

Abstract
AIM: To study the effect of staurosporine (ST) on the cell cycle of human gastric cancer cell lines MGC803 and SGC7901.
METHODS: Cell proliferation was evaluated by trypan blue dye exclusion method. Apoptotic morphology was observed under a transmission electron microscope. Changes of cell cycle and apoptotic peaks of cells were determined by flow cytometry. Expression of P21WAF1 gene was examined using immunohistochemistry and RT-PCR.
RESULTS: The growth of MGC803 and SGC7901 cells was inhibited by ST. The inhibitory concentrations against 50% cells (IC50) at 24 h and 48 h were 54 ng/ml and 23 ng/ml for MGC803, and 61 ng/ml and 37 ng/ml for SGC7901. Typical apoptotic bodies and apoptotic peaks were observed 24 h after cells were treated wth ST at a concentration of 200 ng/ml. The percentage of cells at G0/G1 phase was decreased and that of cells at G2/M was increased significantly in the group treated wth ST at the concentrations of 40 ng/ml, 60 ng/ml, 100 ng/ml for 24 h, compared with the control group (P < 0.01). The expression levels of P21WAF1 gene in both MGC803 and SGC7901 cells were markedly up-regulated after treatment with ST.
CONCLUSION: ST can cause arrest of gastric cancer cells at G2/M phase, which may be one of the mechanisms that inhibit cell proliferation and cause apoptosis in these cells. Effect of ST on cells at G2/M phase may be attributed to the up-regulattion of P21WAF1 gene.
INTRODUCTION
Protein kinase C (PKC) isoforms are serine/threonine kinases involved in signal transduction pathways that govern a wide range of physiological processes including differentiation, proliferation, gene expression, membrane transport and organization of cytoskeletal and extracellular matrix proteins[1]. PKC isoforms are often overexpressed in disease states such as cancer. The important role they play in the processes relevant to neoplastic transformation, carcinogenesis and tumor cell invasion renders PKC a potentially suitable target for anticancer therapy[2].
Staurosporine (ST), a microbial alkaloid (indolocarbazole produced by Streptomyces sp.), has been shown to be a potent inhibitor of a wide range of protein kinases, including different serine/threonine and tyrosine protein kinases, which acts by competing with the ATP-binding region of the kinase catalytic domain[3]. Staurosporine has been reported to exert various pharmacological actions involving protein kinase C both in vivo and in vitro, such as diminishing thrombin enhanced procoagulant activity, reducing carbachol-induced insulin secretion[4-7]. PKC function is altered in some neoplasias, and this dysfunction has been related to uncontrolled proliferation[8,9].
Completion of the cell cycle requires the coordination of a variety of macromolecular syntheses, assemblies, and movements[10]. Coordination of the timing and order of these processes are achieved by a regulatory system that responds to checkpoints at major transitions in the cycle. Two key checkpoints are at the G1/S and G2/M phase transitions. G1 and G2 cyclins, the cyclin-dependent kinases and cycle kinase inhibitors (CKI) are responsible for controlling the transitions. Here, we investigated the effect of the potent phospholipid/ Ca2+-dependent kinase (PKC) inhibitor, ST, on cell cycle of human gastric cancer cell lines, MGC803 and SGC7901.
MATERIALS AND METHODS
Cell lines and cell culture
Human gastric mucinous adenocarcinoma cell line, MGC803, was obtained from Department of Immunity, China Medical University and human gastric carcinoma metastatic lymph node cell line, SGC7901, was obtained from Laboratory of Oncology, the First Affiliated Hospital of China Medical University. The derived cell lines were grown in RPMI 1640 medium supplemented with 10% heat-inactivated fetal calf serum, 50 U/ml penicillin and 50 µg/ml streptomycin. The cells were maintained at 37 °C in a humidified atmosphere containing 5% CO2. Viability of the cells used in these experiments was consistently more than 95% when evaluated by the trypan blue exclusion method. Staurosporine was purchased from Sigma Company (St. Louis, USA).
Analysis of cell viability
Effect of ST on cell growth and viability was measured by directly counting the number of cells by means of trypan blue dye exclusion. Cells at a density of 2.5×105/ml were seeded onto 24-well plates, and then treated with ST at different concentrations for 24 and 48 h. Control cells were also cultured at the same time. Cell proliferation and inhibition curves were drawn, and the inhibitory concentration against 50% cells (IC50) was determined.
