

Abstract
3,4-Methylenedioxymethamphetamine (MDMA; ecstasy) toxicity may cause region-specific changes in serotonergic mRNA expression due to acute serotonin (5-hydroxytryptamine; 5-HT) syndrome. This hypothesis can be tested using in situ hybridization to detect the serotonin 5-HT2A receptor gene htr2a. In the past, such procedures, utilizing radioactive riboprobe, were difficult because of the complicated workflow that needs several days to perform and the added difficulty that the technique required the use of fresh frozen tissues maintained in an RNase-free environment. Recently, the development of short oligonucleotide probes has simplified in situ hybridization procedures and allowed the use of paraformaldehyde-prefixed brain sections, which are more widely available in laboratories. Here, we describe a detailed protocol using non-radioactive oligonucleotide probes on the prefixed brain tissues. Hybridization probes used for this study include dapB (a bacterial gene coding for dihydrodipicolinate reductase), ppiB (a housekeeping gene coding for peptidylprolyl isomerase B), and htr2a (a serotonin gene coding for 5-HT2A receptors). This method is relatively simply, cheap, reproducible and requires less than two days to complete.
Keywords: Neuroscience, Issue 103, Serotonin syndrome, in situ hybridization, MDMA toxicity, 5-HT2A receptor, gene expression, drug abuse
Introduction
Serotonin (5-hydroxytryptamine; 5-HT) syndrome is an acute neurologic disorder caused by 5-HT-promoting drugs such as antidepressants1, while also occurring in situations of MDMA use for recreational purposes2. Molecular mechanisms responsible for mood swings, learning and memory deficits that occur in association with the acute syndrome are not well understood3,4. In situ hybridization is a powerful research tool allowing the detection and quantification of specific mRNAs expressed potentially at a single-cell level. The conventional way to perform in situ hybridization is to utilize a radioactive-labeled riboprobe that specifically hybridizes the gene of interest. However, a major drawback is that the method requires complicated and time-consuming probe preparation and hybridization steps, as well as access to fresh frozen tissues maintained in an RNase-free environment5,6.
Oligonucleotide probes have been recently developed to hybridize shorter RNA fragments than those required for riboprobes7. Furthermore, the probes produce a low background signal without sacrificing specificity8. This newly-developed probe technology can be used in situ hybridization on paraformaldehyde-prefixed brain tissues commonly available in immunocytochemical laboratories.
Here, we describe a protocol for in situ hybridization using oligonucleotide probes on paraformaldehyde-prefixed rat brain and compare findings with those noted in a fresh frozen brain5,6. This protocol is used to test the hypothesis that MDMA substance abuse causes changes in 5-HT2A receptor gene htr2a mRNA in the brain. We began the procedure with MDMA treatment followed by paraformaldehyde tissue perfusion of the animal, in situ hybridization of the thr2a probe, and data analysis. Note that dapB (the bacterial gene coding for dihydrodipicolinate reductase) is used as a negative control, and ppiB (housekeeping gene coding for peptidylprolyl isomerase B) as a positive control.
Protocol
Animal use procedures described below were approved by The Institutional Animal Care and Use Committee (IACUC) at Ross University School of Veterinary Medicine and Florida Atlantic University. Although sterile techniques and gloves are required, the RNase-free environment is not necessary while using this protocol.
1. Preparation
- Tissue preparation and sectioning
- Assign rats randomly into three groups: fresh/saline (SAL; 0.9% NaCl), prefixed/SAL and prefixed/3,4-methylenedioxymethamphetamine (MDMA; see details in table 1).
- Administer three doses of 1 ml/kg SAL and 10 mg/kg MDMA intraperitoneally, respectively, to the SAL and MDMA groups at 2 hr interval. Allow animals to survive the treatment for 6 days.
- On day 7, administer 100 mg/kg ketamine in combination of 5 mg/kg xylazine intraperitoneally to the rats for a deep anesthesia (i.e., loss of corneal blink and tail pinch reflexes).
