

Abstract
The function of retinal ganglion cells (RGCs) can be non-invasively assessed in experimental and genetic models of glaucoma by means of variants of the ERG technique that emphasize the activity of inner retina neurons. The best understood technique is the Pattern Electroretinogram (PERG) in response to contrast-reversing gratings or checkerboards, which selectively depends on the presence of functional RGCs. In glaucoma models, the PERG can be altered before histological loss of RGCs; PERG alterations may be either reversed with moderate IOP lowering or exacerbated with moderate IOP elevation. Under particular luminance-stimulus conditions, the Flash-ERG displays components that may reflect electrical activity originating in the proximal retina and be altered in some experimental glaucoma models (positive Scotopic Threshold response, pSTR; negative Scotopic Threshold Response, nSTR; Photopic Negative Response, PhNR; Oscillatory Potentials, OPs; multifocal ERG, mfERG). It is not yet known which of these components is most sensitive to glaucomatous damage. Electrophysiological assessment of RGC function appears to be a necessary outcome measure in experimental glaucoma models, which complements structural assessment and may even predict it. Neuroprotective strategies could be tested based on enhancement of baseline electrophysiological function that results in improved RGC survival. The use of electrophysiology in glaucoma models may be facilitated by specifically designed instruments that allow high throughput, robust assessment of electrophysiological function.
Keywords: glaucoma, retinal ganglion cells, pattern electroretinogram, electroretinogram, intraocular pressure, animal models
Introduction
For a very long time, a well-established notion has been that the ERG did not reflect electrical activity of retinal ganglion cells (RGCs)(Riggs, 1986). In 1981, however, a study in the cat (Maffei and Fiorentini, 1981) showed that the ERG in response to contrast-reversing patterns at constant mean luminance (Pattern-ERG, PERG) was abolished after section of the optic nerve that retrogradely killed RGCs leaving outer retina neurons and the standard Flash-ERG intact. This crucial experiment has been repeated many times in different mammals including nonhuman primates and rodents (rats and mice), with similar results. These studies sparked a considerable wave of interest resulting in a large number of clinical and experimental reports that the PERG is altered in glaucoma, even at early stages.
More recently, different laboratories discovered that under particular luminance-stimulus conditions, the Flash-ERG displays components (positive Scotopic Threshold response, pSTR; negative Scotopic Threshold Response, nSTR; Photopic Negative Response, PhNR; Oscillatory Potentials, OPs; multifocal ERG, mfERG) that may reflect electrical activity originating in the proximal retina. These inner-retina-sensitive ERG components have also been shown to be altered in clinical and experimental studies of glaucoma. Table I summarizes studies that used PERG and other inner-retina-sensitive ERG components in experimental models of glaucoma.
Table 1. Electrophysiological measurements used in glaucoma models.
Pattern Electroretinogram, PERG; positive Scotopic Threshold response, pSTR; negative Scotopic Threshold Response, nSTR; Photopic Negative Response, PhNR; Oscillatory Potentials, OPs; multifocal ERG, mfERG; Trabecular Meshwork, TM; Morrison, hypertonic saline injection of episcleral veins.(Morrison et al., 1997)
| Species | Model | Altered electrophysiological measure | Reference |
|---|---|---|---|
| Monkey | TM laser | PhNR in mfERG | (Luo et al., 2011) |
| Monkey | TM laser | OPs of mfERG | (Rangaswamy et al., 2006) |
| Monkey | TM laser | mfERG | (Fortune et al., 2012) |
| Monkey | TM laser | PhNR | (Viswanathan et al., 1999) |
| Monkey | TM laser | PERG | (Johnson et al., 1989) |
| Monkey | TM laser | PERG | (Marx et al., 1986) |
| Monkey | TM laser | PERG | (Marx et al., 1988) |
| Monkey | TM laser | nSTR | (Frishman et al., 1996) |
| Monkey | TM laser | mfERG Ops, STR | (Rangaswamy et al., 2006) |
| Monkey | TM laser | nSTR | (Frishman et al., 1996) |
| Monkey | TM laser | PhNR, PERG | (Viswanathan et al., 2000) |
| Monkey | TM laser | mfERG | (Frishman et al., 2000) |
| Monkey | TM laser | mfERG | (Raz et al., 2002) |
| Monkey | TM laser | mfERG | (He et al., 2014) |
| Monkey | TM laser | mfERG | (Nork et al., 2010) |
| Monkey | TM laser | mfERG | (Raz et al., 2003) |
| Monkey | TM laser | nfERG | (Nork et al., 2014) |
| Mouse | DBA/2J | PERG | (Saleh et al., 2007) |
| Mouse | DBA/2J | PERG | (Porciatti et al., 2007) |
| Mouse | DBA/2J | PERG | (Nagaraju et al., 2007) |
| Mouse | DBA/2J | PERG | (Porciatti and Nagaraju, 2010) |
| Mouse | DBA/2J | PERG | (Howell et al., 2007) |
| Mouse | DBA/2NNia | PERG | (Bayer et al., 2001a) |
| Mouse | DBA/2J | PERG | (Howell et al., 2012) |
| Mouse | DBA/2J | ERG | (Heiduschka et al., 2010) |
| Mouse | DBA/2J | ERG | (Harazny et al., 2009) |
| Mouse | DBA/2J | pSTR | (Perez de Lara et al., 2014) |
| Mouse | MYOC | PERG | (Chou et al., 2014b) |
| Mouse | MYOC | PERG | (Zode et al., 2012; Zode et al., 2011) |
| Mouse | Dexamethasone | PERG | (Zode et al., 2014) |
| Mouse | C57BL/6J, Indocyanine green + laser anterior chamber | ERG | (Grozdanic et al., 2003a) |
| Rat | Morrison | pSTR | (Fortune et al., 2004) |
| Rat | Episcleral vein cauthery | ERG, OPs | (Bayer et al., 2001b) |
| Rat | Episcleral vein cauthery | PERG, ERG | (Sandalon et al., 2013) |
| Rat | Morrison | ERG | (Georgiou et al., 2014) |
| Rat | Morrison | PERG | (Husain et al., 2012) |
| Rat | Morrison | PERG | (Abdul et al., 2013) |
| Rat | Morrison | pSTR | (Fortune et al., 2004) |
| Rat | Vortex and episcleral vein cauthery | ERG | (Grozdanic et al., 2003b) |
| Dog | Hereditary angle-closure glaucoma | PERG | (Grozdanic et al., 2010) |
Response generators
Generators of PERG and inner-retina-sensitive ERG components have been investigated in a substantial number of studies. In summary, 1) Optic nerve transection/crush resulting in selective RGC loss invariably cause dramatic loss of PERG signal in cats (Maffei and Fiorentini, 1981; Weber et al., 2008) monkeys (Gianfranceschi et al., 1999) rats(Berardi et al., 1990) and mice (Miura et al., 2009; Porciatti et al., 1996; Xia et al., 2014); pSTR and nSTR appear to be reduced in rats (Bui and Fortune, 2004) and mice (Liu et al., 2014; Smith et al., 2014; Yukita et al., 2015), but relatively less than the PERG (Liu et al., 2014); PhNR, OPs do not seem to be reduced in rats and mice (Li et al., 2005; Liu et al., 2014; Smith et al., 2014); in monkeys, retrograde RGC degeneration modestly alters the mfERG, and mfERG alterations are species-dependent (Nork et al., 2010), 2) Several pharmacological studies have demonstrated that interfering in various ways with activity of inner retina neurons reduces PERG as well as innerretina-sensitive ERG components in cats, rodents and primates(Bui and Fortune, 2004; Hare and Ton, 2002; Hood et al., 1999; Viswanathan et al., 2000). Interestingly, both spiking and non-spiking electrical activity contributes to the PERG and inner-retina-sensitive ERG components (Luo and Frishman, 2011; Miura et al., 2009; Trimarchi et al., 1990) (Harrison et al., 2006; Viswanathan et al., 2000), 3) Intraretinal recordings have demonstrated an inner retina origin for the PERG distinct from ERG b-wave (Baker et al., 1988; Sieving and Steinberg, 1987), 4) In the mouse, the bioelectrical field generated by the PERG is different from that of the Flash-ERG, and it is consistent with generators localized in the optic nerve head (Chou and Porciatti, 2012), 5) Functional retrograde transport of target-derived factors is necessary for PERG generation, as blocking axon transport in the optic nerve Chou (Chou et al., 2013) or lesioning the superior colliculus (Yang et al., 2013) impairs the PERG in the mouse.