Cell morphological analysis with transmission electron microscope
Transmission electronic microscope was employed to observe the effect of ST on cell apoptosis. Cells (1×106 ) were collected after being treated with ST at 200 ng/ml for 24 h, washed twice with PBS solution, centrifuged for 5 min at 1 500 rpm, then fixed in 2% glutaraldehyde at 4 °C for 72 h, then placed in 1% phosphotungstic acid. The cells were desiccated with gradients, and embedded with EPON-812. Ultrathin sections were prepared and observed after double staining with uranium and plumbum under a transmission electron microscope. Each experiment was repeated four times. At the same time, the cells without treatment with ST were used as a control group.
Cell cycle analysis
Flow cytometry was employed to determine the DNA content and the apoptotic peaks of the cells. The cells were seeded on 100 mm-dishes and grown in RPMI-1640 supplemented with 10% FCS. After treated with ST at the concentrations of 40 ng/ml, 60 ng/ml, 100 ng/ml, 200 ng/ml, 500 ng/ml for 24 h respectively, the cells were harvested, trypsinized, washed with D-PBS, fixed by adding slowly 2 ml of cold 70% ethanol into the tube and then stored at 4 °C. After fixation, the cells were washed, centrifuged, and resuspended in 0.05 mg/ml propidium iodide (Fluka Co, MILWAUKEE, USA), 100 units/ml RNase (Fluka Co, MILWAUKEE, USA) in PBS. The sample was incubated at room temperature for 30 min, and analyzed on a FACSCalibur (BD PharMingen, FRANKLIN Lakes, USA). Cell cycle data originally obtained with Cell Quest software (BD PharMingen, FRANKLIN Lakes, USA) were re-analyzed using MODFIT software (Verity Software House, Topsham, USA). At the same time negative controls were constructed.
Immunohistochemistry
P21 protein was detected by immunohistochemistry using specific monoclonal antibody (Maxin Co, Fuzhou, China). After treated with ST at the concentrations 40 ng/ml, 60 ng/ml, 100 ng/ml for 24 h, respectively, the cells were harvested. At the same time the negative controls were constructed. The cells were smeared on a slide with cell smear centrifugal apparatus, and stained using conventional S-P immunohistochemical method. Color was developed with DAB reagent and counterstained with hematoxylin. The cells were observed under a light microscope. The cells clearly showing brown color in their nuclei and plasma were considered to be positive for P21 protein.
Semiquantitative reverse transcription-PCR
The expression of P21 mRNA was determined by RT-PCR. After treated with ST at the concentration of 60 ng/ml, the cells were harvested. At the same time the negative controls were constructed. The cells were washed, and total RNA was extracted with the Qiagen RNA isolation kit (GIBCO Co., New York, USA). Aliquots containing 5 µg/ml of RNA from each treatment were used for the first-strand cDNA synthesis (TaKaRa Co., Dalian, China). In each reaction, 100 µl solution containing 3 µM of random hexamers, 25 mM Tris-HCl, 37 mM KCl, 1.5 mM MgCl2, 10 mM DTT, 0.25 mM dNTP, 40 units of RNAsin,an RNase inhibitor, 50 U/ml Super Taq DNA polymerase, and 200 units of reverse transcriptase was used. The annealing mixture was incubated at room temperature for 15 min, and then incubated in a water bath at 41 °C for 60 min. The reverse transcriptase enzyme was inactivated by heating the solution to 95 °C for 5 min. PCR was then carried out using the Perkin-Elmer PCR reaction kit and primers. The PCR was performed using a thermocycler for 30 cycles consisting of denaturation at 94 °C for 1 min, annealing at 57 °C for 1 min, and extension at 72 °C for 2 min. The PCR products were separated on 2% agarose gel. The primers used for PCR were as follow: P21 sense (5’-GGG GAC AGC AGA GGA AGA C-3’), P21 antisense (5’-CGG CGT TTG GAG TGG TAG A-3’); β-actin sense (5’-GAT TGC CTC AGG ACA TTT CTG- 3’), β-actin antisense (5’-GAT TGC TCA GGA CAT TTC TG-3’). Gene primers were synthesized by Beijing Oake Company (Beijing, China).