- For the fresh/SAL group, quickly decapitate rats and remove the brain for the fresh brain test (see details in the reference5).
- For the prefixed/SAL and prefixed/MDMA groups, make a cut along the sternum about 10 -12 cm and then expose the end of the sternum.
- Hold the end of the sternum with a hemostat and cut the diaphragm laterally on both sides with sharp scissors and then cut upward across the ribs and parallel to the lungs.
- With one hand, hold the ventral tip of the heart with small forceps. With the other hand, pierce the left ventricle with a 18 G syringe needle (note the adapter side of the needle is connected with a Y-shaped silicon hose to ice-cold SAL and paraformaldehyde containers and peristaltic pumps; see the reference9).
- Turn on the SAL pump at the medium level of the flow rate (~5 ml/min) and puncture the right atrium with the forceps to allow the escape of return circulation. Maintain SAL perfusion for 30 min.
- Turn on the paraformaldehyde pump at the medium level and allow the ice-cold 4% paraformaldehyde to perfuse the animal for 30 min.
- Remove perfusion instrument and decapitate the animal. Make a posteroanterior cut through the midline of the skull, and then flap the skull to expose the skull cavity 10.
- Remove the brain with a spatula from the skull cavity and place the brain in a 50 ml centrifuge tube containing 25 ml of 4% paraformaldehyde. Keep the brain within the paraformaldehyde-containing tube at 4 ºC refrigerator for 24 hr.
- Move the brain into a 50 ml centrifuge tube containing 25 ml of 30% sucrose solution at 4 ºC for 3 days. Finally, store the brain within plastic bags at -80 °C till use.
- Mount the brain on a chuck with embedding media and freeze the embedding media containing brain in a cryostat at temperature of -26 ºC. Cut the fresh frozen brain into 20 µm sections and paraformaldehyde-prefixed brain into 10-µm sections. Transfer the section to a RT microscope slide by touching the slide to the section.
- Air-dry at RT for 4 hr. Store the section slides in a tight sealed bag at -80 °C till use.
- Dehydration
- Take slides from the -80 °C freezer and assign sections into one of three tests: ht2a, ppiB (positive control) or dapB (negative control); mark and label sections with a ball pen (do not use a pencil). Submerge slides in 100% alcohol at RT for 5 min. Dry the slides for 5 min in a fume hood.
- Pretreatment
- Pipette 20 µl of the Pretreatment-1 reagent (inactivation of alkaline phosphatase) to the sections on slides. Place the slides on a rack rail in a moisture tray (note that the tray is covered with a lid to maintain a high humidity for preventing evaporation from the reaction solution).
- Gently shake the moisture tray on a horizontal shaker at a low speed for 10 min. Wash the slides twice with double-distilled water (ddH2O) for 2 min. Submerge the slide in a 200 ml beaker containing 150 ml of the Pretreatment-2 reagent (breaking of the cross-linkages induced by paraformaldehyde fixation). Boil the slides for a total of 5 min at 100 ºC (heat-induced restoration of RNA structure11).
- Wash the slides twice with ddH2O for 2 min. Place the slides in 100% alcohol for 5 min. Dry the slides for 5 min in the fume hood. Use a hydrophobic pen to draw a circle around the selected area in the tissue section (e.g., non-cortical structures, including thalamus and hypothalamus as outlined in Figure 2).
- Pipette 10 µl of the Pretreatment-3 reagent (increases in probe penetration) to each selected area, and cover the moisture tray with the lid. Place the tray in the hybridization oven equipped with the horizontal shaker and set the oven temperature at 40 ºC; gently shake the tray for 30 min. Wash the slides twice with ddH2O for 2 min.
2. Hybridization and Amplification
Pipette 10 µl of htr2a, ppiB and dapB probe reagents, respectively, on the selected areas. Cover the moisture tray with the lid to prevent evaporation of the reagent. Gently shake on the horizontal shaker set at a low speed. Incubate the slides at 40 ºC in the oven for 2 hr.