Despite extensive investigation, the precise cellular origin of the PERG and inner-retinasensitive ERG components is not yet known. While the PERG appears to be more specifically related to the presence of functional RGCs, different inner-retina-sensitive ERG components appear to arise from different classes of neurons and/or non-neuronal glial cells in the inner retina and be species-specific. IOP elevation often alters one or more of these components, and this may also occur when RGCs are retrogradely degenerated, suggesting a preganglionic component (Nork et al., 2010). Further investigation is needed to better understand species differences and which component is most sensitive to RGC damage (Liu et al., 2014) (Smith et al., 2014).
The standard scotopic and photopic ERG is generally considered insensitive to glaucoma, and is often used as a control that the procedure to elevate IOP in experimental models has not induced a generalized retinal damage (Bui et al., 2013). Some studies, however, have shown that the ERG can be altered in experimental models as well as in genetic models (Table 1).
Visually Evoked Potentials (VEP) have been sometimes used to assess whether RGC axons are capable to carrying retinal signals along the geniculo-cortical pathway (Chou et al., 2014b; Heiduschka et al., 2010). It should be considered that the numerical relationship between topographically matched RGCs and contralateral geniculate relay cells is approximately 3:1 in mice (Coleman et al., 2009). This retina-geniculate convergence, together with the compensatory mechanisms occurring in the visual cortex in response to progressive reduction of the retinal output in disease (Keck et al., 2008) may offset peripheral losses and complicate the interpretation of VEP alterations.
The PERG
At present, the PERG is the most specific technique for electrophysiological assessment of RGC function in primate and rodent models of glaucoma. In the pattern-reversal stimulus, pattern elements alternate at constant mean luminance. Inner retina neurons with receptive field organized in antagonistic regions respond at each contrast transition, whereas photoreceptor activity generated by adjacent pattern elements is in opposition of phase and cancels out at the electrode (Porciatti, 2007). Thus, the PERG is dominated by inner retina activity whereas outer retina activity is minimized. The pattern stimulus, compared to luminance stimuli, has the unique advantage that its characteristics (spatial frequency, contrast, temporal frequency, chromaticity) can be independently modulated at constant mean luminance. Hence the activity of RGC subpopulations can be maximized by using appropriate pattern stimuli at which RGCs best respond (Hess et al., 1986; Morrone et al., 1994). In addition, important measures of visual performance such as spatial acuity and contrast threshold can be obtained by determining the smallest pattern size and contrast, respectively, at which a PERG signal larger than noise is recordable (Figure 1) (Porciatti, 2007).
Figure 1.
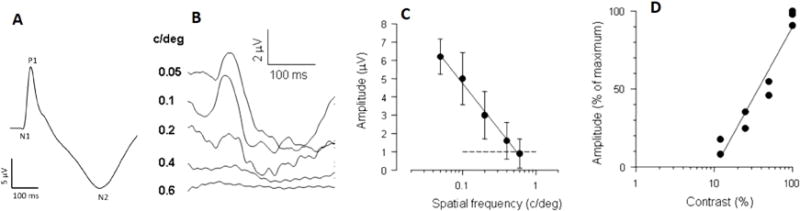
Transient PERGs recorded in C57BL/6J mice in response to high-contrast gratings reversing 2 times/s. A: grand-average of 120 waveforms from different eyes.(Chou et al., 2014a) Note the N1-P1-N2 complex. B: The PERG amplitude decreases with increasing spatial frequency.(Porciatti, 2007) C: The mean (±SEM) PERG amplitude of 6 different mice decreases with increasing spatial frequency and reaches the noise level (dashed line) at 0.6 cy/deg (retinal acuity)(Porciatti, 2007), which is similar to the behavioral acuity.(Gianfranceschi et al., 1999) C: For gratings of 0.05 cy/deg, the PERG amplitude decreases with decreasing contrast. The contrast threshold is about 10%, (Porciatti et al., 1996) which is similar to that measured with the optomotor system. (Prusky et al., 2004)
In experimental models, the PERG is typically recorded from the cornea using a variety of electrodes that do not interfere with the eye optics (Porciatti et al., 2007; Viswanathan et al., 2001). In primates and cats, optical correction for the viewing distance is needed to keep the stimulus focused in the retina. In mice, pupil dilation is not necessary as the small pupil size maximizes the depth-of-field (Remtulla and Hallett, 1985; Schmucker and Schaeffel, 2004). Recently, the PERG technique in mouse models has been greatly simplified by the introduction of a new recording paradigm that simultaneously derives responses from both eyes using one subcutaneous needle electrode in the snout and one channel recording (Chou et al., 2014a). The paradigm is now incorporated in a commercially available instrument (Miami-PERG, Jörvec Corp., Miami, FL).