Statistical analysis
Student’s t-test was used to compare the difference between control and ST treated groups. Data of cell growth were presented as -x ± s. A P value less than 0.05 was considered statistically significant.
RESULTS
ST inhibited proliferation of MGC803 and SGC7901 cells in a time-dependent and concentration-dependent manner
In this study, the exponentially grown MGC803 and SGC7901 cells were treated with 40 ng/ml, 60 ng/ml, 100 ng/ml ST, respectively, and the cell proliferation was measured 24 and 48 h after ST addition. Figures 1 and 2 show the cell proliferation curves at various ST concentrations. The inhibition of proliferation of MGC803 and SGC7901 cells by ST was clearly observed in a time-dependent and concentration-dependent manner. The IC50 was 54 ng/ml and 23 ng/ml for MGC803 and 61 ng/ml and 37 ng/ml for SGC7901 at 24 and 48 h.
Figure 1.
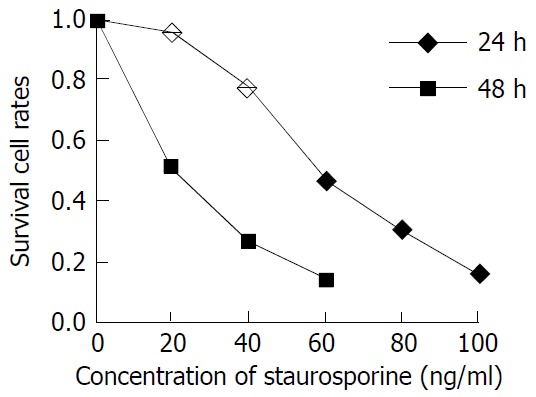
Inhibition of staurosporine on MGC-803 cell proliferation.
Figure 2.
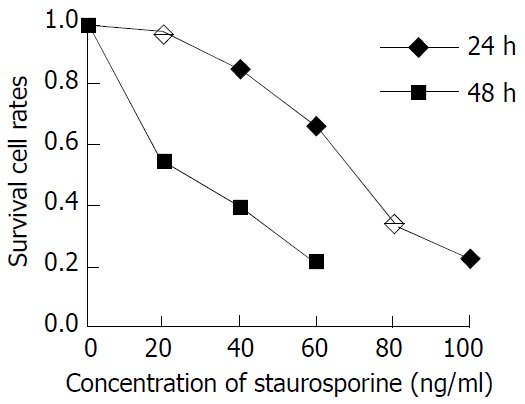
Inhibition of staurosporine on SGC-7901 cell proliferation.
Morphological observation of ST treatment effects
Cell morphological changes were observed under a transmission electron microscope after treatment with ST at the concentration of 200 ng/ml for 24 h. The ultrastructural appearances showed the typical changes in the cell morphology, including blebbing of the plasma membrane, chromatin condensation and formation of apoptotic bodies. Figure 3 shows the morphological changes of MGC803 and SGC7901 cells under a electron microscope after treatment with ST.
Figure 3.
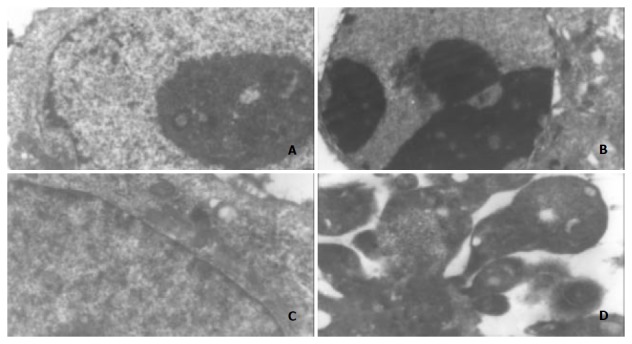
Morphological changes in MGC803 and SGC7901 cells under electron microscope after treatment with ST (200 ng/ml), ×5000. A: MGC803 control cells, B: 200 ng/ml ST-induced MGC803 cells, C: SGC7901 control cells, D: 200 ng/ml ST-induced SGC7901 cells. Note: chromatin condensation and formation of apoptotic bodies.