Wash the slides twice with a washing buffer for 2 min. Pipette 10 µl of the Amplification-1 reagent on each selected area, shake and incubate the slides at 40 ºC for 30 min.
Wash the slides twice with a washing buffer for 2 min. Pipette 10 µl of the Amplification-2 reagent on each selected area, shake and incubate the slides at 40 ºC for 15 min.
Wash the slides twice with a washing buffer for 2 min. Pipette 10 µl of the Amplification-3 reagent on each selected area, shake and incubate the slides at 40 ºC for 30 min.
Wash the slides twice with a washing buffer for 2 min. Pipette 10 µl of the Amplification-4 reagent on each selected area, shake and incubate the slides at 40 ºC for 15 min.
Wash the slides twice with a washing buffer for 2 min. Pipette 10 µl of the Amplification-5 reagent on each selected area, shake and incubate the slides at RT for 30 min.
Wash the slides twice with a washing buffer for 2 min. Pipette 10 µl of the Amplification-6 reagent on each selected area, shake and incubate the slides at RT for 15 min. Wash the slides twice with a washing buffer for 2 min.
3. Signal Detection
Pipette 10 µl of the Red reagent to each selected area (Note that the reagent is a mixture of Red-B and Red-A at a 1:60 ratio and should be used immediately). Place the slide in the moisture tray on the horizontal shaker at RT and shake gently at a low speed for 10 min.
Wash the slides twice with (ddH2O) for 10 min. Dry the slides at 60 ºC in the oven for 15 min. Place the slides in xylene under the fume hood for 5 min. Pipette 20 µl of xylene-based mounting media on each selected area and cover with a coverslip.
4. Image Capture
Take digital microphotographs of the selected areas (e.g., hypothalamus, Figures 1 and 3) with a magnification of 4× and 20× objective lenses. Save the image file.
5. Image Particle Analysis
Drag and drop the image file into the main ImageJ window. Go to the menu bar and select ‘Image’, next select ‘Type’ from the drop-down menu, and select ‘8-bit’.
Select ‘Adjust’, then select ‘Threshold’ to open the dialog box. Adjust the lower bar value in the dialog box so that the unwanted background is removed. Go to the menu bar and select ‘Analyze’, then select ‘Analyze Particles’ to open a second dialog box.
Set particle size at 10, next select ‘Show outlines’ and select ‘Display results” and ‘Summarize’ boxes. Lastly, click ‘OK’ to view particle data in the dialog boxes.
6. Spreadsheet Calculation
Enter the data into the Excel sheet and average the numbers of particles in the SAL group as a baseline. Calculate the percentage level of each section and make a graph accordingly (Figure 4).
Representative Results
Using the oligonucleotide RNA probes (<25 nt), hybridization can be detected as red dots in hypothalamic cells prepared from the paraformaldehyde-prefixed and fresh frozen brains. The htr2a mRNA molecules are present in some cells (indicated by solid arrows), but not others (open arrows). We observed that there is no difference between the paraformaldehyde-prefixed and the fresh frozen tissues (Figure 1). A successful hybridization can be evaluated first by the naked eye (Figure 2). We found that the selected area (marked with circles) hybridized with the dapB probe did not show a visible signal to the naked eye (Figures 2A-B). In contrast, the area hybridized with the ppiB probe showed signals homogenously distributed throughout the region examined (Figures 2C-D). While in the htr2a test, signals are present in certain nuclei (indicated by arrows; Figures 2E-F), although all the circled area has been hybridized with the htr2a probe. Interestingly, the signals in MDMA slides were relatively weak compared to SAL slides. Substantial details of hybridization signals can be evaluated under a microscopic instrument. There is no red dot (Figure 3A-B) in the cells hybridized with the dapB probe whereas signals can be seen throughout the microscopic field hybridized with the ppiB probe (Figure 3C-D). The htr2a signals are mainly located in certain nuclei of the hypothalamus (Figure 3E-F). We observed a reduction in htr2a signals in the brain tissues previously treated with systemic MDMA at 10 mg/kg, which can be quantitatively determined using the ImageJ analysis. As shown in Figure 4, we found a 20% reduction in htr2a expression.