Examples of mouse PERG waveforms and how PERG amplitude changes as function of spatial frequency and contrast are shown in Figure 1. In C57B/6J mice, the PERG waveform (Fig. 1A) consists of a small negative wave peaking at about 50 ms (N1), a positive wave peaking at about 80 ms (P1) and a prominent, broad negativity peaking at about 300 ms (N2). In different mouse strains such DBA/2J the P1 peak latency can be longer by 20 ms compared to C57BL/6J. (Porciatti et al., 2010) The general morphology of the mouse PERG waveform qualitatively resembles that of human and non-human primate PERG that consists of an early negativity (N35) followed by a larger positivity (P50) and a later negativity (N95)(Viswanathan et al., 2000) (Bach et al., 2013). However, it is not yet known whether the mouse N1-P1-N2 complex and the primate N35-P50-N95 complex reflect same neural substrates. One striking difference is that the mouse PERG latency dramatically increases (up to 60 ms) by reducing the stimulus contrast or increasing the spatial frequency (Porciatti et al., 2010) whereas in the primate contrast-dependent latency changes PERG are relatively minor. This has been interpreted as longer integration time over the large receptive field of mouse RGCs. Also, there are significant differences in the PERG amplitude/latency functions as a function of contrast in different mouse genotypes. This may reflect genotype-dependent contrast gain relationship in the circuitry involving RGC receptive field (Porciatti et al., 2010).
Contribution of electrophysiology to better understanding of structure-function relationships in glaucoma models
RGC dysfunction precedes RGC death
Several laboratories using different experimental models of ocular hypertension and different electrophysiological measurements of RGC function have shown that RGC dysfunction occurs early, is progressive, and precedes loss of optic nerve tissue and RGC density. Marx et al, (Marx et al., 1986; Marx et al., 1988) induced sustained IOP elevation in monkeys by means of laser photocoagulation of the trabecular meshwork. While the flash ERG was normal, the PERG was much reduced despite the absence of cupping of the optic nerve head, which occurred subsequently. In a similar monkey model, Luo et al (Luo et al., 2014) recently recorded multifocal ERGs and analyzed several components of the mfERG including the PhNR. All mfERG measures captured functional losses that were more spatially extensive and profound than the structural losses as measured by spectral domain OCT, consistent with neuronal (and/or glial) dysfunction before structural loss. In rats made hypertensive with hypertonic saline injection into an episcleral vein, Fortune et al, (Fortune et al., 2004) demonstrated loss of pSTR before the onset of loss of optic nerve tissue assessed histologically. Comparable electrophysiological changes are reported to occur in genetic mouse glaucoma models. Howell et al, (Howell et al., 2007) showed in DBA/2J mice that the PERG is impaired early in the disease, as IOP becomes progressively elevated but before detectable axon loss. Saleh et al, (Saleh et al., 2007) showed in DBA/2J mice that PERG losses are progressive and precede loss of RNFL thickness (Figure 2). In some genetic models, progressive RGC dysfunction may develop without IOP elevation. Chou et al, (Chou et al., 2014b) showed in transgenic mice expressing Tyr437His mutant of human myocilin protein that PERG amplitude progressively decreases while PERG latency increases between 2 and 18 months at constant normal IOP levels. This model is associated with moderate RGC loss but axonal swelling and astrocytic activation in the optic nerve head. In a related transgenic mouse carrying the Y437H MYOC mutation, Zode et al. (Zode et al., 2011) reported sustained IOP elevation resulting in progressive, parallel loss of PERG signal and RGC density. Altogether, these results demonstrate that RGC dysfunction precedes RGC death and may depend on the level of IOP elevation. The time lag between RGC dysfunction and death may depend both on the magnitude of IOP elevation and the genetic susceptibility to IOP stress (Libby et al., 2005b). Axonal and dendritic dysfunction preceding loss of RGC somas has been also demonstrated with structural studies (Buckingham et al., 2008; Williams et al., 2013).
Figure 2.

Progressive loss of PERG amplitude in DBA/2J mice. A: serial PERG recordings in a population of DBA/2J mice (individual mice, grey lines). The PERG amplitude reaches the noise level (dashed line) at different ages between 8 and 12 months. Mice with PERG amplitude at noise level were eliminated from the pool for histological analysis. The open symbols connected by a thick line represent the group mean ± SEM amplitude. C: corresponding IOP measurements showing a progressive IOP increase with age. D: The PERG amplitude is inversely correlated to IOP. B: Relative changes of PERG amplitude (open circles), axon counts (open squares) and histological RNFL thickness (filled circles) as a function of age. Error bars represent the SD. Progressive loss of RGC axons and RNFL thickness lag behind progressive loss of PERG amplitude. Redrawn from ref. (Saleh et al., 2007). Axon counts estimates are from refs. (Anderson et al., 2005; Libby et al., 2005a)
RGC dysfunction may be reversible with IOP lowering
If RGC dysfunction precedes death and depends on IOP elevation, then it may be reversible upon IOP lowering. This question has been addressed in a number of studies. In DBA/2J mice aged 11 months, the PERG signal is close to the noise level, but RGC death is still moderate (Howell et al., 2007; Libby et al., 2005a; Saleh et al., 2007; Williams et al., 2013). In these mice, Nagaraju et al, (Nagaraju et al., 2007) administered intraperitoneally mannitol 25% (2.5 g/Kg), which reduced IOP by 38%; the PERG amplitude increased by 83%. In mice, IOP is posture-dependent. IOP increases upon head-down (Aihara et al., 2003; Nagaraju et al., 2007) and decreases upon head-up (Nagaraju and Porciatti, 2008). For a 60 deg change from horizontal–either head-down or head-up – the posture-dependent IOP change is of the order of ±5 mm Hg and it is stable over time, which allows testing the effect of posture on RGC function. Nagaraju et al. (Nagaraju and Porciatti, 2008) showed that head-up posture reduced IOP by about 5 mm Hg in DBA/2J mice of different ages. The normal PERG signal of young DBA/2J mice was unaffected, but the abnormal PERG signal of older DBA/2J mice improved in an age-dependent manner: the worse the baseline PERG amplitude, the larger the magnitude of PERG improvement (Figure 3).
Figure 3.