ST induced MGC803 and SGC7901 cells G2/M phase arrest
The effects of ST on cell cycle progression, population distribution and apoptotic incidence in` MGC803 and SGC7901 cells were determined using flow cytometry. ST-induced effects were detected by comparing the cell cycle profiles between ST treated and untreated cells. Notably, the cells demonstrated significant G2/M arrest 24 h after ST treatment (P < 0.01), in comparison to untreated cells. Interestingly, the S phase population was also increased, but to a lesser extent as compared with untreated cells. The percentage of cells in the S, G1, and G2/M phases are shown in Table 1. Apoptotic peaks were observed and cell apoptotic incidence was determined 24 h after treatment with ST at the concentrations of 200 ng/ml and 500 ng/ml. The apoptotic incidence increased to 50.2% and 89.6% in MGC803 cells, and 34.6% and 80.7% in SGC7901 cells after ST treatment. Figure 4 shows ST-induced apoptosis in MGC803 and SGC7901 cells.
Table 1.
Effect of ST on cell cycle of MGC803 and SGC7901 cells(-x ± s)
|
MGC803 |
SGC7901 |
|||||||
| G0/G1(%) | S(%) | G2/M(%) | AI | G0/G1(%) | S(%) | G2/M(%) | AI | |
| Control | 54.3 ± 3.1 | 15.2 ± 0.6 | 13.5 ± 0.2 | 3.1 ± 0.2 | 52.5 ± 4.4 | 10.1 ± 0.6 | 13.5 ± 2.2 | 2.8 ± 0.2 |
| 40 ng/ml | 23.6 ± 1.8a | 13.9 ± 1.1 | 22.6 ± 4.0a | 3.8 ± 0.9 | 27.1 ± 1.4a | 12.4 ± 0.1 | 21.9 ± 2.6a | 3.3 ± 0.3 |
| 60 ng/ml | 11.6 ± 0.7a | 12.6 ± 2.8 | 35.5 ± 0.4a | 4.0 ± 0.3 | 17.0 ± 3.4a | 13.4 ± 2.0 | 39.5 ± 4.9a | 3.7 ± 0.6 |
| 100ng/ml | 3.3 ± 0.2a | 10.9 ± 1.7 | 36.8 ± 5.5a | 5.2 ± 0.4 | 13.7 ± 0.7a | 12.7 ± 0.9 | 38.4 ± 3.1a | 4.4 ± 1.1 |
AI, apoptotic incidence;
P < 0.01 vs Control.
Figure 4.
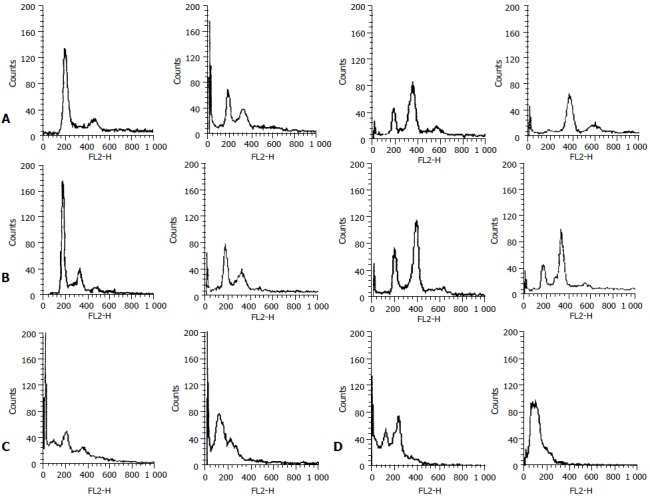
Effect of ST on cell cycle of human gastric cancer cells. A: Changes of cell cycle in MGC803 cells after ST-treatment for 24h, B: Changes of cell cycle in SGC7901 cells after ST-treatment for 24 h, C: ST-induced apoptosis in MGC-803 cells, D: ST-induced apoptosis in SGC-7901 cells.
ST resulted in an increase of P21 expression
The effect of ST on P21 protein levels was determined by immunohistochemistry analysis. Normally, P21 was not expressed in MGC803 and SGC7901 cells. However, P21 was significantly expressed, the positive rates were 19.3%, 26.6%, 31.8% in MGC803 cells, and 20.5%, 24.2%, 30.3% in SGC7901 cells after treatment with ST at the concentration 40 ng/ml, 60 ng/ml, 100 ng/ml for 24 h. Figure 5 shows the levels of P21 expression in MGC803 and SGC7901 cells by immunohistochemistry.
