluster and its putative promoter region for violacein biosynthesis in Pseudoalteromonas sp 520P1. Appl Microbiol Biotech 90, 1963–1971, 10.1007/s00253-011-3203-9 (2011). [DOI] [PubMed] [Google Scholar]
- Lu Y. et al. Production of violet pigment by a newly isolated psychrotrophic bacterium from a glacier in Xinjiang, China. Biochem Eng J 43, 135–141, 10.1016/j.bej.2008.09.009 (2009). [DOI] [Google Scholar]
- Pantanella F. et al. Violacein and biofilm production in Janthinobacterium lividum. J Appl Microbiol 102, 992–999, 10.1111/j.1365-2672.2006.03155.x (2007). [DOI] [PubMed] [Google Scholar]
- Wang H. S. et al. Optimization of culture conditions for violacein production by a new strain of Duganella sp B2. Biochem Eng J 44, 119–124, 10.1016/j.bej.2008.11.008 (2009). [DOI] [Google Scholar]
- Avgustin J. A., Bertok D. Z., Kostanjsek R. & Avgustin G. Isolation and characterization of a novel violacein-like pigment producing psychrotrophic bacterial species Janthinobacterium svalbardensis sp nov. Anton Leeuw Int J G 103, 763–769, 10.1007/s10482-012-9858-0 (2013). [DOI] [PubMed] [Google Scholar]
- Kim S. J. et al. Genome Sequence of Janthinobacterium sp Strain PAMC 25724, Isolated from Alpine Glacier Cryoconite. J Bacteriol 194, 2096–2096, 10.1128/Jb.00096-12 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moss M. O., Ryall C. & Logan N. A. Classification and Characterization of Chromobacteria from a Lowland River. J Gen Microbiol 105, 11–21 (1978). [Google Scholar]
- Li W. J. et al. Duganella violaceinigra sp. nov., a novel mesophilic bacterium isolated from forest soil. Int J Sys Evol Microbiol 54, 1811–1814, 10.1099/ijs.0.63141-0 (2004). [DOI] [PubMed] [Google Scholar]
- Aranda S., Montes-Borrego M. & Landa B. B. Purple-Pigmented Violacein-Producing Duganella spp. Inhabit the Rhizosphere of Wild and Cultivated Olives in Southern Spain. Microbial Ecol 62, 446–459, 10.1007/s00248-011-9840-9 (2011). [DOI] [PubMed] [Google Scholar]
- Hakvag S. et al. Violacein-producing Collimonas sp. from the sea surface microlayer of costal waters in Trondelag, Norway. Mar Drugs 7, 576–588, 10.3390/md7040576 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kampfer P., Wellner S., Lohse K., Martin K. & Lodders N. Duganella phyllosphaerae sp. nov., isolated from the leaf surface of Trifolium repens and proposal to reclassify Duganella violaceinigra into a novel genus as Pseudoduganella violceinigra gen. nov., comb. nov. (vol 35, pg 19, 2012). Syst Appl Microbiol 35, 278–278, 10.1016/j.syapm.2012.02.001 (2012). [DOI] [PubMed] [Google Scholar]
- Hahn M. W. & Hofle M. G. Flagellate predation on a bacterial model community: interplay of size-selective grazing, specific bacterial cell size, and bacterial community composition. Appl Environ Microbiol 65, 4863–4872 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vandamme D., Foubert I. & Muylaert K. Flocculation as a low-cost method for harvesting microalgae for bulk biomass production. Trends Biotechnol 31, 233–239, 10.1016/j.tibtech.2012.12.005 (2013). [DOI] [PubMed] [Google Scholar]
- Witte W. Antibiotic resistance in gram-positive bacteria: epidemiological aspects. J Antimicrob Chemother 44 Suppl A, 1–9 (1999). [DOI] [PubMed] [Google Scholar]
- Archer G. L. Staphylococcus aureus: a well-armed pathogen. Clin Infect Dis 26, 1179–1181 (1998). [DOI] [PubMed] [Google Scholar]
- Chambers H. F. & Deleo F. R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat Rev Microbiol 7, 629–641, 10.1038/nrmicro2200 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niazi J. H., Kim B. C., Ahn J. M. & Gu M. B. A novel bioluminescent bacterial biosensor using the highly specific oxidative stress-inducible pgi gene. Biosens Bioelectron 24, 670–675, 10.1016/j.bios.2008.06.026 (2008). [DOI] [PubMed] [Google Scholar]
- Wang H. S. et al. Biosynthesis and characterization of violacein, deoxyviolaceiri and oxyviolacein in heterologous host, and their antimicrobial activities. Biochem Eng J 67, 148–155, 10.1016/j.bej.2012.06.005 (2012). [DOI] [Google Scholar]
- Ahmad W. A., Yusof N. Z., Nordin N., Zakaria Z. A. & Rezali M. F. Production and characterization of violacein by locally isolated Chromobacterium violaceum grown in agricultural wastes. Appl Biochem Biotech 167, 1220–1234, 10.1007/s12010-012-9553-7 (2012). [DOI] [PubMed] [Google Scholar]
- Mavrodi D. V. et al. Diversity and evolution of the phenazine biosynthesis pathway. Appl Environ Microbiol 76, 866–879, 10.1128/AEM.02009-09 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamura K., Stecher G., Peterson D., Filipski A. & Kumar S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol 30, 2725–2729, 10.1093/molbev/mst197 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adler C. et al. Catecholate Siderophores Protect Bacteria from Pyochelin Toxicity. PLoS One 7, e46754, 10.1371/journal.pone.0046754 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarker S. D., Nahar L. & Kumarasamy Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 42, 321–324, 10.1016/j.ymeth.2007.01.006 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]