

Abstract
Purpose
Heterochromatin protein 1γ (HP1γ) interacts with chromosomes by binding to lysine 9-methylated histone H3 or DNA/RNA. HP1γ is involved in various biological processes. The purpose of this study is to gain an understanding of how HP1γ functions in these processes by identifying HP1γ-binding proteins using mass spectrometry.
Materials and Methods
We performed affinity purification of HP1γ-binding proteins using G1/S phase or prometaphase HEK293T cell lysates that transiently express mock or FLAG-HP1γ. Coomassie staining was performed for HP1γ-binding complexes, using cell lysates prepared by affinity chromatography FLAG-agarose beads, and the bands were digested and then analyzed using a mass spectrometry.
Results
We identified 99 HP1γ-binding proteins with diverse cellular functions, including spliceosome, regulation of the actin cytoskeleton, tight junction, pathogenic Escherichia coli infection, mammalian target of rapamycin signaling pathway, nucleotide excision repair, DNA replication, homologous recombination, and mismatch repair.
Conclusion
Our results suggested that HP1γ is functionally active in DNA damage response via protein-protein interaction.
Keywords: HP1γ, Protein interaction, DNA damage response
Introduction
Members of the heterochromatin protein 1 (HP1) family contain three functional domains: N-terminal chromo domain, hinge region, and C-terminal chromo-shadow domain [1]. HP1 functions as a regulator for gene expression that induces the heterochromatin structure by binding with meH3K9 [2]. The members of the family, HP1α, HP1β, and HP1γ, are specifically localized in the cell. HP1α and HP1β are primarily localized within centromeric heterochromatin, while HP1γ is localized in both heterochromatic and euchromatic sites [3,4]. Additionally, chromatin immunoprecipitation has been used to associate HP1γ with the DNA of actively transcribed genes [5]. Through its ability to regulate the chromatin structure, HP1γ functions as a transcriptional regulator. For example, HP1γ is responsible for chromatin-mediated human immunodeficiency virus 1 transcriptional silencing and post-integration latency [6]. Moreover, tumor necrosis factor α transcription is silenced during endotoxin tolerance by a cooperative interaction of histone and DNA methylation via HP1 and G9a [7].
Although many studies have shown the functional activity of HP1γ in gene regulation, the exact mechanisms are not fully understood. Genomic instability in chromosomes is a major characteristic of cancer cells. Loss of HP1γ in mice leads to genomic instability during meiosis via altered cell cycle progression [8], suggesting that HP1γ participates in the maintenance of genomic stability. In a recent study, chromatin binding protein HP1α was shown to contribute to chromosome dynamics during early mitosis and chromosome segregation via its interaction with Borealin, one component of the chromosomal passenger complex [9]. In addition to accurate chromosome segregation, HP1 also plays a role in DNA damage response that preserves the chromosome structure [10]. For example, a recent study reported that, in collaboration with the tumor suppressor BRCA1, HP1γ contributes to recovery from DNA damage [10]. Whilst there is evidence that HP1γ participates in maintaining the number of chromosomes and their structure, the precise mechanisms by which HP1 prevents genomic instability remain unclear. Therefore, in order to achieve a more complete understanding of how HP1γ functions in a diversity of biological processes and pathways, we performed affinity purification and identified several HP1γ-binding proteins. Our results demonstrate that HP1γ may be a multifunctional protein, including the preservation of genome integrity.
Materials and Methods
1. Cell culture
Human embryonic kidney (HEK) 293T cells were obtained from American Type Culture Collection and grown (at 5% CO2 and 37°C) in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (GE Healthcare Hyclone, Logan, UT) and 1% anti-biotics/anti-mycotics.
2. Plasmids and transfection
FLAG-HP1γ expression plasmids were used, as previously described [10]. Transient transfection was performed using FuGENE 6 reagent (Roche Applied Science, Indianapolis, IN).
3. Antibodies
Anti-FLAG antibody was purchased from Sigma (St. Louis, MO), and γH2AX antibody (EMD Millipore, Billerica, MA) was used as previously described [11]. The antibodies used for immunoblotting were as follows: anti-phospho-H3 Ser 10 (EMD Millipore), anti-Bloom syndrome protein (BLM) (Abcam, Cambridge, MA), and anti-proliferating cell nuclear antigen (PCNA) (Abcam).