Figure 5.
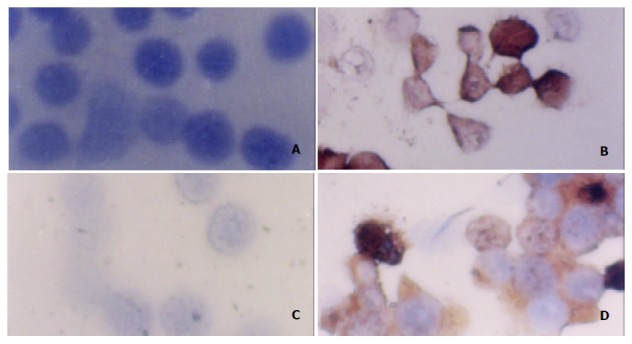
Expression of P21 protein in MGC803 and SGC7901 cells after ST-treatment as determined by immunohistochemistry (SP×400). A: MGC803 control cells, B: MGC803 cells treated with 60 ng/ml ST, C: SGC7901 control cells, D: SGC7901 cells treated with 60 ng/ml ST.
ST upregulated P21WAF1 expression
Normally, P21WAF1 mRNA was almost not expressed in MGC803 and SGC7901 cells when detected by RT-PCR. However, treatment with ST at the concentration of 60 ng /ml induced the upregulation of P21WAF1 mRNA in MGC803 and SGC7901 cells. Figures 6 and 7 show that P21WAF1 mRNA expression was upregulated in MGC803 and SGC7901 cells after treatment with ST.
Figure 6.
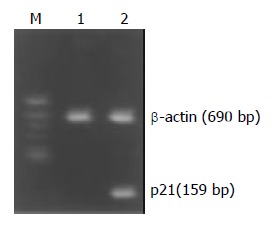
Effect of ST treatment on P21WAF1 mRNA expression in MGC803 cells. M: DNA Marks, 1: control cells, 2: cells treated with 60 ng/ml ST.
Figure 7.
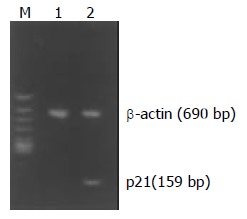
Effect of ST treatment on P21WAF1 mRNA expression in SGC7901 cells. M: DNA Marks, 1: control cells, 2: cells treated with 60 ng/ml ST.
DISCUSSION
Cancer cells have been found to be different from normal cells in many important characteristics, including loss of differentiation and decrease of apoptosis[11,12]. These differences did not arise simply from uncontrolled cellular growth but rather from a process of cellular evolution[13-15]. Cell cycle plays an important role in the modulation of tumor cell growth, and attention has been paid to preventing unlimited proliferation of tumor cells by cell cycle control[16].
Twenty years ago, staurosporine (ST) was isolated from bacteria and identified as a potent inhibitor of PKC activity[17-19]. In this study, the proliferation of human gastric cell lines, MGC803 and SGC7901, was significantly inhibited in a time-and concentration-dependent manner and classical apoptosis (sub-diploid peak on flow cytometry, and typical morphological changes) was observed after treated with ST. ST also blocked the G2/M phase of the cell cycle. In order to examine the distribution of DNA content, the cells treated with ST were detected using flow cytometry. The first peak (2C C = haploid DNA content) was produced by the cell population in G1 phase, the second (4C) was produced by the cells in G2/M phase with or without cells in G1 phase at a higher DNA ploidy (tetraploidy, G1t), and the third (8C) was produced by cells in G2 phase at a higher DNA ploidy (tetraploidy, G2t). The results presented in this study indicated that the cells were blocked in G2/M phase. These results lead to the suggestion that, ST can make damaged cells stagnate at G2/M phase of cell cycle, and inhibit proliferation of cells, allowing them to repair the damage. If this happens, cells will reenter the cycle, otherwise, they will undergo apoptosis or death. More importantly, ST affects the selectively on G2/M phase of these cells and participates in the regulation of the cell cycle, and thus the blocking effect of ST on cells in G2/M phase might be the mechanism of its antitumor effect. These observations may provide some useful information that ST can be used as an antitumor agent.