In conclusion, our results showed that using oligonucleotide probes is specific, and suitable for the prefixed brain tissue. Furthermore, protocol procedures, including in situ hybridization, signal detection and data analysis can be completed within two days.
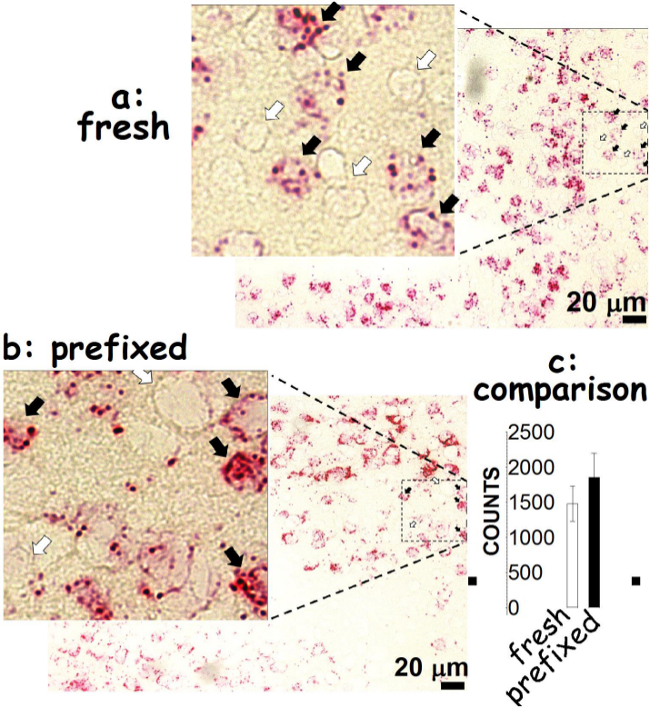 Figure 1. Comparison of the fresh and prefixed frozen tissues. Procedures used for fresh frozen brains have been described elsewhere5. Note that the fresh frozen sections were cut at 20 µm in thickness while the fixed frozen sections at 10 µm. The oligonucleotide probe was htr2a that hybridizes the 5-HT2A receptor mRNA in the hypothalamus. Solid arrows indicate cells containing htr2a mRNA molecules in the fresh frozen (A) and prefixed tissues (B). Open arrows, no htr2a mRNA molecules detected. Bar: 20 µm. Using NIH ImageJ analysis, htr2a was counted. Numbers of htr2a counts are not different between the fresh and paraformaldehyde-prefixed frozen tissues (C). Please click here to view a larger version of this figure.
Figure 1. Comparison of the fresh and prefixed frozen tissues. Procedures used for fresh frozen brains have been described elsewhere5. Note that the fresh frozen sections were cut at 20 µm in thickness while the fixed frozen sections at 10 µm. The oligonucleotide probe was htr2a that hybridizes the 5-HT2A receptor mRNA in the hypothalamus. Solid arrows indicate cells containing htr2a mRNA molecules in the fresh frozen (A) and prefixed tissues (B). Open arrows, no htr2a mRNA molecules detected. Bar: 20 µm. Using NIH ImageJ analysis, htr2a was counted. Numbers of htr2a counts are not different between the fresh and paraformaldehyde-prefixed frozen tissues (C). Please click here to view a larger version of this figure.