Head-up posture reduces IOP and improves abnormal PERG in susceptible ages in DBA/2J mice. A: IOP decreases during 60° head-up posture (HU) by about 5 mm Hg compared to horizontal baseline (B) and recovery (R) in mice of different ages with different baseline IOP. B: PERG amplitude increases during HU in mice 6 and 10 months old, when baseline PERG is reduced. C: Improvement of PERG amplitude is also obtained in mice 11 months old with abnormal baseline PERG after mannitol 20% treatment, which reduced IOP by about 38%. Redrawn from ref.(Porciatti and Nagaraju, 2010)
RGC dysfunction may be inducible with moderate IOP elevation in susceptible eyes
If RGC dysfunction precedes death and depends on IOP elevation, then it may be inducible upon transient IOP elevation (Bui et al., 2005) in susceptible eyes. As mentioned above, IOP increases during head-down posture. As IOP elevation is a gravity-induced response to elevation of the episcleral venous pressure, it is rather independent of mouse strain, age, and baseline IOP. Nagaraju et al, (Nagaraju et al., 2007) showed that 60° head-down posture resulting in IOP elevation of about 5 mm Hg did not have any effect on the normal PERG signal of DBA/2J mice 2 months old. However, it reduced the PERG signal of DBA/2J mice 6 months old and even more the PERG of mice 10 months old (Figure 4). Some mouse strains, such as C57BL/6J appear to be less susceptible to IOP elevation than DBA/2J. Anderson et al, (Anderson et al., 2006) generated C57BL/6J-Tyrp1b.GpnmbR150X mice that develop the same iris disease as DBA/2J mice, but are resistant to high IOP and glaucoma. In these mice the PERG is normal (Howell et al., 2007). Experimental IOP elevation causes different mfERG alterations in different monkey species (Nork et al., 2010).
Figure 4.
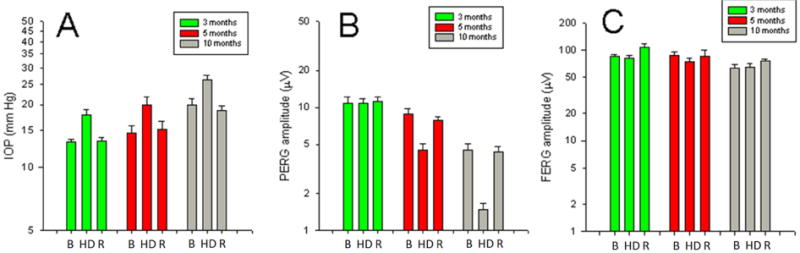
Head-down posture increases IOP and reduces PERG in susceptible ages in DBA/2J mice. A: IOP increases during 60° head-down posture (HD) by about 5 mm Hg compared to horizontal baseline (B) and recovery (R) in mice of different ages with different baseline IOP. B: PERG amplitude decreases during HD in an age-dependent manner. C: The light-adapted, peak-to-trough flash ERG amplitude does not change during HD. Vertical Bars represent the mean, whiskers represent the SEM. Redrawn from ref. (Nagaraju et al., 2007)
Neuroprotection and RGC function
A number of neuroprotective agents are known to spare at least part of RGCs from death in glaucoma models (Danesh-Meyer, 2011). Less is known about protection of RGC function. Surviving RGCs may not be functional. Also, it is not known whether neuroprotective agents alter RGC function in control eyes (McGill et al., 2007). So far, a few studies have linked RGC electrical activity to the number of surviving RGCs in genetic mouse glaucoma models. Zode et al, (Zode et al., 2012; Zode et al., 2014) showed that topical administration of ocular sodium 4-phenylbutyrate rescues glaucoma and preserves the PERG in transgenic MYOC(Y437H) mice with elevated IOP as well as in mice with dexamethasone-induced IOP elevation. Howell et al, (Howell et al., 2012) showed in DBA/2J mice that radiation treatment prevents neuronal damage and spares the PERG. Howell et al, (Howell et al., 2007) showed in DBA/2J mice that the Wlds allele, which is known to protect against insults to axons, strongly protects against DBA/2J glaucoma and preserves the PERG.
If RGC dysfunction precedes death, then the magnitude of electrophysiological change (worsening/improvement) upon transient IOP elevation/lowering may help to assess susceptibility/reversibility of RGC to IOP stress and predict the outcome of the disease without or with treatment. This hypothesis can be satisfactorily modeled (Porciatti and Ventura, 2012) and is supported by the studies on PERG modifiability to IOP challenge quoted above and others studies showing that IOP lowering rescues RGCs from death in glaucoma models (Schuettauf et al., 2002; Zode et al., 2011; Zode et al., 2014). A similar rationale could be used for neuroprotective strategies other than IOP lowering. This seems an important field of investigation, as it would allow testing neuroprotective strategies and formulating predictions based on baseline assessment of electrophysiological modifiability instead of waiting several months to assess the histological outcome.
Strengths and weaknesses of electrophysiological testing in glaucoma
As glaucoma impacts primarily RGCs but also inner and perhaps outer retina neurons, any technique that probes electrical activity of inner retina is an excellent candidate to monitor glaucomatous functional changes. Thus, both the PERG and a number of flash-ERG derived techniques may be effectively used to this purpose, and the choice of one method over the other is a matter of pros/cons considerations on particular experimental models and procedures at the level of individual laboratories (Table 1). While the PERG appears to be the response most specific and sensitive for RGC dysfunction, and is also recordable in minutes under light-adapted conditions, it depends on intact eye optics. Large eyes of cats and monkeys need to be refracted for the viewing distance, and the presence of optical opacities precludes PERG recording. As mice are prone to developing reversible cataracts under stress, care is needed with the level of anesthesia and eye manipulation to prevent the formation of cataracts. Similar considerations apply to the mfERG. The presence of cataract is less of a problem with flash-ERG derived techniques. On the other hand, recording threshold responses such as pSTR requires long dark adaptation periods and a series of responses to different flash intensities to identify the response threshold.
It should be also considered that both the PERG and inner-retina sensitive ERG components may be secondarily altered for pathological conditions affecting the outer retina. In experimental models of glaucoma, when induced IOP elevation is high enough to cause outer retina injury, it is advisable to record a standard ERG to control for generalized effects.
Concluding remarks
Electrophysiological assessment of RGC function appears to be a sensitive and necessary outcome measure in experimental glaucoma models, which complements structural assessment and may even predict it. In addition to the PERG, several components of the luminance ERG appear to be inner-retina sensitive and have a promising role. The use of electrophysiology in mouse models may be facilitated by new, user-friendly protocols and instruments that allow high throughput, robust assessment of RGC function.
Retinal ganglion cell function is measurable with electrophysiology in in-vivo glaucoma models.
PERG is the most specific and sensitive method.