The present study was undertaken to delineate the mechanism of ST effect on the G2/M phase arrest of cells, and techniques including inmmunohistochemistry and RT-PCR were used to detect the expression of P21WAF1 protein and gene. Our results revealed that P21WAF1 expression was low in MGC803 and SGC7901 cells. However, ST significantly increased the expression of P21 in MGC803 and SGC7901 cells. Cell cycle events, including microtubule dynamics, membrane organization and DNA synthesis, were tightly controlled, and specific changes were induced at particular points in the eukaryotic cell cycle during cell proliferation[20,21]. Negative controls on cell cycle progression were exerted during development, differentiation, senescence, and cell death. These negative controls might play an important role in preventing tumorigenesis. P21WAF1, the pioneer member of P21 family of cyclin-CDK inhibitor class of proteins, has been implicated as a growth arrest mediator in cell terminal differentiation and apoptosis[22]. Regulation of cell cycle progression is orchestrated by a family of CDKs, which can be negatively regulated by CDK inhibitors, such as P21WAF1. P21WAF1 is a downstream effector of p53 that mediates both G1 and G2/M phase arrest. Mechanistically, the P21WAF1-mediated arrest of the G2/M cell cycle transition has been suggested to include a P21WAF1-CDK2 and P21WAF1-PCNA protein interaction[23,24]. P21 is the strongest kinase inhibitor, and has been found to inhibit the CDK4/6-cyclinD complex when overexpressed, leading to growth arrest and, under some conditions, to apoptosis[25-27]. Although our knowledge on cell cycle checkpoints is still limited, it is clear that many such control points existed within the cell cycle and that they played a major role in maintaining the integrity of the genome[28,29]. At least two checkpoints could detect DNA damage, one at the G1-S transition and another at the G2-M transition[30-32]. ST up-regulates P21WAF1, which then affects cell growth by repressing G2/M phase, probably through interaction with cyclin-CDK complex. However, this hypothesis needs to be further investigated.
In conclusion, the present study demonstrates that ST can effectively inhibit the proliferation of human gastric cancer MGC803 and SGC7901 cells, induces apoptosis, blocks these cells in G2/M phase, and up-regulates the P21WAF1 expression. This study provides some experimental data for the use of ST in the treatment of gastric carcinoma.
Footnotes
Supported by The China State Key Basic Research Program, No. G1998051203
Edited by Xia HHX and Wang XL
References
- 1.Caponigro F, French RC, Kaye SB. Protein kinase C: a worthwhile target for anticancer drugs. Anticancer Drugs. 1997;8:26–33. doi: 10.1097/00001813-199701000-00003. [DOI] [PubMed] [Google Scholar]
- 2.Aiello LP, Bursell SE, Clermont A, Duh E, Ishii H, Takagi C, Mori F, Ciulla TA, Ways K, Jirousek M, et al. Vascular endothelial growth factor-induced retinal permeability is mediated by protein kinase C in vivo and suppressed by an orally effective beta-isoform-selective inhibitor. Diabetes. 1997;46:1473–1480. doi: 10.2337/diab.46.9.1473. [DOI] [PubMed] [Google Scholar]
- 3.Toledo LM, Lydon NB, Elbaum D. The structure-based design of ATP-site directed protein kinase inhibitors. Curr Med Chem. 1999;6:775–805. [PubMed] [Google Scholar]
- 4.Zaugg K, Rocha S, Resch H, Hegyi I, Oehler C, Glanzmann C, Fabbro D, Bodis S, Pruschy M. Differential p53-dependent mechanism of radiosensitization in vitro and in vivo by the protein kinase C-specific inhibitor PKC412. Cancer Res. 2001;61:732–738. [PubMed] [Google Scholar]