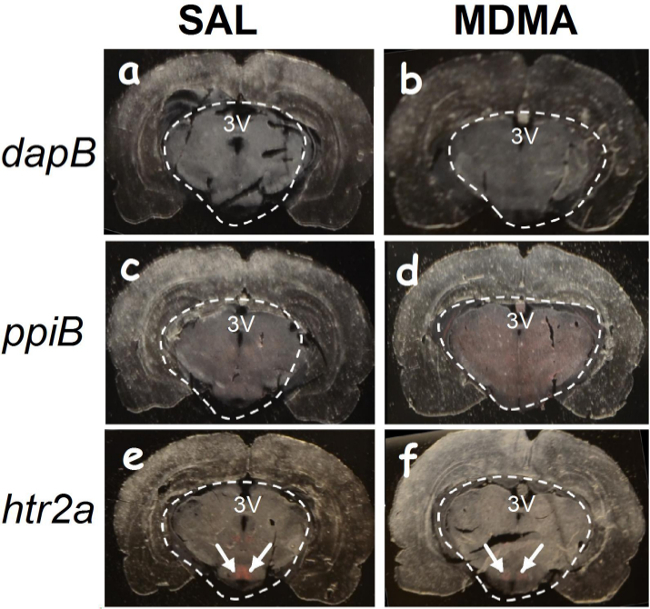 Figure 2. A naked-eye view of hybridization signals. Tissues were prefixed with paraformaldehyde in deeply anesthetized rats previously treated with saline (SAL) and 3,4-methylenedioxymethamphetamine (MDMA). The white-dashed lines indicate the region hybridized with oligonucleotide probes. The red color denotes mRNA signals to the naked eye. No red signal is observed while using the dapB probe hybridized to sections of SAL-pretreated (A) or MDMA-pretreated rats (B). In contrast, red signals are homogeneously distributed throughout the hypothalamic regions while using the ppiB probe (C-D). Interestingly, red signals produced by the htr2a probe are only in some regions (E-F; Arrows indicate the hypothalamic area). Stereotaxic coordinates: -3.50 ~ -3.80 relative to the bregma. 3V, the third ventricle. Please click here to view a larger version of this figure.
Figure 2. A naked-eye view of hybridization signals. Tissues were prefixed with paraformaldehyde in deeply anesthetized rats previously treated with saline (SAL) and 3,4-methylenedioxymethamphetamine (MDMA). The white-dashed lines indicate the region hybridized with oligonucleotide probes. The red color denotes mRNA signals to the naked eye. No red signal is observed while using the dapB probe hybridized to sections of SAL-pretreated (A) or MDMA-pretreated rats (B). In contrast, red signals are homogeneously distributed throughout the hypothalamic regions while using the ppiB probe (C-D). Interestingly, red signals produced by the htr2a probe are only in some regions (E-F; Arrows indicate the hypothalamic area). Stereotaxic coordinates: -3.50 ~ -3.80 relative to the bregma. 3V, the third ventricle. Please click here to view a larger version of this figure.
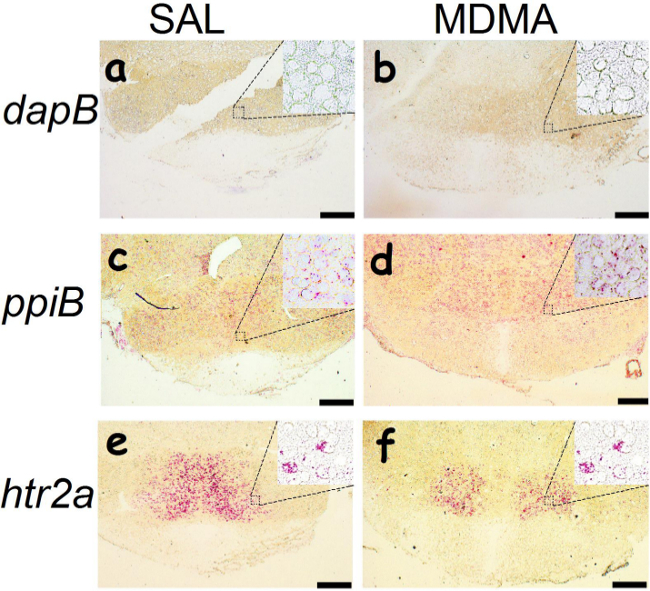 Figure 3. A light microscopic view of hybridization signals. Tissues were prefixed with paraformaldehyde in deeply anesthetized rats previously treated with saline (SAL) and 3,4-methylenedioxymethamphetamine (MDMA). Oligonucleotide probes are dapB (A-B), ppiB (C-D) and htr2a (E-F). Hybridization signals are identified as red dots using objective magnification powers ranging from 4× (Bars: 200 µm) to 20× (inset microphotographs). Please click here to view a larger version of this figure.