4. Cell synchronization
Cells were synchronized at the late G1 phase using a double thymidine block method [12]. Briefly, the cells were plated in 150-mm diameter Petri dishes, and thymidine was added to a final concentration of 2 mM after cell adherence. The cells were cultured for 16 hours. After removal of the thymidine and incubation for 10 hours in fresh medium, thymidine was again added to a final concentration of 2 mM and the cells were cultured for an additional 16 hours. After removal of the thymidine, synchronized cells were cultured in fresh medium and collected at different times for cell cycle analysis and immunoblotting. The cells were synchronized in the prometaphase with 17 hours of nocodazole treatment and then released into fresh medium for further incubation. Immunoblotting analysis was performed to confirm the indicated phases of the cell cycle using antibodies against phospho-H3 Ser10, a mitotic marker.
5. shRNA plasmid construction
The shRNA for HP1γ was generated using a pSUPER. retro.puro, an H1 promoter-driven RNAi retroviral vector (Oligoengine, Seattle, WA). The shRNA primers were designed to target HP1γ (5´-GATCGACGTGTAGTGAATG-3´).
6. Immunoprecipitation and mass spectrometry
The cells were grown at 80%-90% confluence in 150-mm culture dishes, then washed with ice-cold phosphate-buffered saline. The harvested cells were lysed with ice-cold Tris lysis buffer (175 mM Tris, pH 8.0, 150 mM NaCl, 1% NP-40, 1 mM dithiothreitol, 0.5 mM phenylmethylsulfonyl fluoride, and complete protease/phosphatase-inhibitor cocktail, according to Thermo Scientifics’ instructions). The cellular extract (10 mg of total lysates for each sample) was mixed with 30 μL of EZview Red ANTI-FLAG M2 Affinity Gel (Sigma) and incubated for 2 hours at 4°C, with rotation. The bound proteins were eluted from the beads by boiling for 5 minutes in 2× sodium dodecyl sulfate sample buffer containing 5% (v/v) β-mercaptoethanol, and resolved by a one-dimensional polyacrylamide gel electrophoresis. Coomassie-stained HP1γ-interacting proteins were in-gel digested with trypsin (Promega, Madison, WI) and analyzed by capillary column liquid chromatography-tandem mass spectrometry (LC-MS/MS), using LTQ-Orbitrap mass spectrometry systems (Thermo Finnigan, San Jose, CA) equipped with nanospray ionization sources. Tandem mass spectra were interpreted by the Sorcerer program using the SEQUEST algorithm, and subsequently by the Scaffold program (version Scaffold_4.0.5, Proteome Software Inc., Portland, OR). Peptide and protein identifications were accepted if they could be established with greater than 95% probability and contained at least two identified unique peptides. All searches were performed against the human protein sequence database (IPI human DB v3.87 fasta).
For coimmunoprecipitation, the cell lysates were incubated with anti-HP1γ antibodies (EMD Millipore) and the immunoprecipitated complex was analyzed by immunoblotting using anti-BLM or anti-PCNA antibody.
7. Immunofluorescence
The HeLa cells were strained with appropriate primary antibodies, followed by incubation with Alexa Fluor 488- or Alexa Fluor 546-conjugated secondary antibodies (Invitrogen, Carlsbad, CA). The cell nuclei were stained with 4´,6-diamidino-2-phenylindole. The samples were analyzed under an LSM700 confocal laser scanning microscope (Carl Zeiss, Thornwood, NY).