- 5.Gescher A. Analogs of staurosporine: potential anticancer drugs. Gen Pharmacol. 1998;31:721–728. doi: 10.1016/s0306-3623(98)00069-x. [DOI] [PubMed] [Google Scholar]
- 6.6Xia M, Xue SB, Xu CS. Shedding of TNFR1 in regenerative liver can be induced with TNF alpha and PMA. World J Gastroenterol. 2002;8:1129–1133. doi: 10.3748/wjg.v8.i6.1129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Li MS, Li PF, He SP, Du GG, Li G. The promoting molecular mechanism of alpha-fetoprotein on the growth of human hepatoma Bel7402 cell line. World J Gastroenterol. 2002;8:469–475. doi: 10.3748/wjg.v8.i3.469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ouyang GL, Li QF, Peng XX, Liu QR, Hong SG. Effects of tachyplesin on proliferation and differentiation of human hepatocellular carcinoma SMMC-7721 cells. World J Gastroenterol. 2002;8:1053–1058. doi: 10.3748/wjg.v8.i6.1053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Han Y, Han ZY, Zhou XM, Shi R, Zheng Y, Shi YQ, Miao JY, Pan BR, Fan DM. Expression and function of classical protein kinase C isoenzymes in gastric cancer cell line and its drug-resistant sublines. World J Gastroenterol. 2002;8:441–445. doi: 10.3748/wjg.v8.i3.441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gescher A. Staurosporine analogues - pharmacological toys or useful antitumour agents. Crit Rev Oncol Hematol. 2000;34:127–135. doi: 10.1016/s1040-8428(00)00058-5. [DOI] [PubMed] [Google Scholar]
- 11.Chen B, He L, Savell VH, Jenkins JJ, Parham DM. Inhibition of the interferon-gamma/signal transducers and activators of transcription (STAT) pathway by hypermethylation at a STAT-binding site in the P21WAF1 promoter region. Cancer Res. 2000;60:3290–3298. [PubMed] [Google Scholar]
- 12.Zeng YX, el-Deiry WS. Regulation of P21WAF1/CIP1 expression by p53-independent pathways. Oncogene. 1996;12:1557–1564. [PubMed] [Google Scholar]
- 13.Wu Q, Kirschmeier P, Hockenberry T, Yang TY, Brassard DL, Wang L, McClanahan T, Black S, Rizzi G, Musco ML, et al. Transcriptional regulation during P21WAF1/CIP1-induced apoptosis in human ovarian cancer cells. J Biol Chem. 2002;277:36329–36337. doi: 10.1074/jbc.M204962200. [DOI] [PubMed] [Google Scholar]
- 14.Kovalsky O, Lung FD, Roller PP, Fornace AJ. Oligomerization of human Gadd45a protein. J Biol Chem. 2001;276:39330–39339. doi: 10.1074/jbc.M105115200. [DOI] [PubMed] [Google Scholar]
- 15.Mihalik R, Uher F, Pocsik É L, Benczur M, Kopper L. Detection of Drug-induced Apoptosis by Flow Cytometry after Alkaline Extraction of Ethanol Fixed Cells. Pathol Oncol Res. 1996;2:78–83. doi: 10.1007/BF02893956. [DOI] [PubMed] [Google Scholar]
- 16.Heerdt BG, Houston MA, Mariadason JM, Augenlicht LH. Dissociation of staurosporine-induced apoptosis from G2-M arrest in SW620 human colonic carcinoma cells: initiation of the apoptotic cascade is associated with elevation of the mitochondrial membrane potential (deltapsim) Cancer Res. 2000;60:6704–6713. [PubMed] [Google Scholar]
- 17.Salvioli S, Dobrucki J, Moretti L, Troiano L, Fernandez MG, Pinti M, Pedrazzi J, Franceschi C, Cossarizza A. Mitochondrial heterogeneity during staurosporine-induced apoptosis in HL60 cells: analysis at the single cell and single organelle level. Cytometry. 2000;40:189–197. doi: 10.1002/1097-0320(20000701)40:3<189::aid-cyto3>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- 18.He SW, Shen KQ, He YJ, Xie B, Zhao YM. Regulatory effect and mechanism of gastrin and its antagonists on colorectal carcinoma. World J Gastroenterol. 1999;5:408–416. doi: 10.3748/wjg.v5.i5.408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bernard B, Fest T, Prétet JL, Mougin C. Staurosporine-induced apoptosis of HPV positive and negative human cervical cancer cells from different points in the cell cycle. Cell Death Differ. 2001;8:234–244. doi: 10.1038/sj.cdd.4400796. [DOI] [PubMed] [Google Scholar]