Retinal ganglion cell function is a necessary outcome measure
Acknowledgments
Financial support: NIH-NEI RO1EY019077, R43EY023460, center grant P30-EY014801, unrestricted grant to Bascom Palmer Eye Institute from Research to Prevent Blindness, Inc.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Financial disclosure: none
References
- Abdul Y, Akhter N, Husain S. Delta-opioid agonist SNC-121 protects retinal ganglion cell function in a chronic ocular hypertensive rat model. Invest Ophthalmol Vis Sci. 2013;54:1816–1828. doi: 10.1167/iovs.12-10741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aihara M, Lindsey JD, Weinreb RN. Episcleral venous pressure of mouse eye and effect of body position. Curr Eye Res. 2003;27:355–362. doi: 10.1076/ceyr.27.6.355.18194. [DOI] [PubMed] [Google Scholar]
- Anderson MG, Libby RT, Gould DB, Smith RS, John SW. High-dose radiation with bone marrow transfer prevents neurodegeneration in an inherited glaucoma. Proc Natl Acad Sci U S A. 2005;102:4566–4571. doi: 10.1073/pnas.0407357102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson MG, Libby RT, Mao M, Cosma IM, Wilson LA, Smith RS, John SW. Genetic context determines susceptibility to intraocular pressure elevation in a mouse pigmentary glaucoma. BMC Biol. 2006;4:20. doi: 10.1186/1741-7007-4-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bach M, Brigell MG, Hawlina M, Holder GE, Johnson MA, McCulloch DL, Meigen T, Viswanathan S. ISCEV standard for clinical pattern electroretinography (PERG): 2012 update. Doc Ophthalmol. 2013;126:1–7. doi: 10.1007/s10633-012-9353-y. [DOI] [PubMed] [Google Scholar]
- Baker CL, Jr, Hess RR, Olsen BT, Zrenner E. Current source density analysis of linear and non-linear components of the primate electroretinogram. J Physiol. 1988;407:155–176. doi: 10.1113/jphysiol.1988.sp017408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayer AU, Brodie SE, Mittag T. Pilot study of pattern-electroretinographic changes in the DBA/2NNia mouse. Animal model of congenital angle-closure glaucoma. Ophthalmologe. 2001a;98:248–252. doi: 10.1007/s003470170157. [DOI] [PubMed] [Google Scholar]
- Bayer AU, Danias J, Brodie S, Maag KP, Chen B, Shen F, Podos SM, Mittag TW. Electroretinographic abnormalities in a rat glaucoma model with chronic elevated intraocular pressure. Exp Eye Res. 2001b;72:667–677. doi: 10.1006/exer.2001.1004. [DOI] [PubMed] [Google Scholar]
- Berardi N, Domenici L, Gravina A, Maffei L. Pattern ERG in rats following section of the optic nerve. Exp Brain Res. 1990;79:539–546. doi: 10.1007/BF00229323. [DOI] [PubMed] [Google Scholar]
- Buckingham BP, Inman DM, Lambert W, Oglesby E, Calkins DJ, Steele MR, Vetter ML, Marsh-Armstrong N, Horner PJ. Progressive ganglion cell degeneration precedes neuronal loss in a mouse model of glaucoma. J Neurosci. 2008;28:2735–2744. doi: 10.1523/JNEUROSCI.4443-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bui BV, Edmunds B, Cioffi GA, Fortune B. The gradient of retinal functional changes during acute intraocular pressure elevation. Invest Ophthalmol Vis Sci. 2005;46:202–213. doi: 10.1167/iovs.04-0421. [DOI] [PubMed] [Google Scholar]
- Bui BV, Fortune B. Ganglion cell contributions to the rat full-field electroretinogram. J Physiol. 2004;555:153–173. doi: 10.1113/jphysiol.2003.052738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bui BV, He Z, Vingrys AJ, Nguyen CT, Wong VH, Fortune B. Using the electroretinogram to understand how intraocular pressure elevation affects the rat retina. J Ophthalmol. 2013;2013:262467. doi: 10.1155/2013/262467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou TH, Park KK, Luo X, Porciatti V. Retrograde signaling in the optic nerve is necessary for electrical responsiveness of retinal ganglion cells. Investigative Ophthalmology and Visual Science. 2013;54:1236–1243. doi: 10.1167/iovs.12-11188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou TH, Bohorquez J, Toft-Nielsen J, Ozdamar O, Porciatti V. Robust mouse pattern electroretinograms derived simultaneously from each eye using a common snout electrode. Invest Ophthalmol Vis Sci. 2014a;55:2469–2475. doi: 10.1167/iovs.14-13943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou TH, Porciatti V. The bioelectric field of the pattern electroretinogram in the mouse. Invest Ophthalmol Vis Sci. 2012;53:8086–8092. doi: 10.1167/iovs.12-10720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou TH, Tomarev S, Porciatti V. Transgenic Mice Expressing Mutated Tyr437His Human Myocilin Develop Progressive Loss of Retinal Ganglion Cell Electrical Responsiveness and Axonopathy With Normal IOP. Invest Ophthalmol Vis Sci. 2014b;55:5602–5609. doi: 10.1167/iovs.14-14793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coleman JE, Law K, Bear MF. Anatomical origins of ocular dominance in mouse primary visual cortex. Neuroscience. 2009;161:561–571. doi: 10.1016/j.neuroscience.2009.03.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Danesh-Meyer HV. Neuroprotection in glaucoma: recent and future directions. Curr Opin Ophthalmol. 2011;22:78–86. doi: 10.1097/ICU.0b013e32834372ec. [DOI] [PubMed] [Google Scholar]
- Fortune B, Bui BV, Morrison JC, Johnson EC, Dong J, Cepurna WO, Jia L, Barber S, Cioffi GA. Selective ganglion cell functional loss in rats with experimental glaucoma. Invest Ophthalmol Vis Sci. 2004;45:1854–1862. doi: 10.1167/iovs.03-1411. [DOI] [PubMed] [Google Scholar]
- Fortune B, Burgoyne CF, Cull GA, Reynaud J, Wang L. Structural and functional abnormalities of retinal ganglion cells measured in vivo at the onset of optic nerve head surface change in experimental glaucoma. Invest Ophthalmol Vis Sci. 2012;53:3939–3950. doi: 10.1167/iovs.12-9979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frishman LJ, Saszik S, Harwerth RS, Viswanathan S, Li Y, Smith EL, 3rd, Robson JG, Barnes G. Effects of experimental glaucoma in macaques on the multifocal ERG. Multifocal ERG in laser-induced glaucoma. Doc Ophthalmol. 2000;100:231–251. doi: 10.1023/a:1002735804029. [DOI] [PubMed] [Google Scholar]