Results
1. Establishment of the interactome of HP1γ
To identify new HP1γ-binding proteins, we performed affinity purification, using G1/S phase or prometaphase HEK293T cell lysates that transiently express mock or FLAG-HP1γ. Coomassie staining was performed for HP1γ-binding complexes using cell lysates prepared by affinity chromatography FLAG-agarose beads (Fig. 1). We detected several bands that indicated elution from each transfected cell line. The bands were digested and mass spectrometry analysis was performed. A comparison of the binding proteins found in cell lysates eluted from each transfected cell line is shown in Supplementary Table 1. Most of the identified proteins were novel interacting proteins, which had not been previously reported. Subcellular distribution analysis showed that the proteins with HP1γ interaction were present in all major cellular compartments, including the nucleus, cytoplasm, and plasma membrane (Fig. 2A, Supplementary Table 2). These proteins were found to be involved in various biological processes, such as splicing, regulation of the actin cytoskeleton, tight junctions, pathogenic Escherichia coli infection, the mammalian target of rapamycin signaling pathway, nucleotide excision repair, DNA replication, homologous recombination, and mismatch repair (Fig. 2B, Supplementary Table 3). We also analyzed the domains in the HP1γ-binding proteins and found that HP1γ associates with many proteins via several functional domains (Fig. 2C, Supplementary Table 4). Therefore, our interactome analysis demonstrated that HP1γ has several potential roles in various biological processes.
Fig. 1.

Identification of HP1γ-binding proteins. Coomassie staining of affinity-purified FLAG-HP1γ complexes in G1/S phase or prometaphase HEK293T cells. The cell extracts prepared from each transfected cell were subjected to affinity purification using FLAG affinity beads. The elutes were analyzed by SDS-PAGE and visualized by Coomassie staining. The Coomassie-stained proteins immunoprecipitated with anti-FLAG antibodies in 1-3 lanes were in-gel digested with trypsin and analyzed by LC-MS/MS. The numbers on the left-hand side indicate molecular weights. Lane 1, the FLAG-(empty) vector-transfected HEK293T cell lysates as a control; lane 2, the FLAG-HP1γ vector-transfected HEK293T cell lysates in the G1/S phase; lane 3, the FLAG-HP1γ vector-transfected HEK293T cell lysates in the prometaphase. The immunoprecipitated FLAG-HP1γ (FLAG-HP1γ) and light chain of immunoglobulin (IgG light chain) are indicated by arrows. Immunoblotting using antibodies against phospho-H3 Ser10, a mitotic marker, was performed to discriminate the indicated phases of cell cycle. HP1γ, heterochromatin protein 1γ; SDS-PAGE, sodium dodecyl sulphate-polyacrylamide gel electrophoresis; LC-MS/MS, liquid chromatography-tandem mass spectrometry; IB, immunoblot.