- 20.Stokke T, Smedshammer L, Jonassen TS, Blomhoff HK, Skarstad K, Steen HB. Uncoupling of the order of the S and M phases: effects of staurosporine on human cell cycle kinases. Cell Prolif. 1997;30:197–218. doi: 10.1046/j.1365-2184.1997.00087.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zeng ZC, Jiang GL, Wang GM, Tang ZY, Curran WJ, Iliakis G. DNA-PKcs subunits in radiosensitization by hyperthermia on hepatocellular carcinoma hepG2 cell line. World J Gastroenterol. 2002;8:797–803. doi: 10.3748/wjg.v8.i5.797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bertrand R, Solary E, O'Connor P, Kohn KW, Pommier Y. Induction of a common pathway of apoptosis by staurosporine. Exp Cell Res. 1994;211:314–321. doi: 10.1006/excr.1994.1093. [DOI] [PubMed] [Google Scholar]
- 23.Fang JY, Lu YY. Effects of histone acetylation and DNA methylation on P21( WAF1) regulation. World J Gastroenterol. 2002;8:400–405. doi: 10.3748/wjg.v8.i3.400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wartenberg M, Fischer K, Hescheler J, Sauer H. Modulation of intrinsic P-glycoprotein expression in multicellular prostate tumor spheroids by cell cycle inhibitors. Biochim Biophys Acta. 2002;1589:49–62. doi: 10.1016/s0167-4889(01)00185-9. [DOI] [PubMed] [Google Scholar]
- 25.Yuste VJ, Sánchez-López I, Solé C, Encinas M, Bayascas JR, Boix J, Comella JX. The prevention of the staurosporine-induced apoptosis by Bcl-X(L), but not by Bcl-2 or caspase inhibitors, allows the extensive differentiation of human neuroblastoma cells. J Neurochem. 2002;80:126–139. doi: 10.1046/j.0022-3042.2001.00695.x. [DOI] [PubMed] [Google Scholar]
- 26.Shimizu T, Takahashi N, Tachibana K, Takeda K. Complex regulation of CDK2 and G1 arrest during neuronal differentiation of human prostatic cancer TSU-Prl cells by staurosporine. Anticancer Res. 2001;21:893–898. [PubMed] [Google Scholar]
- 27.Narita Y, Asai A, Kuchino Y, Kirino T. Actinomycin D and staurosporine, potent apoptosis inducers in vitro, are potentially effective chemotherapeutic agents against glioblastoma multiforme. Cancer Chemother Pharmacol. 2000;45:149–156. doi: 10.1007/s002800050023. [DOI] [PubMed] [Google Scholar]
- 28.Chen X, Lowe M, Herliczek T, Hall MJ, Danes C, Lawrence DA, Keyomarsi K. Protection of normal proliferating cells against chemotherapy by staurosporine-mediated, selective, and reversible G(1) arrest. J Natl Cancer Inst. 2000;92:1999–2008. doi: 10.1093/jnci/92.24.1999. [DOI] [PubMed] [Google Scholar]
- 29.Sausville EA, Johnson J, Alley M, Zaharevitz D, Senderowicz AM. Inhibition of CDKs as a therapeutic modality. Ann N Y Acad Sci. 2000;910:207–21; discussion 221-2. doi: 10.1111/j.1749-6632.2000.tb06710.x. [DOI] [PubMed] [Google Scholar]
- 30.Shchepina LA, Popova EN, Pletjushkina OY, Chernyak BV. Respiration and mitochondrial membrane potential are not required for apoptosis and anti-apoptotic action of Bcl-2 in HeLa cells. Biochemistry (Mosc) 2002;67:222–226. doi: 10.1023/a:1014418231692. [DOI] [PubMed] [Google Scholar]
- 31.Swe M, Sit KH. Staurosporine induces telophase arrest and apoptosis blocking mitosis exit in human Chang liver cells. Biochem Biophys Res Commun. 1997;236:594–598. doi: 10.1006/bbrc.1997.6985. [DOI] [PubMed] [Google Scholar]
- 32.Stepczynska A, Lauber K, Engels IH, Janssen O, Kabelitz D, Wesselborg S, Schulze-Osthoff K. Staurosporine and conventional anticancer drugs induce overlapping, yet distinct pathways of apoptosis and caspase activation. Oncogene. 2001;20:1193–1202. doi: 10.1038/sj.onc.1204221. [DOI] [PubMed] [Google Scholar]