Figure 3. A light microscopic view of hybridization signals. Tissues were prefixed with paraformaldehyde in deeply anesthetized rats previously treated with saline (SAL) and 3,4-methylenedioxymethamphetamine (MDMA). Oligonucleotide probes are dapB (A-B), ppiB (C-D) and htr2a (E-F). Hybridization signals are identified as red dots using objective magnification powers ranging from 4× (Bars: 200 µm) to 20× (inset microphotographs). Please click here to view a larger version of this figure.
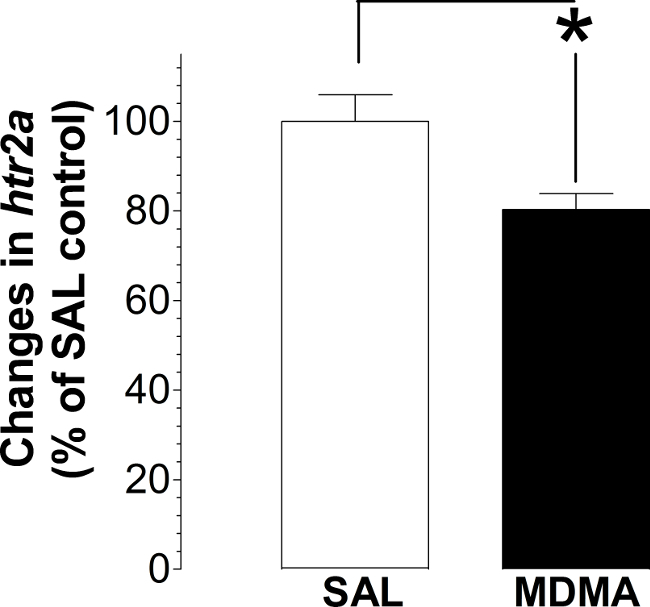 Figure 4. Effect on hypothalamic htr2a expression in rats previously treated with MDMA. Samples were assayed in duplicates and repeated three times. Data are expressed as a mean percentage ± s.e.m. of the SAL group (a total of 8 rats used in this study). *P <0.05 vs. the SAL group using Student’s t test.
Figure 4. Effect on hypothalamic htr2a expression in rats previously treated with MDMA. Samples were assayed in duplicates and repeated three times. Data are expressed as a mean percentage ± s.e.m. of the SAL group (a total of 8 rats used in this study). *P <0.05 vs. the SAL group using Student’s t test.
| Fresh | Paraformaldehyde-prefixeda | |||
| SALb | SALb | MDMAc | ||
| Target probe (htr2a) | √ | √ | √ | |
| Positive probe (ppiB) | – | √ | √ | |
| Negative probe (dapB) | – | √ | √ |
√, indicates the group examined with oligonucleotide probe in this study. –, not examined; a, each group contains 4 different animals; b, the group pretreated intraperitoneally with physical saline (SAL; 0.9% NaCl); c, pretreated intraperitoneally with 10 mg/kg MDMA.
Table 1. Experimental design
Discussion
One of the major concerns of in situ hybridization test is to choose appropriate techniques used for RNA preservation because of RNase enzymes. It is well known that these enzymes are widely present in the cells and environment which can affect the results. However, enzyme activity can be quickly distinguished by placing the tissue in the dry ice/alcohol solution5,12. Although quick preservation is critical for hybridization using a riboprobe13, little is known about experimental conditions for oligonucleotide probes. In this study using oligonucleotide probes, we found that there was no difference between the paraformaldehyde-prefixed and fresh brains quickly preserved in the ice/alcohol solution. This suggests that brain RNA prefixed by paraformaldehyde had no significant effect on the quality of hybridization using the oligonucleotide probe. Since the prefixed brain is easily sectioned compared to the fresh brain, this protocol provides an alternative tissue preparation for the hybridization test.
