
FIGURE 3.
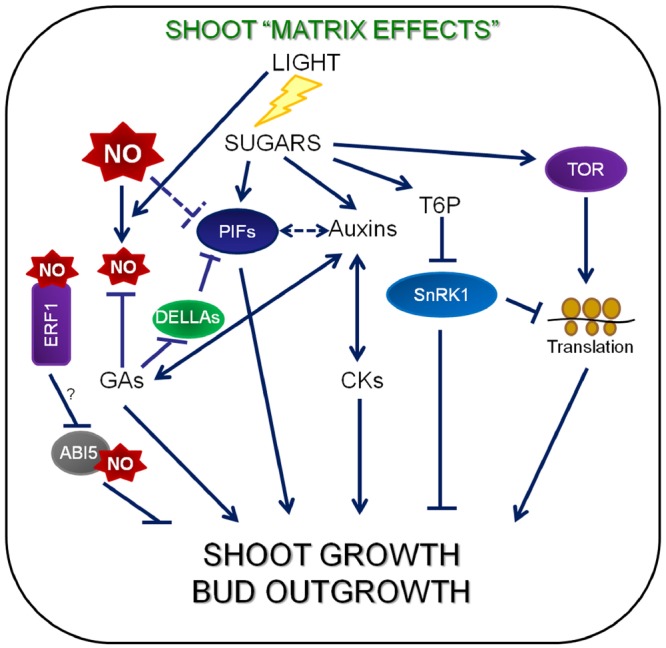
Molecular components of sugar signaling networks and their involvement in shoot growth. CKs, auxins and sugars function as long-distance signals. When photosynthesis takes place, sugars content increase in shoot and low sugars in roots implies that GAs are transported from the roots to the shoots and send sugars to the low sugar roots. Sucrose availability (high metabolic status) shows a good correlation with the level of plant T6P, which acts as an inhibitor of SnRK1. The active TOR kinase is important for C/S1-group bZIP protein synthesis and enhances translation and growth. Sucrose down-regulates the expression of C/S1-group bZIP at the translation level and the C/S1-bZIP transcription factor network is involved in the regulation of SnRK1 target genes (Lastdrager et al., 2014). The repression of ribosomal protein gene expression by SnRK1 (Baena-González et al., 2007) inhibits translation. Sucrose effects on auxin levels are more pronounced in roots than in shoots, suggesting sugars may impact auxin transport and/or conjugation pathways as well. DELLA and PIF proteins are good candidates for molecular hubs operating at the crossroads of many pathways (Ljung et al., 2015). There are NO and GA antagonist functions in the control of light-regulated photomorphogenesis through the balance between DELLAs and PIFs (Lozano-Juste and León, 2011). ERF1 has been suggested to function in light-regulated control of hypocotyl elongation, in a different way to the Cys-Arg/N-end rule pathway (Zhong et al., 2012).