- Frishman LJ, Shen FF, Du L, Robson JG, Harwerth RS, Smith EL, 3rd, Carter-Dawson L, Crawford ML. The scotopic electroretinogram of macaque after retinal ganglion cell loss from experimental glaucoma. Invest Ophthalmol Vis Sci. 1996;37:125–141. [PubMed] [Google Scholar]
- Georgiou AL, Guo L, Francesca Cordeiro M, Salt TE. Electroretinogram and visual-evoked potential assessment of retinal and central visual function in a rat ocular hypertension model of glaucoma. Curr Eye Res. 2014;39:472–486. doi: 10.3109/02713683.2013.848902. [DOI] [PubMed] [Google Scholar]
- Gianfranceschi L, Fiorentini A, Maffei L. Behavioural visual acuity of wild type and bcl2 transgenic mouse. Vision Res. 1999;39:569–574. doi: 10.1016/s0042-6989(98)00169-2. [DOI] [PubMed] [Google Scholar]
- Grozdanic SD, Betts DM, Sakaguchi DS, Allbaugh RA, Kwon YH, Kardon RH. Laser-induced mouse model of chronic ocular hypertension. Invest Ophthalmol Vis Sci. 2003a;44:4337–4346. doi: 10.1167/iovs.03-0015. [DOI] [PubMed] [Google Scholar]
- Grozdanic SD, Betts DM, Sakaguchi DS, Kwon YH, Kardon RH, Sonea IM. Temporary elevation of the intraocular pressure by cauterization of vortex and episcleral veins in rats causes functional deficits in the retina and optic nerve. Exp Eye Res. 2003b;77:27–33. doi: 10.1016/s0014-4835(03)00089-7. [DOI] [PubMed] [Google Scholar]
- Grozdanic SD, Kecova H, Harper MM, Nilaweera W, Kuehn MH, Kardon RH. Functional and structural changes in a canine model of hereditary primary angle-closure glaucoma. Invest Ophthalmol Vis Sci. 2010;51:255–263. doi: 10.1167/iovs.09-4081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harazny J, Scholz M, Buder T, Lausen B, Kremers J. Electrophysiological deficits in the retina of the DBA/2J mouse. Doc Ophthalmol. 2009;119:181–197. doi: 10.1007/s10633-009-9194-5. [DOI] [PubMed] [Google Scholar]
- Hare WA, Ton H. Effects of APB, PDA, and TTX on ERG responses recorded using both multifocal and conventional methods in monkey. Effects of APB, PDA, and TTX on monkey ERG responses. Doc Ophthalmol. 2002;105:189–222. doi: 10.1023/a:1020553020264. [DOI] [PubMed] [Google Scholar]
- Harrison WW, Viswanathan S, Malinovsky VE. Multifocal pattern electroretinogram: cellular origins and clinical implications. Optom Vis Sci. 2006;83:473–485. doi: 10.1097/01.opx.0000218319.61580.a5. [DOI] [PubMed] [Google Scholar]
- He L, Yang H, Gardiner SK, Williams G, Hardin C, Strouthidis NG, Fortune B, Burgoyne CF. Longitudinal detection of optic nerve head changes by spectral domain optical coherence tomography in early experimental glaucoma. Invest Ophthalmol Vis Sci. 2014;55:574–586. doi: 10.1167/iovs.13-13245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heiduschka P, Julien S, Schuettauf F, Schnichels S. Loss of retinal function in aged DBA/2J mice – New insights into retinal neurodegeneration. Exp Eye Res. 2010;91:779–783. doi: 10.1016/j.exer.2010.09.001. [DOI] [PubMed] [Google Scholar]
- Hess RF, Baker CL, Zrenner E, Schwarzer J. Differences between electroretinograms of cat and primate. J Neurophysiol. 1986;56:747–768. doi: 10.1152/jn.1986.56.3.747. [DOI] [PubMed] [Google Scholar]
- Hood DC, Frishman LJ, Viswanathan S, Robson JG, Ahmed J. Evidence for a ganglion cell contribution to the primate electroretinogram (ERG): effects of TTX on the multifocal ERG in macaque. Vis Neurosci. 1999;16:411–416. doi: 10.1017/s0952523899163028. [DOI] [PubMed] [Google Scholar]
- Howell GR, Libby RT, Jakobs TC, Smith RS, Phalan FC, Barter JW, Barbay JM, Marchant JK, M N, Porciatti V, Whitmore AV, Masland RH, John SW. Axons of retinal ganglion cells are insulted in the optic nerve early in DBA/2J glaucoma. J Cell Biol. 2007;179:1523–1537. doi: 10.1083/jcb.200706181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howell GR, Soto I, Zhu X, Ryan M, Macalinao DG, Sousa GL, Caddle LB, MacNicoll KH, Barbay JM, Porciatti V, Anderson MG, Smith RS, Clark AF, Libby RT, John SW. Radiation treatment inhibits monocyte entry into the optic nerve head and prevents neuronal damage in a mouse model of glaucoma. J Clin Invest. 2012;122:1246–1261. doi: 10.1172/JCI61135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Husain S, Abdul Y, Crosson CE. Preservation of retina ganglion cell function by morphine in a chronic ocular-hypertensive rat model. Invest Ophthalmol Vis Sci. 2012;53:4289–4298. doi: 10.1167/iovs.12-9467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson MA, Drum BA, Quigley HA, Sanchez RM, Dunkelberger GR. Pattern-evoked potentials and optic nerve fiber loss in monocular laser-induced glaucoma. Invest Ophthalmol Vis Sci. 1989;30:897–907. [PubMed] [Google Scholar]
- Keck T, Mrsic-Flogel TD, Vaz Afonso M, Eysel UT, Bonhoeffer T, Hubener M. Massive restructuring of neuronal circuits during functional reorganization of adult visual cortex. Nat Neurosci. 2008;11:1162–1167. doi: 10.1038/nn.2181. [DOI] [PubMed] [Google Scholar]
- Li B, Barnes GE, Holt WF. The decline of the photopic negative response (PhNR) in the rat after optic nerve transection. Doc Ophthalmol. 2005;111:23–31. doi: 10.1007/s10633-005-2629-8. [DOI] [PubMed] [Google Scholar]
- Libby RT, Anderson MG, Pang IH, Robinson ZH, Savinova OV, Cosma IM, Snow A, Wilson LA, Smith RS, Clark AF, John SW. Inherited glaucoma in DBA/2J mice: pertinent disease features for studying the neurodegeneration. Vis Neurosci. 2005a;22:637–648. doi: 10.1017/S0952523805225130. [DOI] [PubMed] [Google Scholar]
- Libby RT, Gould DB, Anderson MG, John SW. Complex genetics of glaucoma susceptibility. Annu Rev Genomics Hum Genet. 2005b;6:15–44. doi: 10.1146/annurev.genom.6.080604.162209. [DOI] [PubMed] [Google Scholar]