Fig. 2.
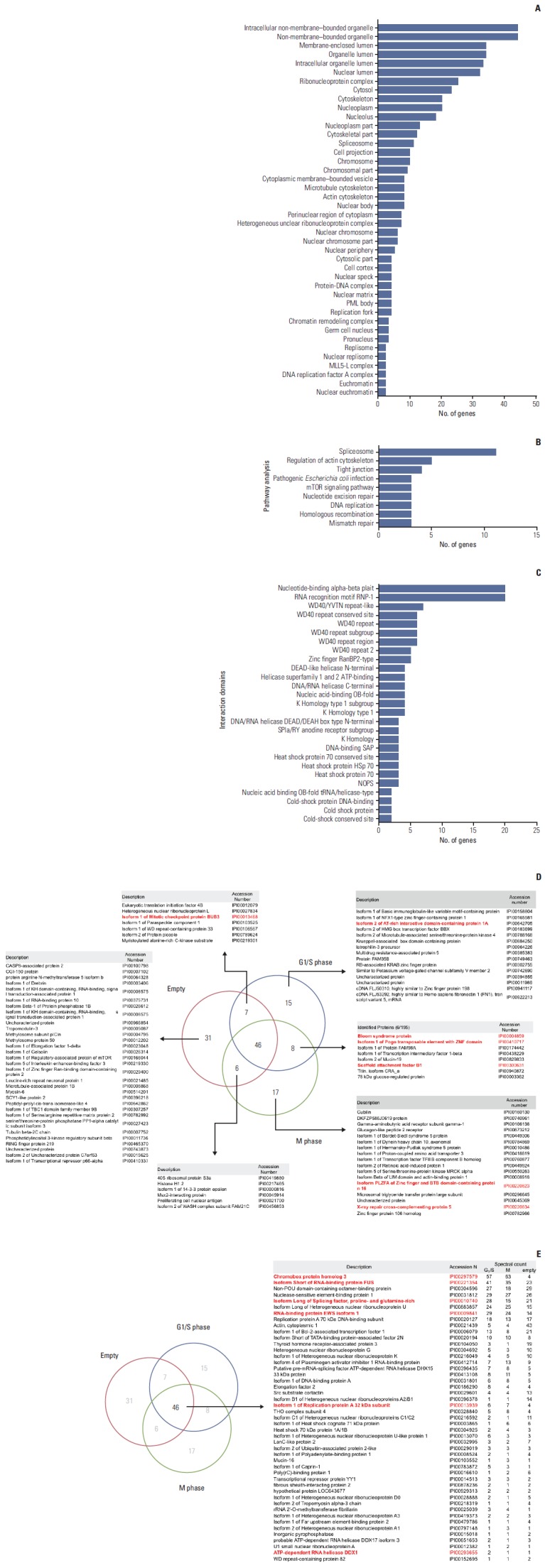
Proteins that interact with heterochromatin protein 1γ (HP1γ). (A) Diagram showing the cellular components of identified proteins that interact with HP1γ. (B) Diagram showing related molecular functions of these identified proteins. (C) Diagram showing domains that associate with HP1γ in identified proteins, visualized using Cytoscape. (D, E) Nonproportional Venn diagrams showing subsets of identified proteins in this study. Subset areas are not proportional to the actual relative subset sizes. Number of proteins identified in the immunoprecipitated complexes using the FLAG-(empty) vector-transfected cell lysates as a control (subset Empty), the FLAG-HP1γ vector-transfected cell lysates in the G1/S phase (subset G1/S), or the FLAG-HP1γ vector-transfected cell lysates in the M phase (subset M) are illustrated in the diagrams. The protein identities in each subset are described in the tables. In the subset tables. HP1γ-interacting protein that are implicated in DNA damage response pathways are marked in red.
2. The function of HP1γ in the DNA damage response pathway
By bioinformatic analysis, we identified 13 proteins implicated in the DNA damage response pathway, which comprise about 13% of potential candidates that interact with HP1γ (Table 1, Fig. 2D and E, Supplementary Table 5). First, we subtracted 31 proteins only in mock lysates, from a total of 130 proteins identified in our proteomic study, and then 99 proteins were considered as HP1γ-interacting partner candidates (Fig. 2D, Supplementary Table 1). Among these 99 HP1γ-interacting candidate proteins, we accepted 13 proteins if their unique spectral counts could be detected at greater than or equal to approximately two-fold ratio compared to the control (fold ratio of G1-S or mitosis to mock control) (Fig. 2D and E, Supplementary Table 1), and functional relationship with DNA damage response pathways were validated by gene ontology analysis (Table 1, Supplementary Table 5) or literature survey (Table 1). Among the proteins identified as HP1γ-interacting partners, this data set, following validation analyses in Table 1, could postulate a functional connection between HP1γ and DNA damage response.
Table 1.