We believe that keeping the brain tissue sections inside the sealed moisture tray with constant shaking on the horizontal shaker is essential for successful hybridization. Failure to follow those steps may result in uneven hybridization or high background. To eliminate possibilities of false-negative or false-positive results, tests should include necessary control slides. We found that the dapB probe did not show a visible signal. This is not surprising since dapB is a bacterial gene that is unlikely found in mammalian cells. In contrast, the area hybridized with the ppiB probe showed signals homogenously distributed throughout the region examined. This is predictable since the housekeeping gene should be present in any kind of mammalian cells. It has been suggested that 5-HT2A mRNA is distributed heterogeneously in the brain nuclei, including the hypothalamus14,15. As expected, htr2a signals are present in certain nuclei in the region of the hypothalamus although the probe has been applied to all of the selected areas. It should be kept in mind that the use of alcohol should be avoided after the Red reagent (step 3.1) since red signals can be easily decolored by alcohol. We found that, after cellular RNA stained with the Red reagent, cells cannot be counterstained with hematoxylin or crystal violet although the exact cause of counterstaining failure is unknown. In summary, a characteristic distribution of htr2a mRNA supports the idea that these oligonucleotide probes are gene-specific despite the shorter sequence relative to other probes16,17.
Background signals are another major concern, which likely affects the quality and faithfulness of data analysis. Unlike the high background produced by riboprobes13, oligonucleotide probes normally show a very low background. Thus, image can be easily and reasonably quantified using publicly-available NIH ImageJ software. In this study, we showed an example of this technique by examining changes in htr2a expression in response to systemic MDMA at 10 mg/kg. We demonstrated that MDMA caused a 20% reduction in htr2a expression, supporting previous reports18.
In addition to the three genes outlined above, this protocol has been validated with two other gene probes (trh, thyrotropin-release hormone gene, and trhr, thyrotropin-releasing hormone receptor gene; data not shown), demonstrating a high sensitivity and accuracy in detecting mRNA levels. Since most reactions require only 10 µl of reagents, the cost of an assay is also significantly reduced, compared to radioactive riboprobes. Furthermore, it is important to note that this protocol does not require the RNase-free environment. In conclusion, this protocol improves the speed of in situ hybridization assay, and increases reproducibility with relatively low cost.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgments
This study was supported by the NIH grant (R15DA029863), the Florida Atlantic University undergraduate research grant (M30014) and the Ross University School of Veterinary Medicine research grant. We would like to thank the National Institute on Drug Abuse (Rockville, MD) for providing (±)3,4-methylenedioxymethamphetamine (±MDMA) for this work.
References
- Muzyk AJ, Jakel RJ, Preud'homme X. Serotonin syndrome after a massive overdose of controlled-release paroxetine. Psychosomatics. 2010;51:437–442. doi: 10.1176/appi.psy.51.5.437. [DOI] [PubMed] [Google Scholar]
- Davies O, Batajoo-Shrestha B, Sosa-Popoteur J, Olibrice M. Full recovery after severe serotonin syndrome, severe rhabdomyolysis, multi-organ failure and disseminated intravascular coagulopathy from MDMA. Heart Lung. 2014;43:117–119. doi: 10.1016/j.hrtlng.2013.11.009. [DOI] [PubMed] [Google Scholar]