- Liu Y, McDowell CM, Zhang Z, Tebow HE, Wordinger RJ, Clark AF. Monitoring Retinal Morphologic and Functional Changes in Mice Following Optic Nerve Crush. Investigative Ophthalmology & Visual Science. 2014;55:3766–3774. doi: 10.1167/iovs.14-13895. [DOI] [PubMed] [Google Scholar]
- Luo X, Frishman LJ. Retinal Pathway Origins of the Pattern Electroretinogram (PERG) Invest Ophthalmol Vis Sci. 2011;52:8571–8584. doi: 10.1167/iovs.11-8376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo X, Patel NB, Harwerth RS, Frishman LJ. Loss of the low-frequency component of the global-flash multifocal electroretinogram in primate eyes with experimental glaucoma. Invest Ophthalmol Vis Sci. 2011;52:3792–3804. doi: 10.1167/iovs.10-6667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo X, Patel NB, Rajagopalan LP, Harwerth RS, Frishman LJ. Relation between macular retinal ganglion cell/inner plexiform layer thickness and multifocal electroretinogram measures in experimental glaucoma. Invest Ophthalmol Vis Sci. 2014;55:4512–4524. doi: 10.1167/iovs.14-13937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maffei L, Fiorentini A. Electroretinographic responses to alternating gratings before and after section of the optic nerve. Science. 1981;211:953–955. doi: 10.1126/science.7466369. [DOI] [PubMed] [Google Scholar]
- Marx MS, Podos SM, Bodis-Wollner I, Howard-Williams JR, Siegel MJ, Teitelbaum CS, Maclin EL, Severin C. Flash and pattern electroretinograms in normal and laser-induced glaucomatous primate eyes. Invest Ophthalmol Vis Sci. 1986;27:378–386. [PubMed] [Google Scholar]
- Marx MS, Podos SM, Bodis-Wollner I, Lee PY, Wang RF, Severin C. Signs of early damage in glaucomatous monkey eyes: low spatial frequency losses in the pattern ERG and VEP. Exp Eye Res. 1988;46:173–184. doi: 10.1016/s0014-4835(88)80075-7. [DOI] [PubMed] [Google Scholar]
- McGill TJ, Prusky GT, Douglas RM, Yasumura D, Matthes MT, Nune G, Donohue-Rolfe K, Yang H, Niculescu D, Hauswirth WW, Girman SV, Lund RD, Duncan JL, LaVail MM. Intraocular CNTF Reduces Vision in Normal Rats in a Dose-Dependent Manner. Investigative Ophthalmology & Visual Science. 2007;48:5756–5766. doi: 10.1167/iovs.07-0054. [DOI] [PubMed] [Google Scholar]
- Miura G, Wang MH, Ivers KM, Frishman LJ. Retinal pathway origins of the pattern ERG of the mouse. Exp Eye Res. 2009;89:49–62. doi: 10.1016/j.exer.2009.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrison JC, Moore CG, Deppmeier LM, Gold BG, Meshul CK, Johnson EC. A rat model of chronic pressure-induced optic nerve damage. Exp Eye Res. 1997;64:85–96. doi: 10.1006/exer.1996.0184. [DOI] [PubMed] [Google Scholar]
- Morrone C, Fiorentini A, Bisti S, Porciatti V, Burr DC. Pattern-reversal electroretinogram in response to chromatic stimuli: II. Monkey. Vis Neurosci. 1994;11:873–884. doi: 10.1017/s0952523800003837. [DOI] [PubMed] [Google Scholar]
- Nagaraju M, Porciatti V. Head-Up Tilt Reduces IOP and Improves Abnormal PERG in DBA/2J Glaucoma. Invest Ophthalmol Vis Sci. 2008;49:1556. [Google Scholar]
- Nagaraju M, Saleh M, Porciatti V. IOP-Dependent Retinal Ganglion Cell Dysfunction in Glaucomatous DBA/2J Mice. Invest Ophthalmol Vis Sci. 2007;48:4573–4579. doi: 10.1167/iovs.07-0582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nork TM, Kim CB, Heatley GA, Kaufman PL, Lucarelli MJ, Levin LA, Ver Hoeve JN. Serial multifocal electroretinograms during long-term elevation and reduction of intraocular pressure in non-human primates. Doc Ophthalmol. 2010;120:273–289. doi: 10.1007/s10633-010-9231-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nork TM, Kim CBY, Munsey KM, Dashek RJ, Hoeve JNV. Regional Choroidal Blood Flow and Multifocal Electroretinography in Experimental Glaucoma in Rhesus Macaques. Investigative Ophthalmology & Visual Science. 2014;55:7786–7798. doi: 10.1167/iovs.14-14527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez de Lara MJ, Santano C, Guzman-Aranguez A, Valiente-Soriano FJ, Aviles-Trigueros M, Vidal-Sanz M, de la Villa P, Pintor J. Assessment of inner retina dysfunction and progressive ganglion cell loss in a mouse model of glaucoma. Exp Eye Res. 2014;122:40–49. doi: 10.1016/j.exer.2014.02.022. [DOI] [PubMed] [Google Scholar]
- Porciatti V. The mouse pattern electroretinogram. Doc Ophthalmol. 2007;115:145–153. doi: 10.1007/s10633-007-9059-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porciatti V, Chou TH, Feuer WJ. C57BL/6J, DBA/2J, and DBA/2J.Gpnmb mice have different visual signal processing in the inner retina. Mol Vis. 2010;16:2939–2947. [PMC free article] [PubMed] [Google Scholar]
- Porciatti V, Nagaraju M. Head-up tilt lowers IOP and improves RGC dysfunction in glaucomatous DBA/2J mice. Exp Eye Res. 2010;90:452–460. doi: 10.1016/j.exer.2009.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porciatti V, Pizzorusso T, Cenni MC, Maffei L. The visual response of retinal ganglion cells is not altered by optic nerve transection in transgenic mice overexpressing Bcl-2. Proc Natl Acad Sci U S A. 1996;93:14955–14959. doi: 10.1073/pnas.93.25.14955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porciatti V, Saleh M, Nagaraju M. The pattern electroretinogram as a tool to monitor progressive retinal ganglion cell dysfunction in the DBA/2J mouse model of glaucoma. Invest Ophthalmol Vis Sci. 2007;48:745–751. doi: 10.1167/iovs.06-0733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porciatti V, Ventura LM. Retinal ganglion cell functional plasticity and optic neuropathy: a comprehensive model. J Neuroophthalmol. 2012;32:354–358. doi: 10.1097/WNO.0b013e3182745600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prusky GT, Alam NM, Beekman S, Douglas RM. Rapid quantification of adult and developing mouse spatial vision using a virtual optomotor system. Invest Ophthalmol Vis Sci. 2004;45:4611–4616. doi: 10.1167/iovs.04-0541. [DOI] [PubMed] [Google Scholar]