HP1γ interacting proteins function in the DNA damage response
| Description | Accession No. | M. W. (kDa) | Unique spectral count |
Significance validation |
|||||
|---|---|---|---|---|---|---|---|---|---|
| G1-S | M | Empty | Fold ratio (G1-S or M/control) | GO | Article references | Co-IP Ex. | |||
| Bloom syndrome protein (BLM) | IPI00004859 | 159 | 2 | 1 | 0 | O | O | - | O |
| Chromobox protein homolog 3 (CBX3, the bait in this study) | IPI00297579 | 21 | 57 | 63 | 4 | O | O | - | Not done |
| Isoform 1 of Replication protein A 32 kDa subunit (RPA2) | IPI00013939 (+2) | 29 | 6 | 7 | 4 | O | O | - | Not done |
| Isoform 2 of AT-rich interactive domain-containing protein 1A (ARID1A) | IPI00642705 (+2) | 218 | 2 | N.D. | N.D. | O | O | - | Not done |
| Isoform Long of splicing factor, proline- and glutamine-rich (SFPQ) | IPI00010740 | 76 | 28 | 15 | 21 | O | O | - | Not done |
| Scaffold attachment factor B1 (SAFB) | IPI00300631 (+5) | 103 | 5 | 1 | N.D. | O | O | - | Not done |
| X-ray repair cross-complementing protein 5 (XRCC5) | IPI00220834 | 83 | N.D. | 2 | N.D. | O | O | - | Not done |
| Isoform 1 of Mitotic checkpoint protein BUB3 (BUB3) | IPI00013468 (+2) | 37 | 2 | N.D. | 1 | O | O | - | Not done |
| ATP-dependent RNA helicase DDX1 (DDX1) | IPI00293655 (+1) | 82 | 2 | 1 | 1 | O | - | Δ [13] | Not done |
| Isoform 1 of Pogo transposable element with ZNF domain (POGZ) | IPI00410717 (+5) | 155 | 2 | 2 | N.D. | O | - | O [14] | Not done |
| Isoform PLZFA of Zinc finger and BTB domain-containing protein 16 (PLZF) | IPI00220823 (+1) | 62 | N.D. | 2 | N.D. | O | - | Δ [15] | Not done |
| Isoform Short of RNA-binding protein FUS (FUS) | IPI00221354 (+3) | 53 | 41 | 35 | 23 | O | - | Δ [16] | Not done |
| RNA-binding protein EWS isoform 1 (EWS) | IPI00009841 (+4) | 69 | 29 | 24 | 14 | O | - | Δ [17] | Not done |
| Proliferating cell nuclear antigen (PCNA) | IPI00021700 | 29 | N.D. | 1 | 3 | - | O | O [1] | O |
HP1γ, heterochromatin protein 1γ; M. W., molecular weight; N.D., not detected; O, validated by fold ratio, GO, or coimmunoprecipitation experiments (Co-IP Ex). In Article references, references indicated as O report its interaction with HP1γ and functional relation to DNA damage response; references indicated as Δ report only functional relation to DNA damage response.
Therefore, we investigated the potential role of HP1γ in the DNA damage response pathway. Next, we tested whether HP1γ was indeed interacting with these identified proteins related to DNA damage response pathway. Interaction of HP1γ with BLM, the putative interacting partner identified by mass spectrometry (No. 3 in Supplementary Table 1), was detected by coimmunoprecipitation analysis (Fig. 3A). One of the surrogate proteins implicated in DNA damage response, PCNA, was not identified as an HP1γ-interacting protein by our mass spectrometric analysis (Fig. 2D and E, Supplementary Table 1), yet it was capable to interact with HP1γ in a coimmunoprecipitation experiment (Fig. 3A). One possible explanation for the difference in the interaction results of tandem mass spectrometry and coimmunoprecipitation experiments is that the antibody used affects the efficacy of protein-protein interaction in such a way that the specificity of anti-HP1γ antibody is higher for endogenous HP1γ proteins, but lower for anti-FLAG antibody for the recombinant FLAG-HP1γ proteins is lower. Another possible explanation is the discrepancy between two separate experiments. Without knowing the specificity of these antibodies or the difference between experiments, such issues are difficult to address.
Fig. 3.