- Bosch OG, et al. Verbal memory deficits are correlated with prefrontal hypometabolism in 18FDG PET of recreational MDMA users. PLoS On. 2013;8:e61234. doi: 10.1371/journal.pone.0061234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smithies V, Broadbear J, Verdejo-Garcia A, Conduit R. Dysfunctional overnight memory consolidation in ecstasy users. J Psychopharmaco. 2014;28:751–762. doi: 10.1177/0269881114525673. [DOI] [PubMed] [Google Scholar]
- Winzer-Serhan UH, Broide RS, Chen Y, Leslie FM. Highly sensitive radioactive in situ hybridization using full length hydrolyzed riboprobes to detect α2 adrenoceptor subtype mRNAs in adult and developing rat brain. Brain Res Brain Res Proto. 1999;3:229–241. doi: 10.1016/s1385-299x(98)00043-9. [DOI] [PubMed] [Google Scholar]
- Zoeller RT, Fletcher DL, Butnariu O, Lowry CA, Moore FL. N-ethylmaleimide (NEM) can significantly improve in situ hybridization results using 35S-labeled oligodeoxynucleotide or complementary RNA probes. J Histochem Cytoche. 1997;45:1035–1041. doi: 10.1177/002215549704500712. [DOI] [PubMed] [Google Scholar]
- Wang F, et al. RNAscope: a novel in situ RNA analysis platform for formalin-fixed, paraffin-embedded tissues. J Mol Diag. 2012;14:22–29. doi: 10.1016/j.jmoldx.2011.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H, et al. RNAscope for in situ detection of transcriptionally active human papillomavirus in head and neck squamous cell carcinoma. J Vis E. 2014. [DOI] [PMC free article] [PubMed]
- Gage GJ, Kipke DR, Shain W. Whole animal perfusion fixation for rodents. J Vis E. 2012. [DOI] [PMC free article] [PubMed]
- Mishra A, et al. Imaging pericytes and capillary diameter in brain slices and isolated retinae. Nat Proto. 2014;9:323–336. doi: 10.1038/nprot.2014.019. [DOI] [PubMed] [Google Scholar]
- Shi SR, Cote RJ, Taylor CR. Antigen retrieval techniques: current perspectives. J Histochem Cytoche. 2001;49:931–937. doi: 10.1177/002215540104900801. [DOI] [PubMed] [Google Scholar]
- Qin Y, Heine VM, Karst H, Lucassen PJ, Joels M. Gene expression patterns in rat dentate granule cells: comparison between fresh and fixed tissue. J Neurosci Method. 2003;131:205–211. doi: 10.1016/j.jneumeth.2003.08.010. [DOI] [PubMed] [Google Scholar]
- Carter BS, Fletcher JS, Thompson RC. Analysis of messenger RNA expression by in situ hybridization using RNA probes synthesized via in vitro transcription. Method. 2010;52:322–331. doi: 10.1016/j.ymeth.2010.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mijnster MJ, et al. Regional and cellular distribution of serotonin 5-hydroxytryptamine2a receptor mRNA in the nucleus accumbens, olfactory tubercle, and caudate putamen of the rat. J Comp Neuro. 1997;389:1–11. [PubMed] [Google Scholar]
- Li Q, et al. Brain region-specific alterations of 5-HT2A and 5-HT2C receptors in serotonin transporter knockout mice. J Neuroche. 2003;84:1256–1265. doi: 10.1046/j.1471-4159.2003.01607.x. [DOI] [PubMed] [Google Scholar]
- Horner KA, Gilbert YE, Noble ES. Differential regulation of 5-HT2A receptor mRNA expression following withdrawal from a chronic escalating dose regimen of D-amphetamine. Brain Re. 2011;1390:10–20. doi: 10.1016/j.brainres.2011.03.033. [DOI] [PubMed] [Google Scholar]
- Mocci G, Jimenez-Sanchez L, Adell A, Cortes R, Artigas F. Expression of 5-HT2A receptors in prefrontal cortex pyramidal neurons projecting to nucleus accumbens. Potential relevance for atypical antipsychotic action. Neuropharmacolog. 2014;79:49–58. doi: 10.1016/j.neuropharm.2013.10.021. [DOI] [PubMed] [Google Scholar]
- Reneman L, et al. The acute and chronic effects of MDMA ('Ecstasy') on cortical 5-HT2A receptors in rat and human. 2002;26:387–396. doi: 10.1016/S0893-133X(01)00366-9. [DOI] [PubMed] [Google Scholar]