- Rangaswamy NV, Zhou W, Harwerth RS, Frishman LJ. Effect of experimental glaucoma in primates on oscillatory potentials of the slow-sequence mfERG. Invest Ophthalmol Vis Sci. 2006;47:753–767. doi: 10.1167/iovs.05-1051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raz D, Perlman I, Percicot CL, Lambrou GN, Ofri R. Functional Damage to Inner and Outer Retinal Cells in Experimental Glaucoma. Investigative Ophthalmology & Visual Science. 2003;44:3675–3684. doi: 10.1167/iovs.02-1236. [DOI] [PubMed] [Google Scholar]
- Raz D, Seeliger MW, Geva AB, Percicot CL, Lambrou GN, Ofri R. The effect of contrast and luminance on mfERG responses in a monkey model of glaucoma. Invest Ophthalmol Vis Sci. 2002;43:2027–2035. [PubMed] [Google Scholar]
- Remtulla S, Hallett PE. A schematic eye for the mouse, and comparisons with the rat. Vision Res. 1985;25:21–31. doi: 10.1016/0042-6989(85)90076-8. [DOI] [PubMed] [Google Scholar]
- Riggs LA. Electroretinography. Vision Res. 1986;26:1443–1459. doi: 10.1016/0042-6989(86)90167-7. [DOI] [PubMed] [Google Scholar]
- Saleh M, Nagaraju M, Porciatti V. Longitudinal Evaluation of Retinal Ganglion Cell Function and IOP in the DBA/2J Mouse Model of Glaucoma. Invest Ophthalmol Vis Sci. 2007;48:4564–4572. doi: 10.1167/iovs.07-0483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandalon S, Konnecke B, Levkovitch-Verbin H, Simons M, Hein K, Sattler MB, Bahr M, Ofri R. Functional and structural evaluation of lamotrigine treatment in rat models of acute and chronic ocular hypertension. Exp Eye Res. 2013;115:47–56. doi: 10.1016/j.exer.2013.06.018. [DOI] [PubMed] [Google Scholar]
- Schmucker C, Schaeffel F. A paraxial schematic eye model for the growing C57BL/6 mouse. Vision Res. 2004;44:1857–1867. doi: 10.1016/j.visres.2004.03.011. [DOI] [PubMed] [Google Scholar]
- Schuettauf F, Quinto K, Naskar R, Zurakowski D. Effects of anti-glaucoma medications on ganglion cell survival: the DBA/2J mouse model. Vision Res. 2002;42:2333–2337. doi: 10.1016/s0042-6989(02)00188-8. [DOI] [PubMed] [Google Scholar]
- Sieving PA, Steinberg RH. Proximal retinal contribution to the intraretinal 8-Hz pattern ERG of cat. J Neurophysiol. 1987;57:104–120. doi: 10.1152/jn.1987.57.1.104. [DOI] [PubMed] [Google Scholar]
- Smith BJ, Wang X, Chauhan BC, Cote PD, Tremblay F. Contribution of retinal ganglion cells to the mouse electroretinogram. Documenta ophthalmologica. Advances in ophthalmology. 2014;128:155–168. doi: 10.1007/s10633-014-9433-2. [DOI] [PubMed] [Google Scholar]
- Trimarchi C, Biral G, Domenici L, Porciatti V, Bisti S. The flash- and pattern-electroretinogram generators in the cat: A pharmacological approach. Clinical Vision Sciences. 1990;6:19–24. [Google Scholar]
- Viswanathan S, Frishman LJ, Robson JG. The uniform field and pattern ERG in macaques with experimental glaucoma: removal of spiking activity. Invest Ophthalmol Vis Sci. 2000;41:2797–2810. [PubMed] [Google Scholar]
- Viswanathan S, Frishman LJ, Robson JG, Harwerth RS, Smith EL., 3rd The photopic negative response of the macaque electroretinogram: reduction by experimental glaucoma. Invest Ophthalmol Vis Sci. 1999;40:1124–1136. [PubMed] [Google Scholar]
- Viswanathan S, Frishman LJ, Robson JG, Walters JW. The photopic negative response of the flash electroretinogram in primary open angle glaucoma. Invest Ophthalmol Vis Sci. 2001;42:514–522. [PubMed] [Google Scholar]
- Weber AJ, Harman CD, Viswanathan S. Effects of optic nerve injury, glaucoma, and neuroprotection on the survival, structure, and function of ganglion cells in the mammalian retina. J Physiol. 2008;586:4393–4400. doi: 10.1113/jphysiol.2008.156729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams PA, Howell GR, Barbay JM, Braine CE, Sousa GL, John SW, Morgan JE. Retinal ganglion cell dendritic atrophy in DBA/2J glaucoma. PLoS One. 2013;8:e72282. doi: 10.1371/journal.pone.0072282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xia X, Wen R, Chou TH, Li Y, Wang Z, Porciatti V. Protection of Pattern Electroretinogram and Retinal Ganglion Cells by Oncostatin M after Optic Nerve Injury. PloS one. 2014;9:e108524. doi: 10.1371/journal.pone.0108524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang X, Chou TH, Ruggeri M, Porciatti V. A New Mouse Model of Inducible, Chronic Retinal Ganglion Cell Dysfunction Not Associated with Cell Death. Investigative Ophthalmology & Visual Science. 2013;54:1898–1904. doi: 10.1167/iovs.12-11375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yukita M, Machida S, Nishiguchi KM, Tsuda S, Yokoyama Y, Yasuda M, Maruyama K, Nakazawa T. Molecular, anatomical and functional changes in the retinal ganglion cells after optic nerve crush in mice. Doc Ophthalmol. 2015;130:149–156. doi: 10.1007/s10633-014-9478-2. [DOI] [PubMed] [Google Scholar]
- Zode GS, Bugge KE, Mohan K, Grozdanic SD, Peters JC, Koehn DR, Anderson MG, Kardon RH, Stone EM, Sheffield VC. Topical ocular sodium 4-phenylbutyrate rescues glaucoma in a myocilin mouse model of primary open-angle glaucoma. Invest Ophthalmol Vis Sci. 2012;53:1557–1565. doi: 10.1167/iovs.11-8837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zode GS, Kuehn MH, Nishimura DY, Searby CC, Mohan K, Grozdanic SD, Bugge K, Anderson MG, Clark AF, Stone EM, Sheffield VC. Reduction of ER stress via a chemical chaperone prevents disease phenotypes in a mouse model of primary open angle glaucoma. J Clin Invest. 2011;121:3542–3553. doi: 10.1172/JCI58183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zode GS, Sharma AB, Lin X, Searby CC, Bugge K, Kim GH, Clark AF, Sheffield VC. Ocular-specific ER stress reduction rescues glaucoma in murine glucocorticoid-induced glaucoma. J Clin Invest. 2014 doi: 10.1172/JCI69774. [DOI] [PMC free article] [PubMed] [Google Scholar]