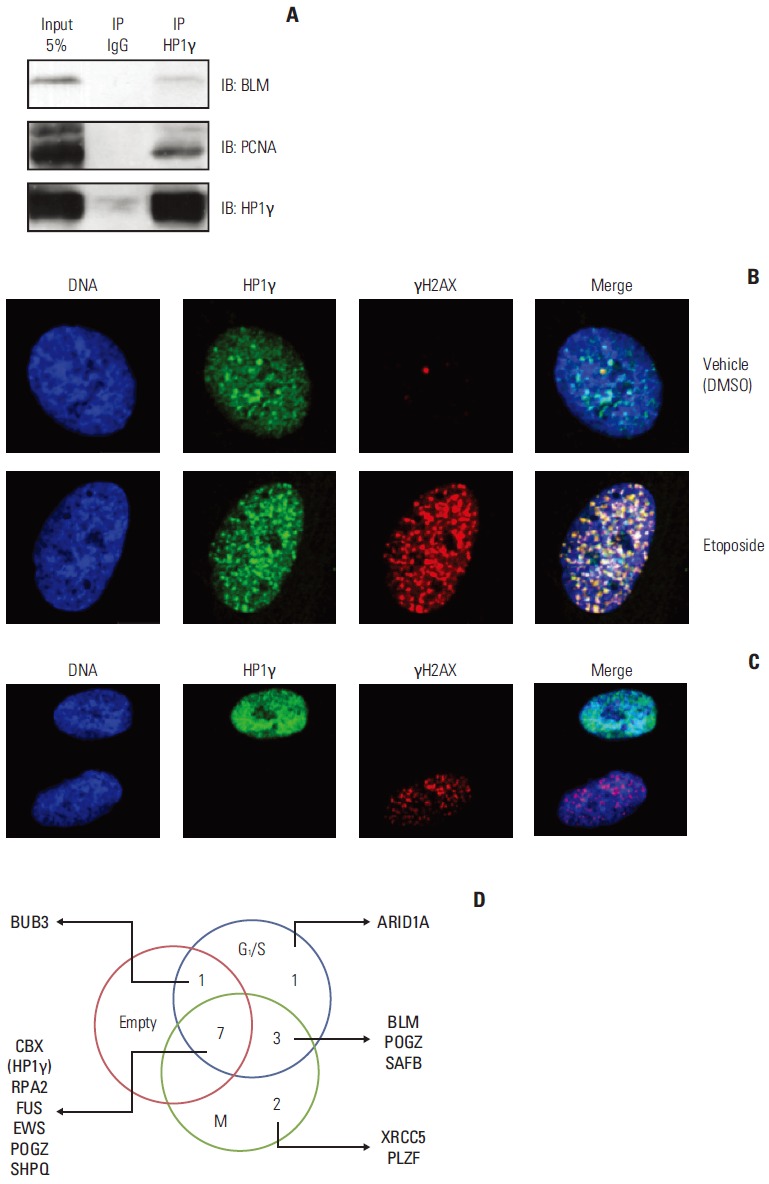
The function of heterochromatin protein 1γ (HP1γ) in the DNA damage response pathway. (A) Bloom syndrome protein (BLM) and proliferating cell nuclear antigen (PCNA) were associated with endogenous HP1γ proteins. BLM and PCNA were coimmunoprecipitated with anti-HP1γ antibodies using HEK293T cell lysates and immunoblotted with indicated antibodies. (B) HeLa cells were treated with 50 μM of etoposide or vehicle (dimethyl sulfoxide, DMSO) for 6 hours and immunostained with the indicated antibodies. (C) HeLa cells were transfected with pSUPER-siHP1γ and immunostained with the indicated antibodies. (D) Nonproportional Venn diagram showing subsets of HP1γ-interacting proteins functionally related to DNA damage response pathways. The numbers in five subsets represent proteins that are interacting with HP1γ and functions in DNA damage response. IB, immunoblot; IP, immunoprecipitate.
We hypothesized that HP1γ translocates to DNA damage sites, following DNA damage with γH2AX. The HeLa cells were treated with etoposide and stained for HP1γ and γH2AX (damaged DNA sites or repair foci marker). HP1γ co-localized with γH2AX 6 hours after the treatment with etoposide (Fig. 3B), and this enabled us to determine whether HP1γ was a positive or negative regulator in the DNA damage response pathway. Notably, we discovered the accumulation of γH2AX at DNA damage sites in HP1γ-depleted cells (Fig. 3C), suggesting that HP1γ is required for the maintenance of genome integrity, as the deficiency of HP1γ may induce DNA damage. These data indicated that HP1γ functions as a positive regulator in the DNA damage response pathway.
Protein-protein interaction and post-translational modification play key roles in various biological processes. The co-immunoprecipitation affinity purification technique is widely used to investigate protein functions in these processes. In this study, we constructed an interactome network for HP1γ in order to investigate the protein’s novel functions. We identified several proteins as novel binding partners of HP1γ using an affinity purification system. Our results suggest that HP1γ functions in various biological processes (as listed in “Establishment of the interactome of HP1γ” section). BLM (No. 3 in Supplementary Table 1) and XRCC5 (No. 126 in Supplementary Table 1), essential proteins in the DNA damage response pathway, were identified in our purification analysis as proteins that interact with HP1γ (Table 1, Fig. 3D); this implies that HP1γ played a role in regulating the DNA damage response pathway. In the presence of a DNA damage signal, HP1γ can be translocated to DNA damage sites (Fig. 3A). Furthermore, phosphorylation of H2AX was induced in HP1γ-knockdown cells (Fig. 3B), suggesting that HP1γ is involved in the maintanence of genomic integrity by preventing DNA damage, and that HP1γ deficiency could lead to DNA damage. In summary, our interactome analysis demonstrated that HP1γ potentially plays a role in DNA damage response among various biological processes (Table 1, Fig. 3D).
Discussion
In this study, we suggested that HP1γ functionally links to the DNA damage response via its protein interactome. We found that HP1γ interacting proteins have a wide range of functions including DNA replication, nucleotide excision repair, mismatch repair, and homologous recombination. Consistent with this finding, HP1γ translocated to DNA damage sites following DNA damage and HP1γ was colocalized with γH2AX. Moreover, the γH2AX repair foci were formed by HP1γ-knockdown even in the absence of DNA damaging agents, suggesting that depletion of HP1γ causes genotoxic stress and HP1γ is required for the conservation of genome integrity in unstressed condition. Collectively, these findings suggest that HP1γ may have a role in DNA damage response pathway involving its interaction of repair proteins and other DNA damage response proteins.
Intriguingly, recent studies have shown that HP1 reinforces BRCA1 functions in the homologous recombination repair and the cell cycle checkpoint at the G2/M [18], and transcription [10] in response to genotoxic stress. The functional interplay between HP1 and BRCA1 in DNA damage response pathway is required for ensuring genome integrity. Notably, mutations of HP1γ that disrupt its binding to BRCA1 also cause defects in the BRCA1-mediated DNA damage response functions [10]. Therefore, these findings together with our results raise the possibility that HP1 facilitates DNA damage responses, which may involve its interactions. What the functions of HP1γ interactions are and how the interactions promote DNA damage responses in stressed and unstressed conditions are not elucidated. Future studies are needed to address the functions of the HP1γ interactions and the mechanisms in maintaining genome integrity and in response to DNA damage.
Conclusion
HP1γ binds to many proteins that have diverse cellular and biological functions. The proteins that interact with HP1γ are present in all major cellular compartments, including the nucleus, cytoplasm, and plasma membrane. Intriguingly, 14 HP1γ-interacting proteins identified in this study seem to be functionally connected to DNA response pathways (Table 1, Fig. 3D). Taken together, our results showed that HP1γ is likely to function in several biological processes, and potentially plays an important role in preserving genome integrity.
Acknowledgments
We thank all members of the Lee and Kang lab for helpful advice. This work was supported by the National Research Foundation of Korea (2011-0030043, 2014R1A2A2A01004444, and 2010-0018546).
Footnotes
Conflict of interest relevant to this article was not reported.
Electronic Supplementary Material
Supplementary materials are available at Cancer Research and Treatment website (http://www.e-crt.org).
References
- 1.Maison C, Almouzni G. HP1 and the dynamics of heterochromatin maintenance. Nat Rev Mol Cell Biol. 2004;5:296–304. doi: 10.1038/nrm1355. [DOI] [PubMed] [Google Scholar]
- 2.Jacobs SA, Khorasanizadeh S. Struc ture of H P1 chr omodomain bound to a lysine 9-methylated histone H3 tail. Science. 2002;295:2080–3. doi: 10.1126/science.1069473. [DOI] [PubMed] [Google Scholar]
- 3.Eissenberg JC, Elgin SC. The HP1 protein family: getting a grip on chromatin. Curr Opin Genet Dev. 2000;10:204–10. doi: 10.1016/s0959-437x(00)00058-7. [DOI] [PubMed] [Google Scholar]
- 4.Minc E, Courvalin JC, Buendia B. HP1gamma associates with euchromatin and heterochromatin in mammalian nuclei and chromosomes. Cytogenet Cell Genet. 2000;90:279–84. doi: 10.1159/000056789. [DOI] [PubMed] [Google Scholar]
- 5.Vakoc CR, Mandat SA, Olenchock BA, Blobel GA. Histone H3 lysine 9 methylation and HP1gamma are associated with transcription elongation through mammalian chromatin. Mol Cell. 2005;19:381–91. doi: 10.1016/j.molcel.2005.06.011. [DOI] [PubMed] [Google Scholar]
- 6.du Chene I, Basyuk E, Lin YL, Triboulet R, Knezevich A, Chable-Bessia C, et al. Suv39H1 and HP1gamma are responsible for chromatin-mediated HIV-1 transcriptional silencing and post-integration latency. EMBO J. 2007;26:424–35. doi: 10.1038/sj.emboj.7601517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.El Gazzar M, Yoza BK, Chen X, Hu J, Hawkins GA, McCall CE. G9a and HP1 couple histone and DNA methylation to TNFalpha transcription silencing during endotoxin tolerance. J Biol Chem. 2008;283:32198–208. doi: 10.1074/jbc.M803446200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Abe K, Naruse C, Kato T, Nishiuchi T, Saitou M, Asano M. Loss of heterochromatin protein 1 gamma reduces the number of primordial germ cells via impaired cell cycle progression in mice. Biol Reprod. 2011;85:1013–24. doi: 10.1095/biolreprod.111.091512. [DOI] [PubMed] [Google Scholar]
- 9.Liu X, Song Z, Huo Y, Zhang J, Zhu T, Wang J, et al. Chromatin protein HP1 interacts with the mitotic regulator borealin protein and specifies the centromere localization of the chromosomal passenger complex. J Biol Chem. 2014;289:20638–49. doi: 10.1074/jbc.M114.572842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Choi JD, Park MA, Lee JS. Suppression and recovery of BRCA1-mediated transcription by HP1gamma via modulation of promoter occupancy. Nucleic Acids Res. 2012;40:11321–38. doi: 10.1093/nar/gks947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cho HJ, Oh YJ, Han SH, Chung HJ, Kim CH, Lee NS, et al. Cdk1 protein-mediated phosphorylation of receptor-associated protein 80 (RAP80) serine 677 modulates DNA damage-induced G2/M checkpoint and cell survival. J Biol Chem. 2013;288:3768–76. doi: 10.1074/jbc.M112.401299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cho HJ, Lee EH, Han SH, Chung HJ, Jeong JH, Kwon J, et al. Degradation of human RAP80 is cell cycle regulated by Cdc20 and Cdh1 ubiquitin ligases. Mol Cancer Res. 2012;10:615–25. doi: 10.1158/1541-7786.MCR-11-0481. [DOI] [PubMed] [Google Scholar]
- 13.Li L, Monckton EA, Godbout R. A role for DEAD box 1 at DNA double-strand breaks. Mol Cell Biol. 2008;28:6413–25. doi: 10.1128/MCB.01053-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nozawa RS, Nagao K, Masuda HT, Iwasaki O, Hirota T, Nozaki N, et al. Human POGZ modulates dissociation of HP1alpha from mitotic chromosome arms through Aurora B activation. Nat Cell Biol. 2010;12:719–27. doi: 10.1038/ncb2075. [DOI] [PubMed] [Google Scholar]
- 15.Alcalay M, Meani N, Gelmetti V, Fantozzi A, Fagioli M, Orleth A, et al. Acute myeloid leukemia fusion proteins deregulate genes involved in stem cell maintenance and DNA repair. J Clin Invest. 2003;112:1751–61. doi: 10.1172/JCI17595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wang WY, Pan L, Su SC, Quinn EJ, Sasaki M, Jimenez JC, et al. Interaction of FUS and HDAC1 regulates DNA damage response and repair in neurons. Nat Neurosci. 2013;16:1383–91. doi: 10.1038/nn.3514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Paronetto MP. Ewing sarcoma protein: a key player in human cancer. Int J Cell Biol. 2013;2013:642853. doi: 10.1155/2013/642853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lee YH, Kuo CY, Stark JM, Shih HM, Ann DK. HP1 promotes tumor suppressor BRCA1 functions during the DNA damage response. Nucleic Acids Res. 2013;41:5784–98. doi: 10.1093/nar/gkt231. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.